谷田土壤中细菌群落对铬胁迫的响应特征
2023-07-13景秀清赵鹏宇白雪邓宁赵晓东吴丽华仪慧兰
景秀清,赵鹏宇,白雪,邓宁,赵晓东,吴丽华,仪慧兰
(1.太原师范学院生物科学与技术学院,山西 晋中 030619;2.山西大学生命科学学院,太原 030006)
随着采矿、化工、冶金、运输等工业活动的发展,以及各种农用化学品的使用,重金属不断被释放到环境中,造成农田土壤的大面积污染[1]。目前,我国每年因重金属污染而造成的粮食减产和损失高达1 200万t[2]。由于重金属在土壤中难降解、毒性强、迁移性强,其不仅会对土壤性质,尤其是土壤微生物群落产生不良影响,而且也会被植物吸收后通过食物链进入人体,从而危害人体健康[3]。
微生物作为土壤生态系统的重要组成部分,直接参与土壤养分的循环过程,当土壤环境发生改变时,微生物群落能快速地反映出环境质量的变化情况,是衡量土壤污染与恢复程度的重要指标[4-5]。根际土壤是微生物代谢活动的主要位点,也是重金属在植物体内迁移与积累的主要部位[6]。有研究发现,土壤中重金属积累水平超过临界值后,会使土壤中微生物多样性及微生物群落结构发生变化,从而显著影响土壤生态系统的结构与功能[6-7]。重金属铬(Cr)对土壤中微生物活性和土壤肥力的影响巨大,Cr 污染显著影响土壤理化性质以及微生物群落多样性,不同程度Cr污染的土壤样品中,细菌群落结构与组成差异显著[4,8-10]。同时土壤中的Cr 及其衍生物也会通过环境影响植物的正常生理活动、生长发育及生物量的累积,对植物造成巨大的伤害[11]。鉴于农田土壤污染现状的严峻性以及重金属对农田土壤的长期危害性,本研究将Cr 胁迫视为一种“生态扰动”,研究Cr 胁迫对农田土壤细菌群落组成、结构及构建机制的影响。
‘晋谷21 号’是山西省农业科学院经济作物研究所培育的优质高产谷子(Setaria italica)新品种,谷子幼苗长势较强、抗旱性强、结实性好、抗病、稳产且早熟[12]。本研究以种植‘晋谷21 号’谷子的土壤为研究对象,通过添加外源重金属Cr,探究谷田土壤中细菌群落对Cr 胁迫的响应特征,揭示谷子根际土壤中细菌群落在Cr 胁迫后不同时间节点的动态变化规律,为揭示重金属污染土壤中细菌群落的演变特征提供理论依据。
1 材料与方法
1.1 谷子的种植
实验用谷子为山西省农业科学院经济作物研究所选育的优质高产谷子品种“晋谷21 号”[13]。盆栽试验在实验室中进行。实验用土壤平均pH 7.65,总氮0.13 g∙kg-1,总磷1.32 g∙kg-1,不含重金属Cr,土壤有机质含量为32 g∙kg-1。选取直径15 cm、盆深20 cm的花盆,装500 g土壤,种植前用灭菌蒸馏水调整土壤湿度至田间持水量的60%左右;挑选籽粒饱满的‘晋谷21 号’种子,均匀点放于花盆中,随后将花盆用保鲜膜包裹后置于光照培养室(光照周期16 h/8 h,温度26 ℃)培养,直至谷子出苗方可揭开保鲜膜。继续培养两周左右,待谷子幼苗长势均一开始胁迫处理。
1.2 重金属胁迫处理
实验共分3 组:未处理组(胁迫前,CK)、Cr6+胁迫6 h 组(Cr_6h)和Cr6+胁迫6 d 组(Cr_6d)[10]。每个处理平行做6 个重复实验,共18 个样本。处理时,在对照组托盘中每盆施加300 mL 的H2O,在试验组托盘中每盆施加300 mL 的1 mmol∙L-1K2Cr2O7[2,14-15],分别于处理6 h 或6 d 后取样。取样时尽量保持根际土壤的完整性,避免破坏性取样。所取土样储存在-80 ℃冰箱,用于后续序列测定。
