苏云金芽孢杆菌鲇泽亚种和中华甲虫蒲螨对冷杉梢斑螟的杀虫作用1)
2023-07-10王倩倩迟德富
王倩倩 迟德富
(东北林业大学,哈尔滨,150040)
冷杉梢斑螟(Dioryctriaabietella)隶属于鳞翅目、螟蛾科,是我国重要的林业害虫,严重危害红松球果[1-2]。目前,受害的红松人工林面积已达到黑龙江省已进入结实期的红松人工林总面积的65%,且持续增加[3-4]。现有的灯光诱杀等物理措施防治冷杉梢斑螟,存在着一次性投入大,且对具有趋光性的各种昆虫没有选择性的缺陷,而化学防治虽能快速有效杀灭害虫,但会给环境和人畜带来威胁。因此,急需探索高效且环保的生物措施防治冷杉梢斑螟。
苏云金芽孢杆菌制剂以高效、无污染、对人无毒而被广泛使用[5]。苏云金芽孢杆菌(Bacillusthuringiensis)是1种革兰氏阳性菌,在芽孢形成期间产生α、β、γ、δ多种类型伴孢晶体蛋白,其中δ-内毒素(亦称为“杀虫晶体蛋白”)是最主要的杀虫蛋白。随着昆虫的取食,晶体蛋白进入昆虫中肠后,在中肠碱性条件下(pH>10),二硫键断开,晶体蛋白溶解成原毒素,在中肠蛋白酶的作用下,原毒素裂解为有活性的毒素,有活性的毒素结合到中肠上皮细胞膜的特异性受体上,毒素亚基低聚形成孔洞结构,并插入到顶膜上,这些孔洞允许离子和水分自由出入细胞,使细胞的通透性受到破坏,形成离子通道或孔洞,引起细胞膨胀和裂解,最后导致昆虫死亡[6-7]。苏云金芽孢杆菌对害虫生长发育和相关酶活的影响已有资料报道[8-9],而有关该菌对鳞翅目冷杉梢斑螟的杀虫毒力以及对该虫生理生化的影响未见报道。为了探索利用苏云金芽孢杆菌防控冷杉梢斑螟的可行性,有必要了解该菌对冷杉梢斑螟幼虫的毒力以及对该虫保护与解毒酶活性的影响。
中华甲虫蒲螨(PyemoteszhonghuajiaYu,Zhang and He)是一种昆虫天敌蒲螨,由于其雌性比例高、完成世代发育时间短、易饲养,叮咬寄主时可产生神经毒素将其麻痹、毒杀,能寄生鞘翅目、鳞翅目、等翅目、膜翅目昆虫,是群居和隐蔽性害虫的重要生物控制因子等特点[10-11],而被广泛应用于害虫的防治。张佐双等[12]应用中华甲虫蒲螨防控六星黑点豹蠹蛾(Zeuzeraleuconotum),寄生率高达60.5%,且蒲螨对其有明显的趋性搜寻现象;2005—2008年,贺丽敏等[13]利用中华甲虫蒲螨控制光肩星天牛(Anoplophoraglabripennis)等钻驻性害虫,寄生率高达98.93%;2012年,贺丽敏等[14]还利用中华甲虫蒲螨控制松梢螟(Dioryctriapryeri),防治效果也达75.90%~86.91%。但应用中华甲虫蒲螨防治冷杉梢斑螟的研究未见报道。鉴于冷杉梢斑螟与松梢螟在分布区域、寄主植物、生物学习性等方面均存在明显差异,因此有必要对中华甲虫蒲螨寄生冷杉梢斑螟的能力开展研究。
