氮磷添加对黄河三角洲盐地碱蓬光合特性和抗氧化系统的影响*
2023-08-03马金昭许文静邵鹏帅赵盈涵董凯凯张冬杰杨红军孙景宽
马金昭,许文静,2,邵鹏帅,赵盈涵,董凯凯,张冬杰,杨红军,孙景宽
(1.滨州学院,山东省黄河三角洲生态环境重点实验室,山东 滨州 256603;2.山东农业大学 林学院,泰山森林生态系统国家定位观测研究站,山东 泰安 271018)
黄河三角洲是由黄河携带的大量泥沙在入海口处不断堆积形成的缓冲带,是中国暖温带最完整的湿地生态系统,兼具河、海、陆等生态系统特性[1]。然而在其长期的成陆过程中,因复杂的人为和自然因素影响导致土壤盐渍化加重,严重抑制了植物根系对水分和养分的吸收[2],改变植物细胞膜功能,影响细胞中多种酶的活性和蛋白质构型,造成离子毒害,最终造成滨海湿地植被退化,生态系统遭到破坏[3-4]。近年来,由于化肥的大量施用、生活和生产污水排放以及水土流失等人类活动的影响,黄河口的营养盐入海通量始终保持在较高水平且有波动增加的趋势,这不仅影响到河口近岸水体的营养水平,还将对潮滩土壤的营养状况产生重要影响[5-6]。
氮和磷是植物生长发育和生理代谢的主要营养元素,影响着生态系统的结构与功能[7-8]。氮作为植物生命活动必需的大量营养元素,是蛋白质、核酸、叶绿素和次生代谢产物的重要组分,是植物光合作用的关键性限制因子[9-10]。磷是参与植物生长、繁殖及细胞间能量传递的重要元素,影响着核酸代谢以及叶绿素等光合蛋白的合成[11-13]。盐胁迫下,氮素可有效减少植物对Na+和Cl-的吸收与积累,提高地上部分K+含量[14]。研究发现:单独添加氮和磷时,达乌里胡枝子叶片的净光合速率、水分利用效率和气孔限制值均显著增加[9];同时添加氮磷可有效增加车桑子叶片的叶绿素含量,进而增加其净光合速率和PSⅡ活性[12];盐胁迫下,适量氮素可优化芥菜中乙烯的产生,从而调节脯氨酸含量,促进其光合作用和生长发育[15]。氮和磷是河口湿地的重要限制性养分元素,其在生态系统中的养分供应状况直接影响着植物的形态特征与生理性状[16-17],且氮磷之间存在着复杂的耦合关系[18],这对于缓解植物在生存环境中的养分限制与盐分胁迫尤为关键[7,19]。
黄河三角洲的天然植被组成简单,多以耐盐植物为主,其中盐地碱蓬(Suaeda salsa)具有极强的耐盐性,作为高盐土壤生长的先锋植物对生态系统的正常演替和生物多样性具有重要作用[20-21]。目前关于盐地碱蓬的研究多集中于其在盐碱胁迫下的种子萌发[22]、生态化学计量[23]以及生理生态与基因表达[24-26]等方面,关于其在氮磷添加下的光合特性和抗氧化系统研究较少,氮磷营养对盐地碱蓬生长发育的影响机制尚不清晰。因此,本研究以盐地碱蓬为对象,通过分析其在不同氮磷添加量下的光合生理参数与抗氧化酶活性变化,明确盐地碱蓬光合生理和抗逆系统对氮磷添加的响应特征,以期为黄河三角洲的植被恢复和生态改良提供理论依据。
1 材料与方法
1.1 研究区概况
试验地位于山东省东营市河口区仙河镇黄河三角洲区域(N38°1′18″,E118°44′2″),该区域属于暖温带大陆性季风气候,年均温度12.5 ℃,年均蒸发量1 900 mm,年均降水量660 mm,降水多集中于6—8 月。土壤类型为滨海盐土,其基本理化性状为:pH 7.83,电导率5.38 ms/cm,有机质含量16.2 g/kg,全氮含量0.32 g/kg,全磷含量0.64 g/kg,全钾含量11.2 g/kg,全盐含量7.82 g/kg。主要植被为芦苇和盐地碱蓬等。
1.2 试验设计