1.3 DNA的提取与高通量测序
1.3.1 DNA的提取
称取0.5 g 土壤,参照Power Soil 试剂盒说明书提取土样中的DNA。利用琼脂糖凝胶电泳检测所提取的DNA是否降解,使用酶标仪Infinite200 PRO测定提取物的纯度与浓度,A260/A280吸光度位于1.8~2.0之间,表示所提取的DNA纯度较高,可用于下一步分析。
1.3.2 PCR扩增与测序
将提取的基因组DNA 进行PCR 扩增。扩增体系如下:Taq DNA 聚合酶(5 U·µL-1)0.5 µL,10×PCR 缓冲液5.0µL,dNTPs 溶液4.0µL,模板DNA(20~30 ng·µL-1)1.0µL,正、反向引物(10µmol·L-1)各1.0µL,用无菌水补充至50µL。反应条件:98 ℃预变性1 min;98 ℃10 s,50 ℃30 s,72 ℃30 s,共30个循环;72 ℃终延伸5 min。混合均匀的DNA 样品送至上海美吉生物医药科技有限公司,通过Illumina Miseq 测序平台,对16S rDNA 高变区(V3~V4区)进行测序。待测序完成后进行数据的分析处理。
1.4 生物信息学与统计学分析
原始数据通过序列拼接、质量控制、去除嵌合体等得到最终的高质量序列。按照97%的序列相似性将序列聚类为操作分类单元(OTUs),对原始的OTUs进行重新抽平,最终使得每个样品的序列数目相同,以重新抽平的OTUs 丰度数据为基础进行物种注释分析[16],从多种分类水平统计土壤样本中细菌群落的组成状况。
利用单因素方差分析(ANOVA)评估不同处理土壤细菌群落的组成、α 多样性和群落构建过程[17-18]。使用Venn 图、优势物种相对丰度分析评估Cr 胁迫处理前后土壤细菌群落的组成结构[19]。采用Sobs指数、Shannon 指数、Simpson 指数、ACE 指数、CHAO 指数等研究细菌群落的α多样性[18,20]。利用非度量多维尺度分析(NMDS)[18,21-22]对Cr 胁迫后不同时间细菌群落的时空分布格局进行排序分析。通过PICRUSt2软件包将细菌测序结果与KEGG 数据库比对分析获得其功能注释[18,23]。
1.5 群落构建
种间网络分析的参数运算使用R语言中的igraph程序包[24],并在Gephi 软件中实现可视化。利用R 语言中的Hmisc 等程序包构建中性群落模型(NCM)[25],进一步判断土壤细菌群落的构建机制。参考Stegen等[26]提出的基于系统发育结构的分析方法,观察土壤中细菌群落构建过程中的驱动因素。当所有样品中|betaNTI |≤2,说明随机过程驱动微生物群落结构的变化,当 |betaNTI |>2,说明确定过程驱动微生物群落结构的变化[27-28]。
2 结果与分析
2.1 Cr胁迫对细菌群落结构和多样性的影响
2.1.1 对细菌群落结构的影响
通过对CK、Cr_6h 和Cr_6d 这3 组共18 个样品进行高通量测序,共获得791 427条有效序列,有效序列碱基数目共计329 176 199 bp,平均序列长度415 bp。对各组样品的OTUs 以及组间样品的重叠OTUs 进行Venn 图可视化分析(图1),发现这3 组样本的OTUs总数分别为3 133、2 971 个和3 140 个,特有的OTUs的数目分别是CK 组138 个(3.93%)、Cr_6h 组58 个(1.65%)和Cr_6d 组114 个(3.25%),3 组样本共有的OTUs的数目为2 536个(占所有OTUs的72.27%)。