因此,本文通过苏云金芽孢杆菌、中华甲虫蒲螨分别作用于冷杉梢斑螟,根据死亡率和保护酶、解毒酶活性的变化,分析两者的杀虫效果,旨在为苏云金芽孢杆菌与中华甲虫蒲螨联合高效安全防治冷杉梢斑螟提供参考。
1 材料与方法
供试虫源:冷杉梢斑螟老熟幼虫,2021年9月采自黑龙江省海林市森宝源天然食品有限公司的红松球果。中华甲虫蒲螨购于南京登博生物科技有限公司。
供试菌及药剂:苏云金芽孢杆菌鲇泽亚种(Bacillusthuringiensissubsp.aizawaiBonnefoi et de Barjac,菌种编号BNCC 110056)购于ATCC菌种库北纳创联生物技术有限公司。将冻干粉于无菌水中溶化,涂布于培养基(蛋白胨5 g,牛肉浸取物3 g,NaCl 5 g,琼脂15 g,蒸馏水1 L)上,培养1 d,菌种稳定后,转接于新的培养基上,培养1~2 d,转接于斜面培养基上,待其稳定后,置于冰箱中4 ℃保存。在超净工作台中,用无菌枪头将菌苔刮到装有250 mL LB液体培养基的锥形瓶中,将锥形瓶置于恒温振荡器中,在30 ℃、170 r/min条件培养48 h,获得孢晶混合物,用体积分数为0.05%的吐温-80制备不同浓度菌悬液。
试验仪器:蓝豹恒温振荡器HZQ-300;致微全自动高压灭菌器FD50A;北京东联哈尔(哈东联)洁净工作台;北京东联哈尔(哈东联)智能光照培养箱HPG-400BX;杭州奥盛全波长酶标仪FlexA-200;艾本德5430/R小型台式高速冷冻离心机;体视显微镜Leica M205FA。
苏云金芽孢杆菌鲇泽亚种对冷杉梢斑螟老熟幼虫的生物测定:将苏云金芽孢杆菌鲇泽亚种菌悬液制成不同浓度,将相同大小的块状饲料在不同浓度菌悬液中充分浸蘸,虫饥饿24 h后,放入方形养虫盒中,分别饲喂24 h,更换新的无菌饲料。对照用蒸馏水浸蘸。处理后的试虫移到养虫室饲养。饲养条件:温度(23±1)℃,相对湿度70%左右,光周期为14 h光照、10 h黑暗。每组3个重复,每个重复20头虫。每天检查死虫数,连续检查11 d。
中华甲虫蒲螨对冷杉梢斑螟老熟幼虫的生物测定:将饥饿24 h的冷杉梢斑螟老熟幼虫置于直径为35 mm的塑料培养皿中,每皿1头,用0号毛笔将中华甲虫蒲螨挑出,在解剖镜下分区计数。将0(CK)、20、30、40、50、60头蒲螨分别接到虫体,用保鲜膜封口,昆虫针在保鲜膜上均匀扎孔,再在保鲜膜上盖上培养皿盖,防止冷杉梢斑螟咬破保鲜膜逃逸。每皿1头幼虫,作为一个重复,每个处理10个重复。最后将接入蒲螨的塑料培养皿全部放进智能光照培养箱(温度(25±1)℃,相对湿度55%)中。每天在解剖镜下观察记录冷杉梢斑螟的存活情况(用针刺虫体不动即为死亡),以及蒲螨寄生状况(产生膨腹体的大小、数量)。
