大百合花序分化及生理生化特征
2023-07-15董知洋李雯琪张启翔高亦珂
魏 钰 董知洋 张 蕾 李雯琪 张启翔 高亦珂*
(1.北京林业大学 园林学院/城乡生态环境北京实验室,北京 100083; 2.北京市植物园管理处/北京市花卉工程技术中心,北京 100093)
球根花卉是指具有由地下茎或根变态形成膨大部分的多年生草本花卉,其特点是种类丰富、花型独特以及养护简便,因而在园林园艺上得到广泛应用。大百合(Cardiocrinumgiganteum(Wall.) Makino)是百合科大百合属多年生球根花卉,其植株高大挺拔,花朵洁白雅致,具有极高的观赏价值,享有“百合王子”的美誉[1];其鳞茎可供食用,果实可入药,具有一定经济价值[2]。大百合属为东亚特有属,在百合科的亲缘关系方面具有重要的分类学地位,被认为是百合科最原始的类群[3],具有较高的科研价值。由于大百合分布在山地林下潮湿地带,长期处于野生状态,其价值未能得到足够的认知和重视。大百合在自然条件下从生长到开花发育周期长且花期较短,在园林应用中受到局限,因此需要对其成花机理和开花特性进行探究。进行大百合休眠解除与花序分化特征的研究,是开展大百合成花机制研究与花期调控的理论与实践基础。
花是被子植物特有的生殖器官,其形态和结构具有丰富的多样性[4]。植物花器官的发育一般包括3个过程:成花诱导、花原基形成以及花器官形成与发育[5]。成花诱导是指植物从营养生长向生殖生长转化的过程,由遗传因子与多种环境信号共同促使植物的茎端分生组织转化为花序分生组织;花原基形成阶段,植物生长素不断通过极性运输积累到花序分生组织区域,形成花分生组织和花原基;花器官形成与发育阶段,在成花特征基因的作用下,花原基发育成完整的花器官[6]。植物的成花机制是一个复杂的调控体系,除遗传因子外,还受多种环境信号和内源途径调控,包括光周期、温度、春化及赤霉素等[7-9]。光是调节植物开花的主要环境因子之一,特别是光周期变化能够诱导植物进入繁殖阶段,例如短日照植物菊花,在长日照条件下会抑制开花[10]。环境温度会直接影响开花,总体来说较高的环境温度会促进开花,低温则会延迟开花[11]。中国水仙在高温环境下促进开花[12],而麝香百合和郁金香的开花则分别受到高温和低温的诱导[13]。内源激素在花发育过程中也起着重要作用,其中赤霉素不仅能促进植物器官生长,而且能诱导发育阶段的转变[14],这点在拟南芥中尤为明显[15],但是赤霉素的作用在不同植物中具有一定差异性[16]。球根花卉由于特殊的形态结构与生活习性,相比其它植物,其在花序(芽)分化过程中的生理特性和养分运输等方面有一定的特异性,主要特点是花序(芽)分化大都在地上部分休眠期完成,待外部环境合适后便可生长开花,但是不同的球根花卉花芽分化的特征差异较大。如郁金香花芽分化在夏季休眠期就已经完成[17];百合由于品种众多,花芽分化进程从9月到次年5月不等,且对低温需求的时间也不相同[18-20]。
大百合是一种极具开发潜力的球根花卉,其花序被认为是从有限花序到无限花序的过渡类型。目前,对大百合的研究主要集中在种质资源、亲缘关系、引种栽培以及繁殖技术等方面[21-25],尚未见到关于大百合花序分化过程的形态学及生理变化方面的报道。基于前人对其它植物花序分化的研究,本研究通过观察大百合鳞茎在北京地区自然越冬状态下花序和花芽分化及发育过程的形态特征,同时分析鳞茎内几种主要内源激素、可溶性糖和可溶性蛋白含量的变化规律,旨在探明大百合花序发育过程中茎端分生组织结构变化,解析大百合花序发育过程中内源激素、可溶性糖和可溶性蛋白含量的变化规律,以期为大百合开花机制研究以及花期调控提供理论依据。
1 材料与方法
1.1 试材及取样