试验于2021 年5—10 月进行。选取地势相对一致、地上植被分布均匀的盐地碱蓬群落作为样地,以分析纯尿素(含N 46%)为氮源、磷酸二氢钾(含P2O552%)为磷源[13],采用裂区设计,主区为4 个氮处理(N0、N5、N15和N45分别为0、5、15 和45 g/m2),副区为2 个磷处理(P0和P1分别为0 和1 g/m2),共设置8 个处理:N0P0、N5P0、N15P0、N45P0、N0P1、N5P1、N15P1和N45P1。每个处理3 个重复,共计24 个小区,每小区样地面积为2 m × 2 m,采用随机区组设计。为避免交叉影响,小区间设置宽2 m 的缓冲带,并采用围栏隔断防止被破坏。于2021 年5—8 月中上旬,按照各处理小区所需氮磷量分别将氮磷源溶于4 L 去离子水后均匀喷施于样地表面,对照(N0P0)处理喷洒等量的去离子水,每次喷施间隔约1 个月,共喷施4 次。各小区的常规管理措施保持一致。
1.3 样品采集与测定
于2021 年8 月中下旬盐地碱蓬的生长旺盛期对每个样方进行样品采集与数据测定。在每个样方内随机选取5 株盐地碱蓬,采用钢卷尺测定株高;采用游标卡尺在植株根茎处测定茎粗;采用便携式叶面积仪(YMJ-B,杭州硕联)测定叶面积;将植株完整取样,去除泥土后测定其鲜质量;带回实验室放入烘箱105 ℃杀青30 min,以60 ℃烘干至恒质量,测定其干质量。
盐地碱蓬中上部健康、成熟功能叶片的光合参数(净光合速率Pn、气孔导度Gs、胞间CO2浓度Ci、蒸腾速率Tr和实际大气CO2浓度比值Ca)采用便携式光合仪Li-6400XT (LI-Cor,Lincoil,NE,USA)在设定光照强度1 000 μmol/(m2·s)、叶室内温度25 ℃下测定,并计算气孔限制值(Ls)和水分利用效率(WUE),计算公式为:Ls=1-Ci/Ca;WUE=Pn/Tr[8,11]。
采集各小区盐地碱蓬相同部位的健康功能叶片带回实验室进行鲜样测定。采用95%乙醇和80%丙酮的混合液(体积比为1∶1)浸提24 h,运用分光光度法分别在663、646 和470 nm 处测定叶片叶绿素含量[27];超氧化物歧化酶(SOD)活性采用氮蓝四唑NBT 法测定;过氧化物酶(POD)活性采用愈创木酚法测定;过氧化氢酶(CAT)活性采用过氧化氢法测定;丙二醛(MDA)含量采用硫代巴比妥酸法测定;可溶性蛋白含量采用蒽酮比色法测定[27]。
1.4 数据统计与分析
试验数据采用Excel 2010 处理;采用SAS 8.2 软件进行双因素差异显著性分析和ANOVA方差分析;采用Pearson 法进行相关性分析;采用R 语言4.1.0 软件进行主成分分析;采用Sigmaplot 12.0 制图。
2 结果与分析
2.1 不同处理对盐地碱蓬植株生长特征的影响
由图1 可知:不同氮磷处理对盐地碱蓬单株鲜质量和干质量的影响显著,且氮磷添加具有显著的交互作用。N45P1处理下盐地碱蓬的单株鲜质量和干质量均最高,较其他处理分别显著增加47.2%~645.6%和97.9%~543.9%;当氮添加量为0~15 g/m2时,是否添加磷对盐地碱蓬的单株鲜质量和干质量均无显著影响。此外,N45P0处理的鲜质量较N15P0、N5P0和N0P0处理分别显著增加62.0%、251.1%和406.6%。

图1 不同处理下盐地碱蓬的单株鲜质量与干质量Fig.1 Fresh and dry weight of per plant of Suaeda salsa under different treatments