酶源的制备:将苏云金芽孢杆菌鲇泽亚种菌悬液制成不同浓度,将相同大小的块状饲料在不同浓度菌悬液中充分浸蘸,虫饥饿24 h后,放入方形养虫盒中,分别饲喂浸蘸菌悬液后的饲料4、8、12、24、48 h后取活虫,浇液氮,置于冰箱中-80 ℃冷冻待用,对照(用浸蘸蒸馏水后的饲料饲喂冷杉梢斑螟老熟幼虫)同上。每组3个重复。将待匀浆的幼虫,用流动的蒸馏水冲洗后,放在滤纸上吸干水分,用液氮快速冷却,置于匀浆器中,放入1 mL酶提取液,冰浴匀浆,匀浆液于高速冷冻离心机上以4 ℃、12 000 r/min离心10 min,取上清液作为酶源,置冰上待用。
蒲螨处理组,将不同数量中华甲虫蒲螨接到冷杉梢斑螟幼虫体后,分别于4、8、12、24、48 h取出冷杉梢斑螟幼虫,浇液氮,置于冰箱中-80 ℃冷冻待用,对照同上。每组10个重复。后续操作同苏云金芽孢杆菌鲇泽亚种菌悬液处理组。
乙酰胆碱酯酶(AchE)、羧酸酯酶(CarE)、谷胱甘肽-S-转移酶(GSTs)、过氧化氢酶(CAT)活性的测定,参照格锐思试剂盒说明书进行;过氧化物酶(POD)活性的测定,参照苏州科铭试剂盒说明书进行。
数据处理:本试验采用Excel和SPSS 23.0软件中的单因素方差分析进行死亡率和酶活性的显著性分析,通过概率单位回归计算苏云金芽孢杆菌鲇泽亚种对冷杉梢斑螟的致死中浓度(LC50)。
2 结果与分析
2.1 苏云金芽孢杆菌鲇泽亚种对冷杉梢斑螟老熟幼虫的致死力测定
处理分析取食浸蘸不同菌浓度苏云金芽孢杆菌鲇泽亚种的饲料后冷杉梢斑螟老熟幼虫的死亡数,拟合得到苏云金芽孢杆菌鲇泽亚种对冷杉梢斑螟老熟幼虫的致死中浓度为6 584.065菌落/mL,毒力回归方程为y=0.431x-1.286。表1表明,冷杉梢斑螟老熟幼虫的死亡率随苏云金芽孢杆菌鲇泽亚种菌浓度的提高而升高。
苏云金芽孢杆菌鲇泽亚种对冷杉梢斑螟老熟幼虫的毒力较强。与CK相比,冷杉梢斑螟取食苏云金芽孢杆菌鲇泽亚种初期,幼虫拒食,行动迟缓。取食后期,虫体颜色加深,逐渐变为黑色,虫体逐渐瘫软,失去弹性,用镊子难以夹起,体壁皱缩。死亡虫体发黑,腐烂发臭(图1)。
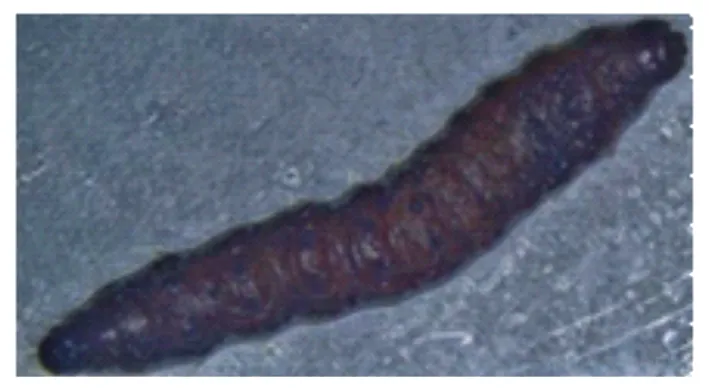
图1 苏云金芽孢杆菌鲇泽亚种致死冷杉梢斑螟老熟幼虫