供试材料大百合(C.giganteum)采自于湖南省邵阳市东安县(26°26′ N,111°0′ E),选取正常、无病虫害且周径在24 cm以上的鳞茎为研究材料。2020年9月将鳞茎露地栽植于北京植物园(39°59′ N,116°12′ E),行间距80 cm×80 cm,鳞茎覆土2 cm,常规管理。12月上旬浇冻水后,地表覆3 cm厚松针保墒。9月上旬开始每隔2周(发育后期每隔1周)取样,每次取3个鳞茎,共取样22次合计66个鳞茎,用以观测大百合花序分化进程至花芽分化完成。
1.2 试验方法
1.2.1花序发育解剖观察
样品制备:剥去鳞茎外层的鳞片及小鳞叶,切取顶芽生长点4~5 mm后置于3D数码显微镜(Leica M205 C,德国)观察拍照;用FAA固定液固定后置于4 ℃冰箱保存。用扫描电镜(Hitachi S-4800FESEM,日本)观察大百合花序发育过程并拍照。
1.2.2内源激素及营养物质含量的测定
顶芽取样观察的同时取大百合中层鳞片(3~4层),将不同球茎的鳞片混合,分别秤取1.5 g样品用于内源激素含量测定,称取2.0 g样品用于可溶性蛋白含量测定,称取1.0 g样品用于可溶性糖含量测定。每组样品均设置3个重复,放入液氮速冻后置于-80 ℃冰箱保存。
本研究采用选择性反应/多反应监测技术(SRM/MRM)质谱分析法[26],对大百合鳞茎内源激素进行定量测定。取标准品,用同位素内标法建立标准曲线;样品液氮研磨后取100 mg,经乙腈溶液提取、过滤、洗脱和吹干,定容至200 μL。采用Waters I-Class LC超高效液相色谱系统进行分离,5500 QTRAP质谱仪(AB SCIEX,美国)在正/负离子模式下进行质谱分析。通过绘制标准曲线计算样品各内源激素含量。采用蒽酮比色法[27]测定可溶性糖含量,采用考马斯亮蓝法[28]测定可溶性蛋白含量。
1.2.3数据分析
采用Microsoft Excel 2010软件进行数据分析并制图,采用SPSS Statistics 22.0软件进行数据误差分析(P<0.01,P<0.05)及相关系数分析。
2 结果与分析
2.1 大百合花序分化进程及其形态特征
大百合花序发育时间较长,从11月上中旬开始分化,到翌年4月上旬花芽全部分化完成,前后历时近5个月。按花序及花芽发育过程的形态特征,大百合花序分化过程可分为以下6个阶段(图1):

(a)花序未分化期(Ⅰ期),×70;(b)花序原基分化初期(Ⅱ期),×70;(c)苞片原基分化期(Ⅲ期),×45;(d)小花原基分化期(Ⅳ期),×35;(e)花被片原基分化期(Ⅴ期),×30;(f)花芽分化完成期(Ⅵ期),×30。下同。GC:生长锥;LP:叶原基;BL:苞片原基;IP:花序原基; SFP:小花原基;PE:花瓣原基;ST:雄蕊原基;PI:雌蕊原基。 (a) Undifferentiation period (stage Ⅰ), ×70; (b) Start of inflorescence primordium differentiation period (stageⅡ), ×70; (c) Bract primordium differentiation period (stage Ⅲ), ×45; (d) Floret primordium differentiation period (stage Ⅳ), ×35; (e) Perianth differentiation period (stage Ⅴ), ×30; (f) Flower bud differentiation completed period (stage Ⅵ), ×30. The same below. GC: Growth cone; LP: Leaf primordium; BP: Bract primordium; IP: Inflorescence primordium; SFP: Small flower primordium; PE: Petal primordium; ST: Stamen primordium; PI: Pistil primordium.