图1 细菌群落OTUs数量Venn图Figure 1 OTUs Venn diagram of bacterial communities
各组样品中的细菌群落共包含38 个门、117 个纲、278 个目、468 个科、843 个属和1 487 个种。在门水平,Cr 胁迫前后土壤中的细菌群落组成基本相似,共比对出11 个相对丰度大于1%的细菌门(图2A)。其中放线菌门(Actinobacteriota)平均相对丰度最高,达到31.11%;其次是厚壁菌门(Firmicutes,19.00%)、变形菌门(Proteobacteria,18.82%)和绿弯菌门(Chlo-roflexi,11.87%),相对丰度介于10%~20%之间;拟杆菌门(Bacteroidota,4.57%)、芽单胞菌门(Gemmatimo-nadota,3.02%)、酸杆菌门(Acidobacteriota,2.44%)和异常球菌门(Deinococcota,2.22%)的相对丰度介于2%~10%之间。与CK 组相比,随着Cr 胁迫时间的延长,优势菌门中拟杆菌门相对丰度持续上升;放线菌门和绿弯菌门在Cr_6h 时的相对丰度有所上升,持续胁迫至Cr_6d,相对丰度未进一步发生显著变化;芽单胞菌门和酸杆菌门的丰度先下降后上升,厚壁菌门相对丰度则持续下降。

图2 细菌群落在门水平和属水平对Cr胁迫的响应Figure 2 Effects of Cr stress on the relative abundance of bacterial community structure at phylum and genus levels
在属水平,共比对出7 个相对丰度大于2%的细菌属(图2B)。其中直丝菌属(Planifilum)平均相对丰度最高,达到6.34%;长孢菌属(Longispora,4.59%)和norank_f_AKYG1722(4.16%)的平均相对丰度大于4%;马杜拉放线菌属(Actinomadura,2.66%)、嗜盐多孔菌属(Haloactinopolyspora,2.51%)、特吕珀菌属(Truepera,2.22%)和糖单胞菌属(Saccharomonospora,2.04%)的平均相对丰度介于2%~3%之间。直丝菌属的相对丰度随着Cr 胁迫时间的延长而下降;长孢菌属和norank_f_AKYG1722的相对丰度先剧烈上升,而后趋于稳定;马杜拉放线菌属和嗜盐多孔菌属的相对丰度先剧烈上升,而后略有下降。长孢菌属和norank_f_AKYG1722在Cr 胁迫前后的相对丰度发生明显变化。上述结果表明Cr 胁迫致使细菌群落组成变化显著。
2.1.2 对细菌群落α多样性的影响
通过单因素方差分析探究细菌群落α 多样性(Sobs 指数、Shannon 指数、Simpson 指数、ACE 指数和Chao指数)在Cr胁迫不同时间节点的变化(图3)。Cr胁迫后,细菌群落的Sobs 指数和ACE 指数无显著变化(P>0.05);Shannon 指数在Cr_6h 较CK 显著下降,在Cr_6d 又有所上升(CK 为6.07、Cr_6h 为5.92、Cr_6d为6.04);Simpson 指数在Cr_6h 上升,在Cr_6d 下降(CK 为0.006 8、Cr_6h 为0.007 8、Cr_6d 为0.006 8);而Chao 指数在Cr_6h 下降,在Cr_6d 回升至CK 水平(CK为2 666、Cr_6h为2 526、Cr_6d为2 634)。结果表明Cr胁迫对土壤细菌群落α多样性影响显著。