2.2 不同菌浓度苏云金芽孢杆菌鲇泽亚种处理冷杉梢斑螟的保护酶和解毒酶的活性变化
取食经105、106、107、108、109个/mL苏云金芽孢杆菌鲇泽亚种处理后饲料的冷杉梢斑螟老熟幼虫,乙酰胆碱酯酶活性多显著高于取食相同时间CK处理的个体(P<0.05,表2)。只有取食经105个/mL苏云金芽孢杆菌鲇泽亚种处理饲料的4、8、24 h时,经106、107个/mL菌液处理饲料的4 h时,经109个/mL菌液处理饲料的4、12、48 h时的乙酰胆碱酯酶活性与CK差异不显著,但依然略高于CK,表明乙酰胆碱酯酶活性被诱导激活。梢斑螟在取食含苏云金芽孢杆菌鲇泽亚种的饲料后,随苏云金芽孢杆菌鲇泽亚种毒素的释放以及作用时间的延长,解毒酶乙酰胆碱酯酶被诱导激活,为维持正常生理活动,乙酰胆碱酯酶提高活性,分解毒素,起到解毒作用。
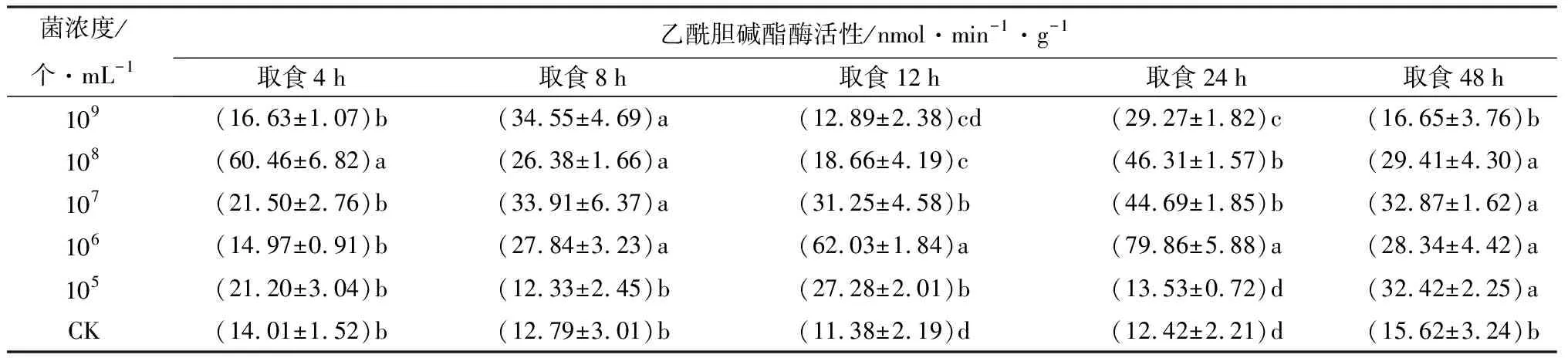
表2 不同菌浓度苏云金芽孢杆菌鲇泽亚种处理冷杉梢斑螟后乙酰胆碱酯酶活性
取食经105、106、107、108、109个/mL苏云金芽孢杆菌鲇泽亚种浸蘸后的饲料4、8、12、24、48 h时,羧酸酯酶活性均显著低于取食相同时间CK处理的个体(P<0.05,表3),表明羧酸酯酶活性被显著抑制。苏云金芽孢杆菌鲇泽亚种通过抑制羧酸酯酶活性,阻碍羧酸酯酶发挥解毒作用,从而扰乱梢斑螟正常生理代谢,最终导致死亡。
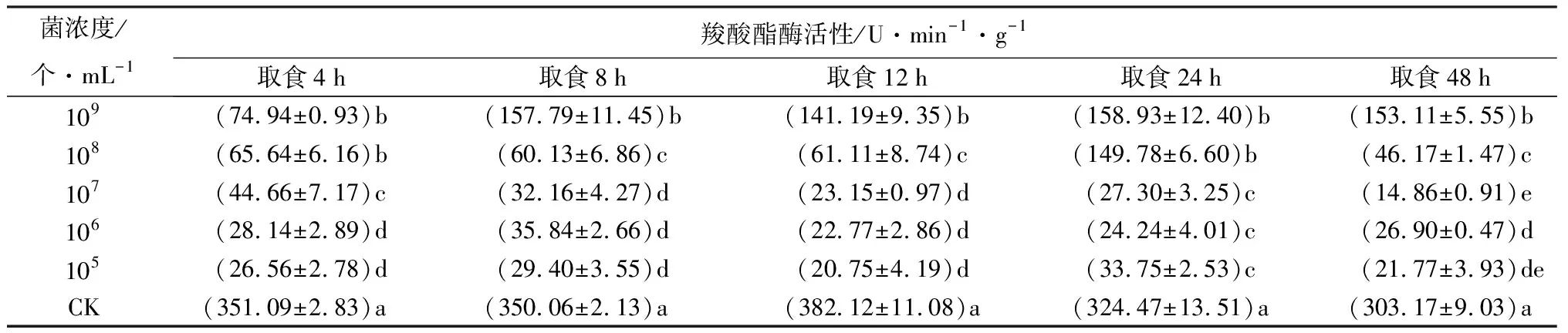
表3 不同菌浓度苏云金芽孢杆菌鲇泽亚种处理冷杉梢斑螟后羧酸酯酶活性
取食经105、106、107、108、109个/mL苏云金芽孢杆菌鲇泽亚种浸蘸的饲料后,谷胱甘肽-S-转移酶活性大多高于取食相同时间CK处理的个体(表4),只有取食经105个/mL菌液处理饲料的12、48 h,及经106、107个/mL菌液处理饲料的48 h,冷杉梢斑螟谷胱甘肽-S-转移酶活性与CK差异不显著。表明谷胱甘肽-S-转移酶活性被诱导激活。说明梢斑螟在取食含苏云金芽孢杆菌鲇泽亚种的饲料后,芽孢和晶体在中肠中特定pH时释放毒蛋白,在中肠蛋白酶作用下水解成毒素,毒素在虫体内不断累积,为维持昆虫正常生理活动,解毒酶谷胱甘肽-S-转移酶活性升高,起到解毒作用。
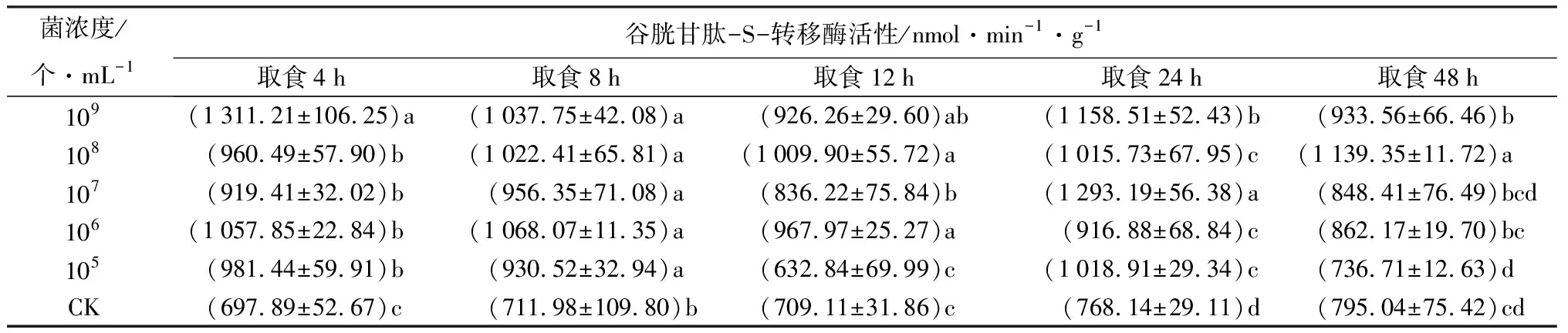
表4 不同菌浓度苏云金芽孢杆菌鲇泽亚种处理冷杉梢斑螟后谷胱甘肽-S-转移酶活性