Ⅰ期:花序未分化期(图1(a))。10月底之前,大百合处于花序未分化期,即营养生长期。此阶段大百合茎端呈扁圆型,茎端周围有2~3片叶原基。由于顶端细胞没有边缘细胞活跃,因此叶原基伸长向内弯曲,同时能够对顶端生长点起到保护作用。
Ⅱ期:花序原基分化初期(图1(b))。10月底至11月中旬,大百合茎尖分生组织形态发生了显著变化,生长点隆起,由未分化时的圆盘形变成半球形,且生长点周边分化出多个苞片原基,这些改变标志着大百合茎端分生组织由营养生长转为生殖生长。
Ⅲ期:苞片原基分化期(图1(c))。11月中旬至翌年1月下旬,大百合的半球形花序原基不断发育,叶状苞片原基数量不断增加直至分化完成,最初形成的苞片原基逐渐远离花序轴,发育成为总状花序下部的苞片;近轴端逐渐被后出现的苞片原基所占据,数量为10~20个不等;此时可见小花原基的雏形。从花序原基分化初期到此阶段历经时间逾60 d,此时为北京地区最冷的时期,由于土壤结冰造成低温胁迫,植株的生理活动降至最低,花序发育缓慢。
Ⅳ期:小花原基分化期(图1(d))。1月下旬至2月下旬,大百合花序原基分化基本完成,每个苞片原基的内腋处明显可见半球形的小花原基。先分化出来的小花原基位于远轴端,进一步发育为花序下部的小花;近轴端小花原基出现晚,发育为花序上部的小花,完成总状花序发育。大百合小花原基分化期近30 d,此时随地表气温回升,植株内部形态变化明显较前期活跃,但是未观察到鳞茎外部有形态变化。
Ⅴ期:花被片原基分化期(图1(e))。2月下旬至3月中旬,小花原基开始进行花器官外层-花被片的分化。位于花序中部的小花原基花被片分化略早于顶端和外侧的小花原基,说明其不同部位花被片的分化顺序存在差异。此阶段花器官发育速度较前几个阶段快,仅20 d左右。
Ⅵ期:花芽分化完成期(图1(f))。3月中旬至4月上旬,每朵小花的雄蕊原基和雌蕊原基先后分化完成,标志着大百合整个花序及花芽分化完成。此时可以观察到鳞茎已发芽并伸出地面约2 cm。
解剖观察结果显示大百合花序分化规律为:1)先进行花序结构分化,然后是小花原基及花器官分化,前后共历时约150 d;2)大百合为有限总状花序,在分化Ⅳ期,可以看到花序顶端分生组织逐步缩小形成一个球形器官(不发育成小花原基),占据了花序顶端位置,起到“封顶”作用,从而导致花序发育的终止,转为小花原基分化阶段;3)小花原基的起始是由花序基部到顶部,从茎端分生组织外围到分生组织中央,具有时序性;花序基部的小花原基虽然先分化出来,但是到了分化Ⅴ期,可以明显看出花序中上部的小花早于基部小花进行花器官分化,而最后所有小花基本同步完成花器官的分化。大百合花序分化过程中不同位置小花的发育存在渐趋同步的特点。