图3 Cr胁迫对细菌群落α多样性的影响Figure 3 Effects of Cr stress on α diversity of bacterial community
利用非度量多维尺度分析,基于Bray-Curtis距离分析Cr 胁迫对土壤中细菌群落时空分布格局的影响。图4 结果表明,处于同一时间节点的细菌群落结构在时空分布格局上高度相似,处于不同时间节点的细菌群落结构在时空分布格局上差异较大。组间差异显著性分析结果(ADONIS,P=0.001)表明Cr 胁迫对土壤细菌群落时空分布格局影响显著。

图4 Cr胁迫对细菌群落时空分布格局的影响Figure 4 Effects of Cr stress on spatial and temporal distribution pattern of bacterial community
2.2 Cr胁迫对细菌群落构建机制的影响
2.2.1 细菌群落构建过程
根据Cr胁迫不同时间段土壤细菌群落的betaNTI值大小(图5),结合Stegen等的群落构建的划分方法[26],推断Cr胁迫对细菌群落构建机制的影响。Cr胁迫使细菌群落的betaNTI值显著下降(即箱线图中的中位数),在CK、Cr_6h、Cr_6d阶段分别为-2.68、-2.11、-1.91,表明细菌群落构建过程在CK 和Cr_6h 阶段由确定过程驱动(|betaNTI |>2),而在Cr_6d阶段由随机过程驱动(|betaNTI |<2)。即随着Cr 胁迫时间的延长,细菌群落构建由确定过程驱动逐渐转变为由随机过程驱动,群落构建机制呈现阶段性变化特征。

图5 Cr胁迫对细菌群落构建机制的影响Figure 5 Effects of Cr stress on bacterial community assembly processes
基于OTU 矩阵绘制中性群落模型(图6),估算OTU 发生频率与其相对丰度变化间的关系。CK、Cr_6h 和Cr_6d 组的模型分别解释了群落方差的63.7%、62.3%和65.3%,总体解释度为76.9%。细菌群落Nm值[元群落规模(N)与迁移率(m)的乘积]在Cr_6d(Nm为33 365)阶段最小,表明细菌群落在该阶段受到的扩散限制作用较大,相对而言在CK(Nm为36 720)、Cr_6h(Nm为37 370)阶段受到的扩散限制作用较小,分布较广泛。
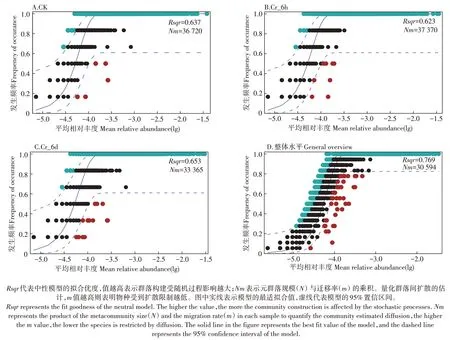
图6 中性细菌群落模型Figure 6 Neutral bacterial community model
2.2.2 细菌群落种间关系网络分析
通过细菌群落共发生网络分析,利用Spearsman相关性分析评估不同变量间的相关性,构建种间关系网络图,探究Cr 胁迫对土壤细菌群落种间关系的影响。表1显示,Cr胁迫后细菌群落共生网络的边数量和平均度持续下降,表明随着Cr 胁迫时间的延长,细菌的网络规模变小,种间相互作用关系趋于简单;与此同时,Cr 胁迫使细菌群落共生网络的平均路径长度上升,表明随着Cr 胁迫时间的延长,土壤细菌菌群间响应速度变慢,不易受外界环境的干扰。