取食经105个/mL苏云金芽孢杆菌鲇泽亚种处理饲料的冷杉梢斑螟老熟幼虫,4、12、24、48 h时,体内过氧化氢酶活性低于CK,8 h时显著高于CK(P<0.05),为CK的1.11倍(表5)。表明取食经105个/mL苏云金芽孢杆菌鲇泽亚种处理的饲料时,过氧化氢酶活性总体呈抑制-激活-抑制趋势。取食经106个/mL菌液处理饲料,4、12、24 h时,梢斑螟过氧化氢酶活性显著低于CK(P<0.05);8、48 h时,显著高于CK(P<0.05),分别为CK的1.13、1.04倍。表明取食经106个/mL苏云金芽孢杆菌鲇泽亚种处理的饲料时,过氧化氢酶活性总体呈抑制-激活-抑制-激活趋势。取食经107个/mL菌液处理饲料,4、8、24 h时,过氧化氢酶活性显著高于CK(P<0.05),分别为CK的1.04、1.03、1.01倍;12、48 h时,显著低于CK(P<0.05)。表明取食经107个/mL苏云金芽孢杆菌鲇泽亚种处理的饲料时,过氧化氢酶活性总体呈激活-抑制-激活-抑制的趋势。取食经108、109个/mL菌液处理饲料的虫体内过氧化氢酶活性均显著低于CK(P<0.05),表明过氧化氢酶活性被明显抑制。过氧化氢酶活性增强,分解H2O2,防止产生自由基毒害。梢斑螟在取食经105个/mL苏云金芽孢杆菌鲇泽亚种处理的饲料后,芽孢和晶体在中肠中特定pH释放毒蛋白,8 h时形成晶体,在中肠蛋白酶作用下水解成毒素,毒素在虫体内不断累积,过氧化氢酶被激活,后期过氧化氢酶活性被抑制。取食106个/mL苏云金芽孢杆菌鲇泽亚种处理的饲料后,初期,梢斑螟虫体过氧化氢酶活性被激活,随着毒素积累,后期再次被激活。取食经107个/mL苏云金芽孢杆菌鲇泽亚种处理的饲料后,过氧化氢酶活性呈激活-抑制-激活-抑制的趋势,表明虫体被该菌浓度处理后,酶活性较敏感,到48 h时,虫体活性降低,趋于死亡,过氧化氢酶活性也降低。取食经108、109个/mL苏云金芽孢杆菌鲇泽亚种处理的饲料后,虫体内过氧化氢酶活性始终低于CK,取食前期,苏云金芽孢杆菌鲇泽亚种抑制过氧化氢酶活性,阻碍过氧化氢酶发挥保护作用,甚至无法发挥作用,导致保护酶系紊乱,扰乱虫体正常生理机能,最终导致虫体死亡,过氧化氢酶活性也降低。
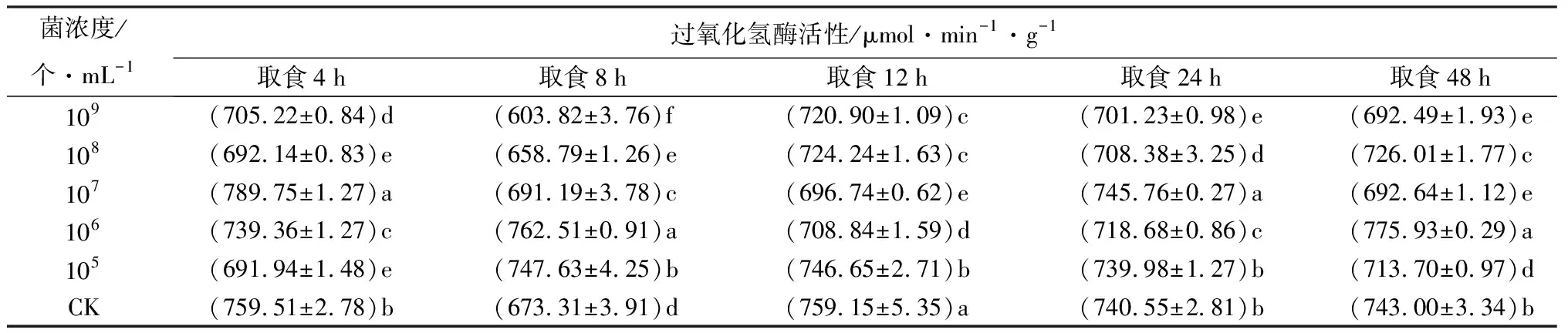
表5 不同菌浓度苏云金芽孢杆菌鲇泽亚种处理冷杉梢斑螟后过氧化氢酶活性
取食经105个/mL苏云金芽孢杆菌鲇泽亚种处理后饲料的冷杉梢斑螟老熟幼虫,4、8 h时,体内过氧化物酶活性显著低于CK(P<0.05,表6);12、24 h时,与CK差异不显著;48 h时显著高于CK,为CK的1.18倍(P<0.05)。