2.2 大百合花序分化过程中内源激素的动态变化
2.2.1大百合花序分化过程中内源激素的含量变化
对大百合花序和花芽分化过程中6种内源激素含量变化进行分析,结果见图2。
吲哚-3-乙酸(IAA)是生长素类中最主要的一种植物激素,其主要作用是促进细胞分裂、伸长和分化,也影响营养器官和生殖器官的生长发育[29],IAA与花芽分化的关系一直存在争议。如图2(a)所示,大百合IAA含量在未分化阶段最低,在花序和花芽分化过程中含量明显增加,在原基分化初期和花芽分化完成期呈现双峰的特征,特别是在花芽分化完成期达到峰值;在分化过程中间阶段差异不显著。在大百合花序未分化时,植株处于休眠状态,因此IAA含量较低;花序分化开始后IAA含量显著升高,推测在花序形成及花器官分化阶段需要大量IAA参与形态建成。
赤霉素(GAs)是调节植物生长发育的重要激素,生理作用主要包括促进茎的伸长、花粉发育、侧枝生长、种子发芽及果实生长等[30],其在植物体内的运输没有极性,对不同植物的作用也具有一定的差异[13]。在众多已发现的赤霉素类激素中有生物活性的主要为GA1、GA3、GA4和GA7,作用各不相同。4类赤霉素在大百合花序分化过程中含量的变化总体趋势是一致的:均为先低后高,在花芽分化完成时均达到最高(图2(b))。含量上,GA3最高,GA7最低,说明GA3是起主要作用的赤霉素类激素。
玉米素核苷(ZR)是最活跃和普遍以天然形式存在的细胞分裂素,在结构上有顺式玉米素核苷(cZR)和反式玉米素核苷(tZR)2种异构体,其主要作用是促进细胞分裂和膨大等,保证植株生长发育[31]。在大百合花序分化各个阶段2种异构体的总体变化趋势一致,即在花序未分化时含量较低,随着花序分化进程含量逐渐增加,到小花原基分化期达到顶峰,之后又呈现下降趋势(图2(c))。
乙烯是气体激素,在合成部位起作用,不被运转;但是其前体1-氨基环丙烷羧酸(ACC)在植物体内能够被运输,因此有学者认为ACC才是激素[29]。在有氧条件下,ACC含量多少决定了生成乙烯的含量。大百合花序发育过程中ACC含量的变化如图2(d)所示,在花序分化初期,ACC含量最高且与其它阶段差异极显著;随着花序发育,ACC含量呈下降趋势,在雄蕊和雌蕊分化期含量最低,且与其它阶段差异极显著。