由图2 可知:不同氮磷处理对盐地碱蓬的株高、茎粗和叶面积有显著影响,且氮磷交互作用对株高有显著影响。N45P1处理的株高分别较N45P0、N0P1、N5P1和 N15P1处理显著增加30.3%、277.2%、77.5%和57.5%;当氮添加量为15~45 g/m2时,与P0处理相比,P1处理可显著增加盐地碱蓬的株高。当添加磷时,N15和N45处理的茎粗较N0和N5处理显著增加,N45处理的叶面积较N0、N5和N15显著增加25.5%~116.1%。在相同氮添加量下,是否添加磷对盐地碱蓬的茎粗和叶面积均无显著影响。

图2 不同处理下盐地碱蓬的株高、茎粗和叶面积Fig.2 Plant height,stem diameter,and leaf area of S.salsa under different treatments
2.2 不同处理对盐地碱蓬叶片光合色素含量的影响
由表1 可知:不同氮磷处理对叶绿素a、叶绿素b 和类胡萝卜素含量有极显著影响,且对叶绿素a 和叶绿素b 含量存在显著的交互作用。与N15、N5和N0处理相比,未添加磷时,N45处理的叶绿素a、叶绿素b 和类胡萝卜素含量分别显著增加103.7%~233.7%、130.6%~274.1%和86.6%~204.6%;磷添加量为1 g/m2时,N45处理的叶绿素a、叶绿素b 和类胡萝卜素含量分别显著增加84.0%~408.4%、99.0%~499.6%和71.9%~269.9%。氮添加量为0~15 g/m2时,是否添加磷对叶绿素a、叶绿素b 和类胡萝卜素含量均无显著影响;氮添加量为45 g/m2时,添加磷可显著增加叶片的叶绿素a 和类胡萝卜素含量。
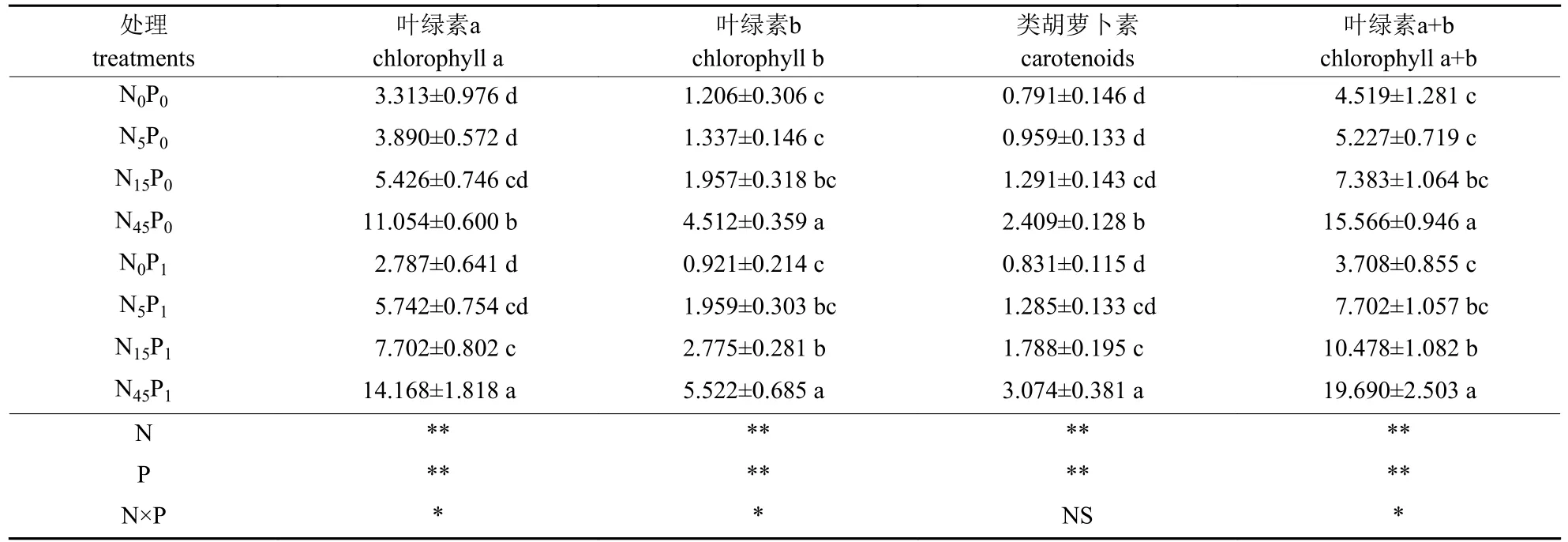
表1 不同处理下盐地碱蓬叶片的光合色素含量Tab.1 Photosynthetic pigment content of Suaeda salsa leaves under different treatments mg/g
2.3 不同处理对盐地碱蓬叶片光合特性的影响
由图3 可知:不同氮处理对盐地碱蓬叶片的净光合速率(Pn)有极显著影响,且未与磷处理有显著的交互作用。N45P0处理的Pn较N0P0、N5P0和N15P0处理显著增加20.9%~168.4%;N45P1处理的Pn较 N0P1和 N5P1处理显著增加58.2%~168.4%;当氮添加量为0~45 g/m2时,是否添加磷对Pn无显著影响。

图3 不同处理下盐地碱蓬叶片的光合特性Fig.3 Photosynthetic characteristics of S.salsa leaves under different treatments
由图3 还可知:不同氮处理对盐地碱蓬叶片的气孔导度(Gs)、胞间CO2浓度(Ci)和气孔限制值(Ls)有极显著影响,不同磷处理对Ci和Ls的影响极显著,且氮磷添加对Gs、Ci和Ls存在显著的交互作用。未添加磷时,N15处理的Gs较其他氮处理显著增加16.5%~81.3%;添加磷时,N5处理的Gs较其他氮处理显著增加38.8%~46.8%。N45P0处理的Ci较N0P0、N5P0和N15P0处理分别显著降低29.2%、30.1% 和 22.5%,N15P1和N45P1处理的Ci也显著低于N0P1和N5P1处理。未添加磷时,N45处理的Ls较其他氮处理显著增加37.5%~66.3%;添加磷时,N15和N45处理的Ls无显著差异。
图3 显示:不同磷处理对叶片水分利用效率(WUE)的影响显著,不同氮处理及与磷的交互作用对盐地碱蓬叶片的蒸腾速率(Tr)和WUE 的影响显著。当氮添加量为0~5 g/m2时,磷添加较未添加处理的Tr显著增加20.9%~44.3%;当氮添加量为45 g/m2时,是否添加磷对Tr无显著影响。当未添加磷时,N45处理的WUE 较其他氮处理显著增加37.5%~66.3%;当添加磷时,N15与N45处理的WUE 无显著差异;除氮添加量为15 g/m2外,其他氮添加量下P0与P1处理的WUE 均无显著差异。