取食经106个/mL苏云金芽孢杆菌鲇泽亚种处理后饲料,8、24 h时,虫体内过氧化物酶活性显著低于CK(P<0.05);12 h时,过氧化物酶活性达到峰值,为CK的1.25倍;4、48 h时,与CK差异不显著。表明取食经106个/mL苏云金芽孢杆菌鲇泽亚种处理后饲料时,过氧化物酶活性总体呈抑制-激活-抑制的趋势。取食经107个/mL苏云金芽孢杆菌鲇泽亚种浸蘸饲料的冷杉梢斑螟老熟幼虫,4、48 h时,过氧化物酶活性显著低于CK(P<0.05);8、12 h时,过氧化物酶活性显著高于CK,8 h时达到峰值,为CK的1.12倍(P<0.05);24 h时,过氧化物酶活性与CK差异不显著。表明取食经107个/mL苏云金芽孢杆菌鲇泽亚种处理后饲料时,过氧化物酶活性总体呈抑制-激活-抑制的趋势。取食经108、109个/mL苏云金芽孢杆菌鲇泽亚种处理后饲料,虫体内过氧化物酶活性多显著低于取食相同时间CK处理的个体(P<0.05),只有经108个/mL菌悬液处理后饲料的4、12、24 h时,与CK差异不显著,表明过氧化物酶活性多被抑制。过氧化物酶主要作用是清除体内过量的H2O2,维持细胞内H2O2的平衡,从而适应外界环境。低菌浓度(105个/mL)的苏云金芽孢杆菌鲇泽亚种处理中,随毒素积累,直到48 h时,过氧化物酶被激活,开始清除体内多余自由基,降低毒性。106、107个/mL菌悬液处理的虫体内过氧化物酶活性变化趋势表明,随苏云金芽孢杆菌鲇泽亚种菌浓度的升高,毒素在虫体累积,为维持昆虫正常生理生化反应,过氧化物酶被激活的时间点提前,随作用时间的延长,毒素在虫体内进一步累积,彻底破坏虫体保护酶系,酶活性降低,虫体活性降低,直至死亡。高菌浓度(108、109个/mL)苏云金芽孢杆菌鲇泽亚种抑制虫体内的过氧化物酶活性,阻碍过氧化物酶发挥作用,导致保护酶系功能紊乱,最终导致死亡,过氧化物酶活性也降低。

表6 不同菌浓度苏云金芽孢杆菌鲇泽亚种处理的冷杉梢斑螟过氧化物酶活性
2.3 中华甲虫蒲螨对冷杉梢斑螟老熟幼虫的致死力测定
处理分析接入不同数量中华甲虫蒲螨的冷杉梢斑螟老熟幼虫的死亡数,拟合得到中华甲虫蒲螨寄生冷杉梢斑螟老熟幼虫的半数致死数量(27.454头),毒力回归方程为y=0.032x-0.866。经接入数量为20、30、40、50、60、0(CK)头中华甲虫蒲螨处理后,冷杉梢斑螟老熟幼虫的死亡率分别为40.00%、60.00%、60.00%、70.00%、90.00%、0,表明冷杉梢斑螟老熟幼虫的死亡率随中华甲虫蒲螨接入量的增加而升高。
由表7可见,中华甲虫蒲螨对冷杉梢斑螟老熟幼虫的寄生能力较强,接入中华甲虫蒲螨60头,冷杉梢斑螟老熟幼虫死亡率高达90.00%。
2.4 中华甲虫蒲螨寄生后冷杉梢斑螟性状