茉莉酸(JA)普遍存在于高等植物中,是一类在植物体内起全局调控作用的生长调节物质,通常分布于植物的幼嫩组织和发育的生殖器官中,通过信号传导途径主要调节植物生长发育进程以及植物防御系统[32]。大百合花序分化过程中可以看到JA含量呈明显下降趋势,在花序未分化时最高,与其它阶段具有极显著差异;从花序分化初期含量显著下降,分化过程中各阶段含量无显著差异(图2(e))。
水杨酸(SA)是植物体内广泛存在的一种天然酚类化合物,是重要的内源信号分子,其不仅能够调节植物的生长发育过程,还具有提高植物抗逆性的作用,诱导植物产生抗逆性以抵抗不良因素造成的伤害[33]。在大百合花序分化过程中SA变化如图2(f)所示:在苞片原基分化期,SA含量最高,与其它时期具有极显著差异;而其它几个分化时期之间则无显著差异。
2.2.2大百合花序分化过程中内源激素的平衡
在大百合花序分化过程中,3种内源激素比值变化详见图3。IAA/GA3与cZR/GA3的动态趋势基本一致,呈明显的单峰曲线变化趋势,均在花序原基分化初期达到一个峰值,而随后趋于稳定。表明高比值的IAA/GA3、cZR/GA3与大百合的花芽分化密切相关,且有利于大百合从营养生长向生殖生长转换。IAA/cZR在分化过程中出现了2个峰值,分别是花序原基分化初期和花芽分化完成阶段,推测IAA/cZR在花序分化的初期和花芽分化完成期都起到了重要调节作用。

图3 大百合花序分化过程中3种内源激素比值变化
2.3 大百合花序分化过程中营养物质的动态变化
2.3.1可溶性糖与可溶性蛋白含量变化
糖和蛋白质都是植物体内重要的有机物,是植株生长发育的物质基础。糖为植物提供能量来源,也是代谢中间体[34];蛋白质是构成细胞和生物体结构的物质,可溶性蛋白是重要的营养物质和渗透调节物质[35]。大百合花序分化过程中可溶性糖与可溶性蛋白含量变化如图4所示。可溶性糖含量总体呈上升趋势,仅在花被片分化期略有下降,推测此阶段为多个小花的花器官分化期,对可溶性糖的消耗较大;可溶性蛋白含量整体呈下降趋势,在未分化期最高,在分化过程中花被片分化期较高,推测在花序和花芽分化过程中需要大量的可溶性蛋白供给花器官的形态建成,在花芽分化完成后可溶性蛋白含量则显著下降。
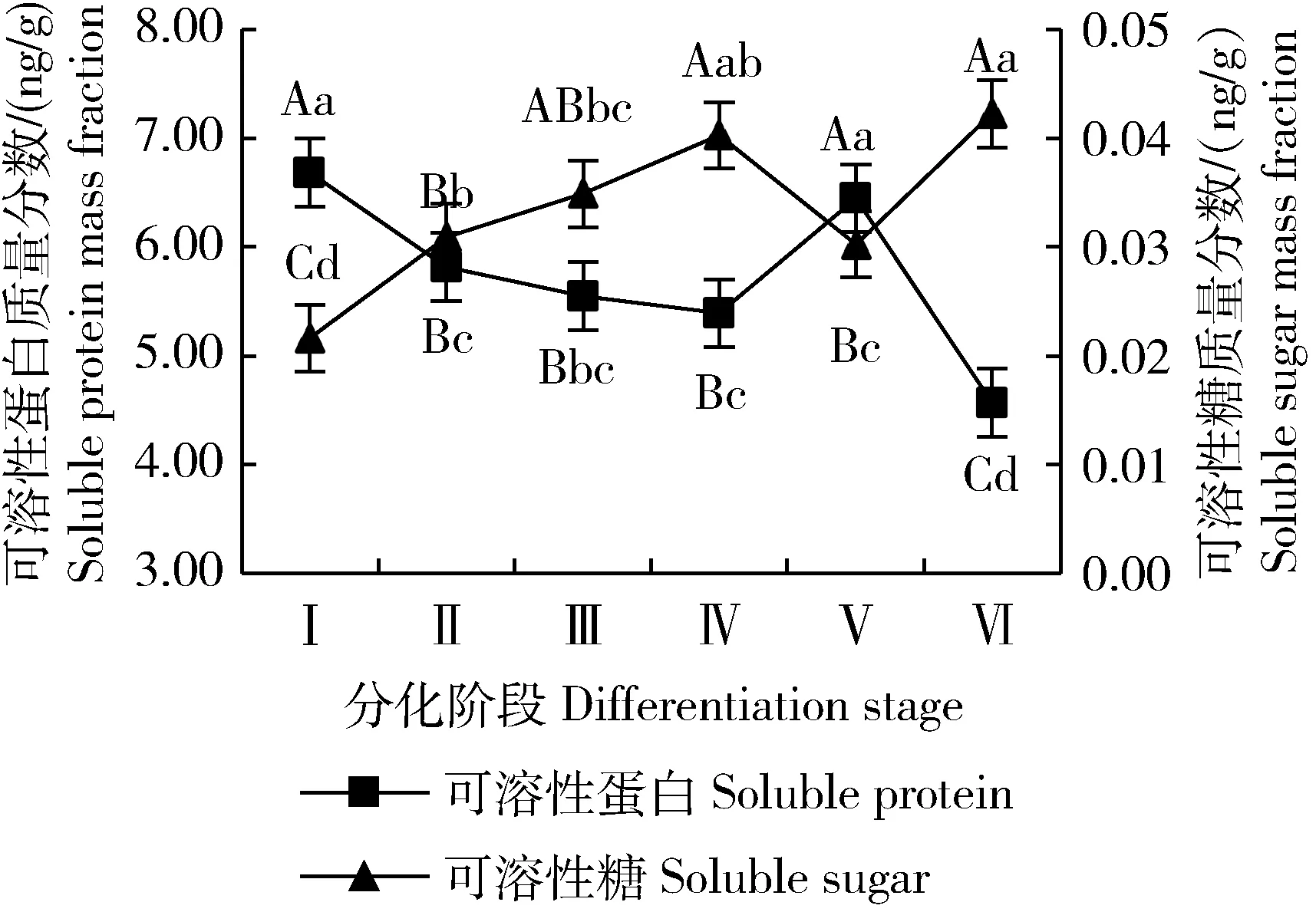
图4 大百合花序分化过程中可溶性糖 与可溶性蛋白质量分数变化
作为植物生长发育的能量来源及调控因子,可溶性糖与可溶性蛋白在大百合花序分化过程中含量的变化趋势是相反的。在未分化期,可溶性糖含量最低,而可溶性蛋白含量最高;在花芽分化完成期,可溶性糖含量达最高,可溶性蛋白含量则为最低,说明二者在大百合花序分化过程中存在一定平衡关系。