2.4 不同处理对盐地碱蓬叶片抗氧化酶活性和脂质过氧化的影响
由表2 可知:不同氮处理对盐地碱蓬叶片的SOD、POD 和CAT 活性有极显著影响,且氮磷交互作用对叶片SOD 和POD 活性有显著影响。当氮添加量为0 和15 g/m2时,磷添加处理的叶片SOD 活性较未添加磷分别显著增加59.0%和20.6%;当氮添加量为0、5 和45 g/m2时,磷添加处理的POD 活性较未添加磷分别显著增加117.6%、67.3%和20.6%;当氮添加量为0~45 g/m2时,是否添加磷对CAT 活性无显著影响。与N0P1、N5P1和N15P1处理相比,N45P1处理的 SOD、POD 和CAT 活性分别显著增加15.7~149.6%、98.5%~260.2%和96.4%~294.4%。

表2 不同处理下叶片抗氧化酶活性以及丙二醛和可溶性蛋白含量Tab.2 Antioxidant enzyme activity,MDA,and soluble protein content of leaves under different treatments
由表2 还可知:不同氮处理对MDA 和可溶性蛋白含量有极显著影响。N45P1处理的MDA含量分别较 N0P1和 N5P1显著降低 56.2% 和 32.2%,但与N15P1和N45P0处理无显著差异;当氮添加量为0~45 g/m2时,是否添加磷对MDA 含量无显著影响。N45P1处理的可溶性蛋白含量最高,较N0P1、N5P1和N15P1分别显著增加111.0%、21.6%和41.4%,较N45P0处理显著增加43.1%;当氮添加量为0~15 g/m2,是否添加磷对可溶性蛋白含量无显著影响。
2.5 盐地碱蓬光合特性与抗氧化系统的关系
由表3 可知:盐地碱蓬的鲜质量和株高以及叶片的气孔限制值和水分利用效率与抗氧化系统呈极显著的相关关系(P<0.01)。盐地碱蓬叶片的叶绿素含量和净光合速率与抗氧化酶(SOD、POD、CAT)活性和可溶性蛋白含量呈极显著正相关、与MDA 含量呈极显著负相关(P<0.01);气孔导度和蒸腾速率仅与MDA 含量呈显著或极显著负相关(P<0.05 或P<0.01);胞间CO2浓度与抗氧化酶活性呈极显著负相关(P<0.01)、与MDA 含量呈极显著正相关(P<0.01)、与可溶性蛋白含量显著负相关(P<0.05)。

表3 盐地碱蓬生长特性、光合特性与抗氧化酶的相关性分析Tab.3 Correlation analysis among growth,photosynthetic characteristics and antioxidant of S.salsa
主成分分析发现前两个主成分解释了数据变异性的85.33%,能够较好地反映这12 个指标的表达信息。从主因子载荷向量(图4)中发现:第1 主成分解释了64.12%,主要包括以叶绿素含量、净光合速率、气孔限制值以及SOD 和POD活性为代表的因子;第2 主成分解释了21.21%,其中起作用的正向负载因子主要为气孔导度和蒸腾速率。延第1 主成分方向,N45P0、N45P1、N15P0和N15P1处理的光合特性及抗氧化系统与N0P0、N0P1组有明显分离;延第2 主成分方向N5P0、N5P1和与N0P0、N0P1组有明显分离。

图4 盐地碱蓬叶片光合特性与抗氧化系统的主成分分析Fig.4 Principal component analysis of photosynthetic characteristics and antioxidant system of S.salsa leaves