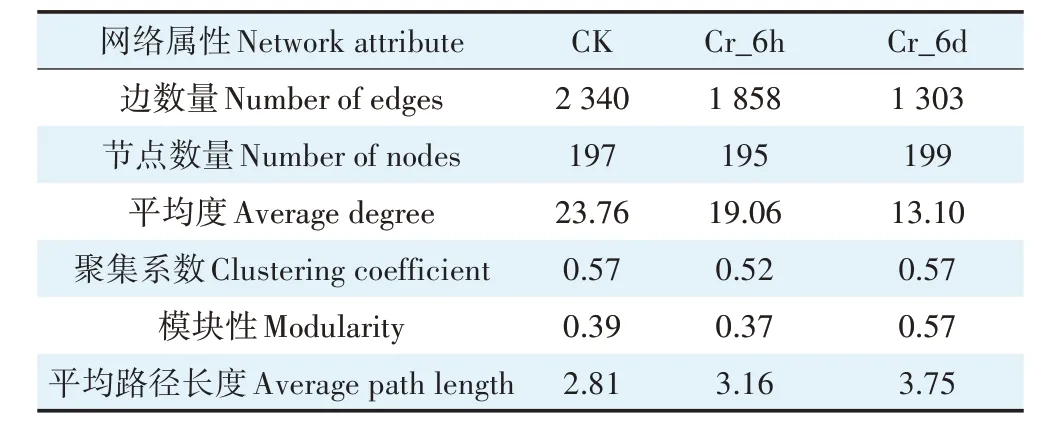
表1 土壤细菌群落网络属性Table 1 Network attributes of soil bacterial communities
根据种间关系网络图(图7)分析可知,Cr 胁迫不同时间节点的关键物种不同:CK 阶段的关键物种为直丝菌属、芽孢杆菌属(Bacillus)、链霉菌属(Strepto-myces)等,Cr_6h 阶段为特吕珀菌属、马杜拉放线菌属、节杆菌属(Arthrobacter)等,Cr_6d 阶段为火山岩海球菌(Marmoricola)、黄色类固醇杆菌(Steroidobacter)等,两者分别属于放线菌门与变形菌门。由此可见,不同阶段土壤细菌群落优势菌属多属于放线菌门与变形菌门,其对重金属具有较强耐性。且在Cr 胁迫不同时间节点上,细菌群落的模块变化趋势与表1 一致,同一模块中的细菌群落以正相关为主,表明Cr 胁迫导致细菌群落以共生关系为主导,竞争关系较弱,且群落种间关系呈阶段性变化特征。

图7 细菌种群间共生网络图Figure 7 Inter-population symbiosis network diagram of bacterial
2.3 Cr胁迫时间序列上细菌群落代谢活性分析
通过PICRUST2 对KEGG Pathway level 2 进行单因素方差分析,筛选出CK、Cr_6h 和Cr_6d 细菌群落代谢活性较高的代谢通路,结果发现各组高活性的代谢通路几乎相同。对细菌群落各代谢通路的代谢活性求平均值,列举代谢活性前10 的代谢通路(表2),其中包括全局和总览图(40.81%)、碳水化合物代谢(9.42%)、氨基酸代谢(8.38%)、能量代谢(4.42%)、辅助因子和维生素的代谢(4.24%)、膜运输(3.02%)、翻译(2.65%)、脂质代谢(2.37%)、复制与修复(2.34%)及细胞群落-原核生物(2.31%)。全局和总览图代谢通路主要包括代谢途径、次生代谢产物的生物合成、不同环境中的微生物代谢、碳代谢、2-氧代羧酸代谢、脂肪酸代谢、氨基酸的生物合成和芳香化合物的降解等途径。由表2中数据可知,Cr胁迫对细菌群落主要代谢通路的代谢活性影响不显著。
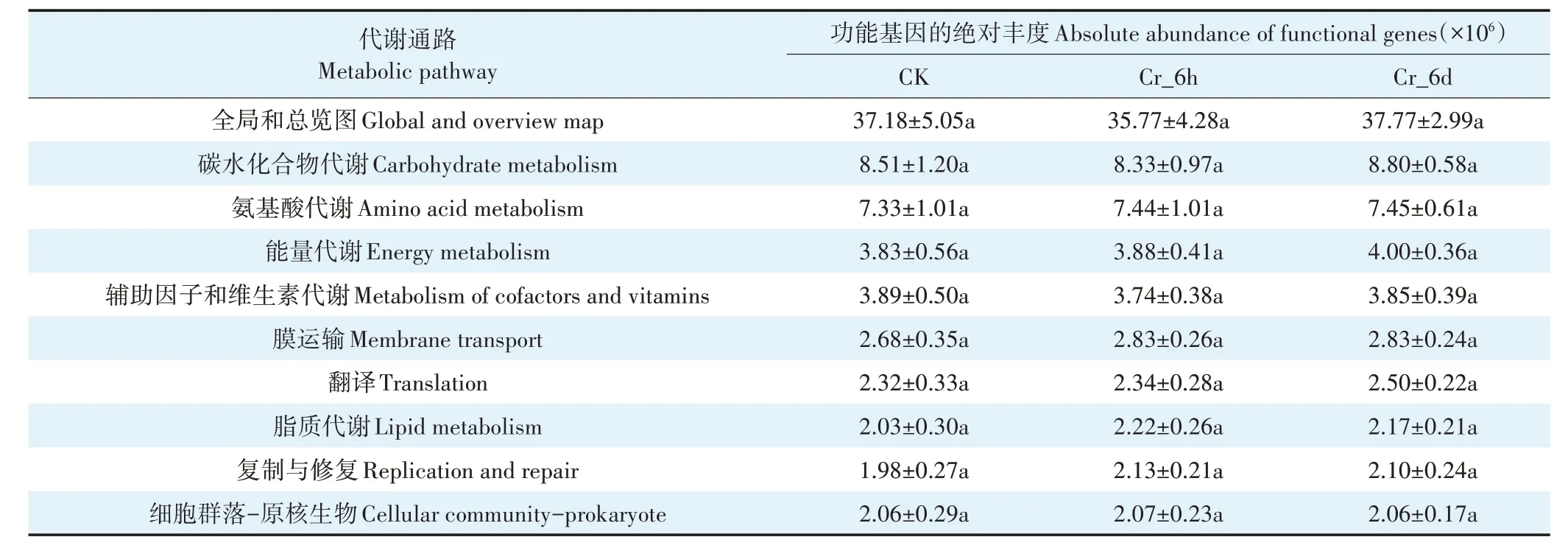
表2 Cr胁迫对谷田土壤细菌群落KEGG代谢活性的影响Table 2 Analysis of soil bacterial communities in the Cr stress time series on KEGG metabolic activity
3 讨论
3.1 谷田土壤细菌群落组成与结构对Cr胁迫的响应
本实验结果发现,Cr 胁迫显著影响了谷田土壤中细菌群落的结构与组成。有研究表明,土壤中重金属超标会导致蛋白质发生变性,改变酶的特异性等过程,进而对微生物的代谢活动产生影响[10,29]。重金属还可能造成微生物的细胞膜损伤,使DNA 结构遭到破坏,进而影响微生物的生长[30-31]。当重金属浓度较高时,还会破坏微生物的群落结构,对微生物生态系统造成严重破坏[11,32]。本研究发现谷田土壤细菌群落主要由放线菌门、厚壁菌门、变形菌门和绿弯菌门等组成,Cr 胁迫显著影响土壤中细菌群落优势菌门的相对丰度(图2A)。