2.3.2内源激素与营养物质相关性分析
在大百合花序分化过程中,各内源激素与营养物质含量的相关系数如表1所示,IAA、GAs和ZR含量都与可溶性糖含量呈正相关,与可溶性蛋白含量呈负相关。其中GA3与可溶性糖和可溶性蛋白含量的相关系数均为最高,分析GA3在与营养物质协同作用促进花序分化过程中起着积极作用。ACC和JA则与之相反,二者与可溶性糖含量呈负相关,与可溶性蛋白含量呈正相关。SA含量与营养物质含量几乎不相关,推测可能是SA主要参与植株的抗逆性,特别是与低温胁迫导致的抗寒性有关,与花芽分化关系不大。
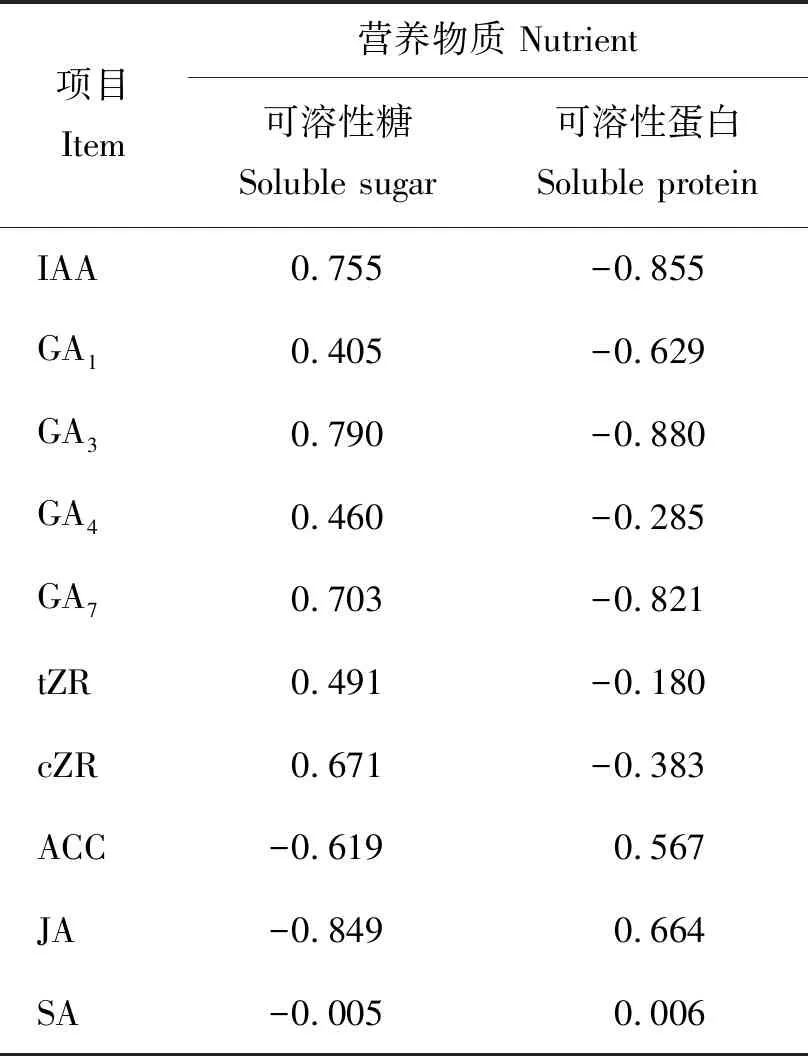
表1 大百合花序分化过程中内源激素含量与 营养物质含量的相关系数
3 讨论与结论
3.1 大百合花芽分化过程
从营养生长阶段到生殖生长阶段的转变,是植物发育变化的关键过程,有些植物则是营养生长和开花同时进行,大白合的花序分化发生在地上部分休眠期,与很多鳞茎植物如百合和番红花等相似[20,36]。本研究明确了大百合花序分化的进程,根据其形态特点划分为6个阶段:花序未分化期、花序原基分化初期、苞片原基分化期、小花原基分化期、花被片分化期和花芽分化完成期。研究发现大百合花序分化过程中不同位置小花的发育存在渐趋同步的特点,即小花发育早期由于发育时序不同,小花原基大小存在差异,但是到了花芽分化完成期小花的大小基本达到一致。这个现象在与其近源的百合、郁金香和豹子花等物种的研究中未见报道,相关分子机制有待进一步研究。
此外,后期物候观测显示,同一来源和批次的大百合种球,开花后每个花序的平均花朵数明显少于取样时观察到的小花原基数量,推测在后期生长发育过程中伴随着花序基部小花退化的情况存在,具体原因还需要持续观察与探索。
3.2 内源激素与大百合花芽分化的关系
内源激素在植物成花与花形态构建过程中起着重要作用,是植物花序(芽)分化的关键诱导因子之一,对茎端分生组织由营养生长向生殖生长转变具有重要的调控作用。内源激素的含量以及激素间平衡与成花过程密切相关[37]。大百合花序发育过程中,IAA和GAs总体呈上升趋势,表明高水平IAA和GAs有利于花序以及花器官的分化,这与对石蒜和百合等植物的研究结果一致[38-39]。有研究表明GAs几乎对每轮花器官发育都有一定的调控作用[30],特别是雄蕊的发育需要更高的GAs含量[40],本研究结果也验证了这一结论。ZR和SA为先上升后下降的单峰变化,分析二者在分化中期起到较为积极的调控作用;特别是ZR在小花原基分化期达到峰值,推测其对花器官细胞的分裂和增大起到了重要作用。ACC和JA总体呈下降趋势,在花序未分化期二者含量较高,可能与低温胁迫导致的抗寒性及休眠有关,花序分化后ACC与JA含量降低推测可能是解除了对花序(芽)发育的抑制作用。