3 讨论
植物的生长发育与光合作用密切相关,光合作用是植物获取光能、合成有机物质、提高养分含量和促进器官发育的关键生物过程[28-29]。叶片净光合速率作为反映植物光合作用的直接指标,在本研究中随着氮素添加量的增加而显著提高,这与王雪梅等[12]的研究相似;然而在本研究中,添加磷素并未显著影响净光合速率。植物叶片的光合色素含量也是植物光合作用的关键因素,密切反映了植物的光合能力[29-30]。在本研究中,氮添加显著提高了叶片的叶绿素a、叶绿素b 和类胡萝卜素含量,且随着氮素添加量的增加而提高,磷也可显著提高植物的光合色素含量,且与氮素的交互作用可显著影响叶绿素a 和叶绿素b 的含量。盐地碱蓬叶片光合作用显著提高的主要原因是氮素的添加提高了叶片Rubisco 酶及其他参与光合作用的酶活性,促进了叶绿素合成[12,31],并且与添加的磷素发生耦合作用,共同促进了盐地碱蓬的光合作用。
氮磷营养的合理添加在影响光合作用的同时还能调节植物叶片的蒸腾速率,进而影响水分利用效率[9]。生长在盐渍化区域的植物通常具有细胞小、细胞壁厚和气孔导度小等特点,以减少水分流失,增施氮肥可以有效增大细胞尺寸,使细胞壁变薄,进而提高气孔导度[32],通过增加细胞间隙的CO2供应来提高作物的光合能力。在本研究中,氮添加显著影响叶片的气孔导度与胞间CO2浓度,且与磷添加具有显著的交互作用。5 g/m2氮添加量处理的蒸腾速率显著高于其他氮处理,而磷添加对其有显著影响。45 g/m2氮添加量处理的水分利用效率显著高于0 和5 g/m2处理,并与磷添加产生了显著的交互作用,促进了水分利用效率的提高,表明氮磷添加可以有效提高植物的水分吸收与利用能力,有助于其更好地适应不利环境;相关性分析发现:盐地碱蓬的光合特性与抗氧化系统存在显著的相关关系,叶片光合速率和叶绿素含量的提高可有效增加抗氧化酶活性,降低MDA 含量。
植物受到盐分胁迫时,体内急剧增加的活性氧自由基容易对机体造成严重损害,而植物体内会通过增加SOD、POD 和CAT 等抗氧化酶的活性来减少这种损害,从而使活性氧的产生与清除处于平衡状态[24,33]。张雪等[34]研究发现:氮素缺乏会引起黄瓜幼苗叶片内过氧化氢和超氧阴离子积累,进而造成抗氧化酶活性显著增强。本研究发现:添加氮使得抗氧化酶(SOD、POD、CAT)活性显著提高,且SOD 与POD 的活性随着氮素水平的增加而显著增加;磷添可显著增强SOD和POD 活性,且与氮添加产生显著的交互作用,进而提高盐地碱蓬的抗逆性。MDA 作为细胞膜脂过氧化的产物,其含量是反映膜脂过氧化作用强弱和膜脂损伤程度的重要指标[33,35]。氮添加显著降低了盐地碱蓬叶片中的MDA 含量,然而15 和45 g/m2处理间的MDA 含量无显著差异,这可能与其生命活性所需的氮素达到饱和状态有关。在外源氮添加的情况下,盐地碱蓬的可溶性蛋白含量显著增加,并与磷添加有显著的交互作用。在植物的生长发育过程中,氮和磷均是参与蛋白质合成的主要元素,故氮磷添加能有效提高盐地碱蓬抗氧化酶的活性,增强其生存能力。
4 结论
氮添加能显著影响盐地碱蓬的形态特征,且与磷素在生物量与株高上存在显著的交互作用,其中氮添加量为45 g/m2且配施磷处理的生物量最高。光合色素含量随着氮素添加量的增加而增加,且与磷素在叶绿素a 和叶绿素b 含量上有显著的交互作用。氮添加可显著提高叶片的光合特性,且氮磷添加对叶片Gs、Tr和WUE 有显著的交互作用。氮处理能有效增加叶片抗氧化酶活性和可溶性蛋白含量,降低MDA 含量,且对于SOD 和POD 活性氮磷交互作用显著。可见,氮磷添加可有效增加黄河三角洲盐地碱蓬的光合特性与抗氧化酶活性,促进其生长发育。