Cr胁迫使细菌群落优势菌门中放线菌门和绿弯菌门的相对丰度先显著上升,而后随着Cr 胁迫的持续,优势菌门的相对丰度不再发生显著变化,表明放线菌和绿弯菌对Cr 污染土壤的适应能力相对较强,原因可能是环境中放线菌可通过产生植物激素、抗生素和酶类物质,调整土壤的微生态环境[33]。变形菌门相对丰度随Cr胁迫时间增长先下降,后趋于平稳,表明Cr 胁迫对变形菌产生了一定的毒性影响,但变形菌可根据自身形态的变化来提高对外界环境条件的适应能力[34]。厚壁菌门的相对丰度在Cr_6h 变化较小,之后随着时间增长而下降,表明厚壁菌门对Cr 胁迫有一定的抗性,原因可能是厚壁菌可以产生芽孢,以适应Cr胁迫造成的环境压力[35-36]。
在属水平中(图2B),放线菌门的直丝菌属丰度随Cr 胁迫时间的增长而显著下降,表明直丝菌属对Cr 比较敏感,持续的Cr 胁迫对直丝菌产生了较大的毒害作用。而放线菌门的长孢菌属、马杜拉放线菌属和嗜盐多孔菌属相对丰度在Cr_6h 显著上升、在Cr_6d 有所下降的阶段性变化特征,可能是由于这些菌属对Cr 污染土壤的适应能力相对较强。由此可见,Cr 胁迫改变了谷田土壤细菌群落中耐性菌与敏感菌的比例,使细菌群落的组成与结构发生了改变。如CK、Cr_6h 和Cr_6d 细菌群落中优势菌的相对丰度存在显著差异,表明谷田土壤中的细菌群落在遭受Cr 胁迫后群落结构发生变化,此时群落可能会招募土壤中的耐受菌如放线菌等,通过自身的代谢活动来尽可能地改善土壤环境。本研究结果进一步表明,Cr胁迫后土壤中细菌群落的多样性指数与部分细菌属丰度变化趋势一致,如长孢菌属、马杜拉放线菌属和嗜盐多孔菌属的相对丰度变化趋势与Simpson 指数的变化趋势一致(图2B,图3),表明Cr 污染会对谷田土壤细菌群落的丰度与多样性产生影响。原因可能是重金属会对土壤中细菌群落产生一定的生理生化毒性[8,37-38],因此对Cr 胁迫抗性较低的细菌群落在胁迫条件下很难存活,丰度呈下降趋势,但对Cr 胁迫耐受性较强的细菌群落在胁迫后会呈现阶段性的动态变化过程,而土壤细菌群落多样性的变化可能会影响土壤生态系统的结构与功能。
有研究表明重金属污染会影响微生物的形态与细胞代谢,降低土壤微生物的丰度与多样性[9,29,39-40]。本研究发现,Shannon 指数在Cr_6h 下降,在Cr_6d 又回到CK 水平;Simpson 指数在Cr_6h上升,在Cr_6d又回到CK 水平;而Chao 指数在Cr_6h 下降,在Cr_6d 又开始回升。3 个指数的变化趋势相互验证了谷田土壤细菌群落对Cr 胁迫的应激与响应过程即群落的演变特征,即Cr 胁迫不同时间节点土壤细菌群落多样性变化显著,且呈现阶段性变化特征。Cr 胁迫对谷田土壤中细菌群落产生了一定的毒害作用,显著降低了细菌群落的多样性,但随着Cr 胁迫的持续,群落多样性逐渐开始恢复,表明Cr 胁迫在一定时间范围内对谷田土壤细菌群落造成了一定程度的生态扰动。
3.2 谷田土壤细菌群落构建过程对Cr胁迫的响应
本研究发现,Cr胁迫后谷田土壤细菌群落的betaNTI指数持续下降,在CK 和Cr_6h 阶段土壤细菌群落构建更多地受到确定过程的影响(|betaNTI |>2),而在Cr_6d阶段更多地受到随机过程的驱动(|betaNTI|<2)(图5)。前人研究发现驱动生物群落构建的诸多因素可归纳为选择、扩散、成种/多样化/突变、生态漂变4 个过程[41-42]。选择过程包含环境选择和生物互作两个方面,微生物群落构建与环境因素密切相关,例如:细菌和真菌的群落组成受到空间、土壤、植被、气候等因素的影响[43-44];古细菌群落组成与土壤水分、养分和pH 值等因素密切相关[45];病毒组成则与海拔、土壤pH 值和钙含量等相关[46]。根据相互作用方式的不同,选择过程对微生物群落的影响不同,但普遍认为环境选择对微生物群落的影响是确定性的过程[47]。