花序(芽)分化过程中形态的变化不仅受到激素含量的影响,同时也与激素之间的平衡有重要关系[41]。大百合花序分化过程中IAA/GA3和cZR/GA3的变化趋势一致,均在花序分化初期达到峰值,分化过程比较稳定,表明高比值的IAA/GA3与cZR/GA3有利于促进大百合从营养生长向生殖生长转换;IAA/cZR在花芽分化完成期达到峰值,在花序原基分化初期为次峰值,推测其对植株发育阶段的转变起着重要调节作用。总体看来,细胞分裂素对苞片原基和小花原基等新器官的形成起重要作用,而生长素和赤霉素在花芽分化后期形态建成作用更显著。
3.3 营养物质与大百合花芽分化的关系
花序(芽)分化过程中需要消耗大量的营养物质。有研究显示,碳水化合物是完成花形态建成的重要物质基础[42-43]。可溶性糖是花芽分化过程中可以直接运输和利用的养分形式,是植物生长发育的能量基础,糖及其代谢物对植物的成花具有调节作用[34]。大百合花序分化过程中可溶性糖含量总体呈上升趋势,在小花原基分化期达到一个高峰,说明较高水平的可溶性糖含量可以促进大百合花序和花芽的分化与发育,这与西红花等球根花卉的花芽发育规律相似[44]。可溶性蛋白是花器官形态建成的重要物质基础,在花芽分化时需要较大量。大百合花序分化过程中可溶性蛋白始终保持较高的含量,说明其需要大量的蛋白质供给花器官的形态建成。一般情况下,组织细胞代谢活性旺盛则可溶性蛋白的含量会较高,本研究结果显示可溶性蛋白的含量整体呈下降趋势,尤其是花芽分化后期,推测可能此阶段细胞体积迅速增加的速率大于可溶性蛋白的增加速率而导致这一现象。可溶性糖与可溶性蛋白含量的变化趋势完全相反,推测在有限的营养物质中,二者通过平衡关系来调控成花的过程。
大百合是东亚特有植物,也是我国具有特色的野生球根花卉之一。本研究明确了大百合花序及花芽分化进程以及形态特征,了解了大百合花序分化过程中内源激素和营养物质的变化规律,为大百合生物学特性及成花机理研究提供了基础资料,同时可作为大百合花期调控的参考依据。大百合属于温度敏感型植物,其花序分化的具体时间及每个发育阶段持续的长度与温度有一定关系,相关规律还需进行深入探究。
感谢中国科学院植物研究所孔宏智研究员、姜闯道研究员和傅学浩博士在研究过程中给予的指导与帮助。