中华甲虫蒲螨寄生冷杉梢斑螟致死后,虫体死状多为“C”形或“I”形。梢斑螟虫体刚刚死亡时,体壁遍布蒲螨刺吸叮咬留下的黑斑(图2A);死后0~2d,虫体上的蒲螨产生膨腹体,随膨腹体膨大,产生小螨,虫体逐渐干瘪,变硬(图2B、C)。
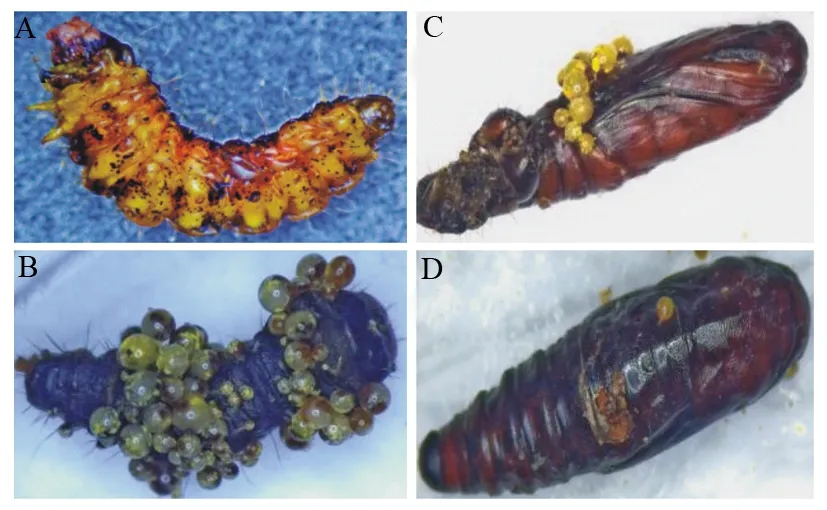
A为中华甲虫蒲螨寄生后冷杉梢斑螟死状;B为中华甲虫蒲螨寄生冷杉梢斑螟产生膨腹体;C为中华甲虫蒲螨寄生虫蛹产生膨腹体;D为中华甲虫蒲螨寄生化蛹完全的虫蛹产生膨腹体。
经过24 d的观测,中华甲虫蒲螨既可以寄生幼虫,也可以寄生蛹。幼虫在接入蒲螨后2.7~14.7 d,蒲螨产生膨腹体;在开始化蛹后2 d,蛹上的蒲螨产生膨腹体。在冷杉梢斑螟化蛹初期,寄生效率更高。蒲螨寄生部位偏好冷杉梢斑螟的头部和节间膜,产生膨腹体的位置比较分散,遍布全身。蒲螨寄生虫蛹,多在虫蛹还未完全闭合处(第4腹节处)寄生,因此产生膨腹体的位置比较集中。中华甲虫蒲螨也可寄生已经化蛹完全的虫蛹,从图2D可见,接入蒲螨后,直至幼虫化蛹,虫蛹被蒲螨叮咬出1个明显的孔洞,旁边有膨腹体产生。试验发现,虫体上的蒲螨产生膨腹体,虫体即死亡,也有虫死后2 d,虫体上的蒲螨产生膨腹体,膨腹体直径平均每天增长0.01~0.02 mm,1 d最大增长0.13 mm,幼虫产膨腹体直径最大可达0.88 mm,蛹产膨腹体直径最大可达1.05 mm,持续观察发现,产小螨的膨腹体的最小直径为0.6 mm,最大直径为1.05 mm。
2.5 中华甲虫蒲螨对冷杉梢斑螟保护酶和解毒酶活性的影响
接20、30、40、50、60头中华甲虫蒲螨到冷杉梢斑螟老熟幼虫后,4、8、12、24、48 h时,梢斑螟虫体内乙酰胆碱酯酶活性多显著高于CK(P<0.05,表8),只有接50头的4、48 h和接20头的48 h,乙酰胆碱酯酶活性与CK差异不显著。表明乙酰胆碱酯酶活性整体处于被诱导激活状态。梢斑螟在被蒲螨叮咬、刺吸后,随寄生时间的延长,乙酰胆碱酯酶活性不断升高,起到降低体内毒性的作用,随接入数量增多(50、60头)以及接入时间的延长(48 h),虫体进入深度麻痹状态,免疫系统紊乱,虫体活性降低,酶活性也降低。

表8 接入不同数量中华甲虫蒲螨的冷杉梢斑螟乙酰胆碱酯酶活性