研究表明,土壤微生物群落的构建由确定过程与随机过程的权衡作用驱动[48],这是由微生物对不同环境的偏好和适应性造成的。群落构建过程中的随机过程相较于确定过程可产生更多样的生态功能,对驱动农田土壤中细菌群落的组装具有较强的效应。随机过程对可持续农田生态系统中群落功能产生正向反馈机制,缓解重金属胁迫引起的生态扰动,进而维持农田土壤中生态系统的稳定性[49-50]。本研究发现,自然状态下谷田土壤细菌群落的构建过程由确定性过程驱动,Cr 胁迫显著降低细菌群落的多样性,使细菌群落结构遭到破坏,随着Cr 胁迫的持续,群落组成和结构逐渐发生改变,耐受菌的相对丰度增加,以适应胁迫环境,随机过程在群落构建中逐渐占主导地位,使群落朝着产生更多生态功能的方向发展,对维持农田土壤中生态系统的稳定性起正向反馈作用。
3.3 谷田土壤细菌群落的种间关系对Cr胁迫的响应
微生物生态网络属性可表征微生物群落对逆境的反应及相关生态位功能间的关系[51]。在许多生态系统中不同物种之间的相互作用对群落的生态分布格局与群落功能至关重要。胁迫梯度假说认为随着胁迫持续时间或强度的增加,微生物群落中种间竞争关系将减弱,而互利共生关系将增强[52-53]。资源竞争、微生物互养、重金属胁迫等都可促进微生物间的共生合作关系[54-56]。
本研究中共生网络图(图7)表明细菌群落不同属之间存在显著的相互作用,且相互作用多为正相关,节点数量稳定。另外,细菌群落共生网络(表1)的边数量和平均度随着Cr 胁迫时间的延长而下降,细菌群落共生网络的平均路径长度却随着Cr 胁迫时间的延长而上升,表明随着Cr 胁迫处理时间的延长,细菌的网络规模变小,菌群间相互作用关系趋于简单,响应速度变慢,不易受外界环境的干扰。共生网络中直接与间接相互作用对生态系统起着重要调控作用,它们对生态扰动做出同步反应,并产生了积极的反馈和共同振荡[57]。与Cr 胁迫相比,CK 组的共生网络规模更大、相互作用更复杂,部分原因可能是未遭受Cr 胁迫时谷田土壤细菌群落呈稳定状态,物种活性高且种间相互作用复杂,而复杂的网络关系有利于提升群落的稳定性,进而提高资源转移的效率[58];随着Cr 胁迫的持续,细菌群落的多样性降低,响应速度变慢,种间相互作用关系趋于简单,且以共生关系为主导,从而可提高细菌群落对Cr胁迫的适应能力。
4 结论
(1)Cr 胁迫显著影响了谷田土壤中的细菌群落。Cr 胁迫使细菌群落的优势菌门和优势菌属的相对丰度发生改变,细菌群落的多样性变化显著。
(2)随着Cr 胁迫的持续,谷田土壤中细菌群落构建由确定过程驱动逐渐转变为由随机过程驱动。
(3)Cr胁迫使谷田土壤中细菌群落的共生网络规模变小,菌群间响应速度变慢,且以共生关系为主导。
(4)Cr 胁迫对谷田土壤细菌群落主要代谢通路的代谢活性影响较小。
数据可用性声明
文章中的高通量测序原始数据是公开可用的,可在Na-tional Center for Biotechnology Information(https://www.ncbi.nlm.nih.gov/)数据库中下载使用,登录号:PRJNA930506。