接20头中华甲虫蒲螨到冷杉梢斑螟老熟幼虫,48 h时,梢斑螟虫体内羧酸酯酶活性显著高于CK,为CK的1.24倍(P<0.05,表9);处理8、12、24 h时,羧酸酯酶活性显著低于CK(P<0.05);处理4 h时,羧酸酯酶活性与CK差异不显著。表明接20头中华甲虫蒲螨时,虫体内羧酸酯酶活性总体呈抑制-激活的趋势。接30、40、50、60头中华甲虫蒲螨,4、8、12、24、48 h时,虫体羧酸酯酶活性均低于CK,只有接30头中华甲虫蒲螨4h时、接60头中华甲虫蒲螨48 h时,与CK差异不显著,表明羧酸酯酶活性多被抑制。接入少量蒲螨(20头),蒲螨刺吸、叮咬虫体,注入神经毒素,毒素不断积累,直到48 h,羧酸酯酶活性被激活。随接入数量的增加和接入时间的延长,虫体在短时间内迅速麻痹且麻痹程度不断加深,体内免疫系统逐渐紊乱,虫体活性降低,羧酸酯酶活性也降低。
接20、30、40、50、60头中华甲虫蒲螨到冷杉梢斑螟老熟幼虫4、8、12、24、48 h时,梢斑螟虫体内谷胱甘肽-S-转移酶活性多高于CK处理相同时间的个体(表10),只有接20、30头中华甲虫蒲螨8 h时、接40头中华甲虫蒲螨8、12、48 h时和接50头中华甲虫蒲螨8、12、24 h时,谷胱甘肽-S-转移酶活性与CK差异不显著。表明谷胱甘肽-S-转移酶活性总体呈激活的趋势。蒲螨叮咬梢斑螟体壁,同时注入毒素,激活谷胱甘肽-S-转移酶活性,谷胱甘肽-S-转移酶发挥解毒作用,维持昆虫正常生理活动。

表10 接入不同数量中华甲虫蒲螨的冷杉梢斑螟谷胱甘肽-S-转移酶活性
接20、30头中华甲虫蒲螨到冷杉梢斑螟老熟幼虫,24 h时,虫体内过氧化氢酶活性显著高于CK(P<0.05),分别为CK的1.04、1.02倍(表11);4、8、12、48 h时,过氧化氢酶活性多数显著低于CK(P<0.05),只有接20头中华甲虫蒲螨48 h时,过氧化氢酶活性与CK差异不显著。表明接20、30头中华甲虫蒲螨时,过氧化氢酶活性总体呈抑制-激活-抑制的趋势。接40、50头中华甲虫蒲螨到冷杉梢斑螟老熟幼虫,4、12、24、48 h时,虫体内过氧化氢酶活性显著低于CK(P<0.05);8 h时,过氧化氢酶活性显著高于CK(P<0.05),分别为CK的1.09、1.05倍。表明接40、50头中华甲虫蒲螨时,过氧化氢酶活性总体呈抑制-激活-抑制的趋势。随接入蒲螨数量的增加以及接入时间的延长,虫体内毒素累积,过氧化氢酶被激活的时间点提前,过氧化氢酶可以分解体内多余的自由基,降低毒性,从而维持虫体正常生理活动。接入后期,毒素继续在虫体内积累,虫体进入深度麻痹,活性降低,过氧化氢酶活性也降低。接60头中华甲虫蒲螨4、8、12、24、48 h时,过氧化氢酶活性显著低于CK(P<0.05),表明过氧化氢酶活性被明显抑制。由于接蒲螨头数较多,注入虫体毒素也相应增多,虫体迅速进入麻痹状态,蒲螨不断吸取虫体内的营养物质,虫活性降低直至死亡,过氧化氢酶活性也降低。
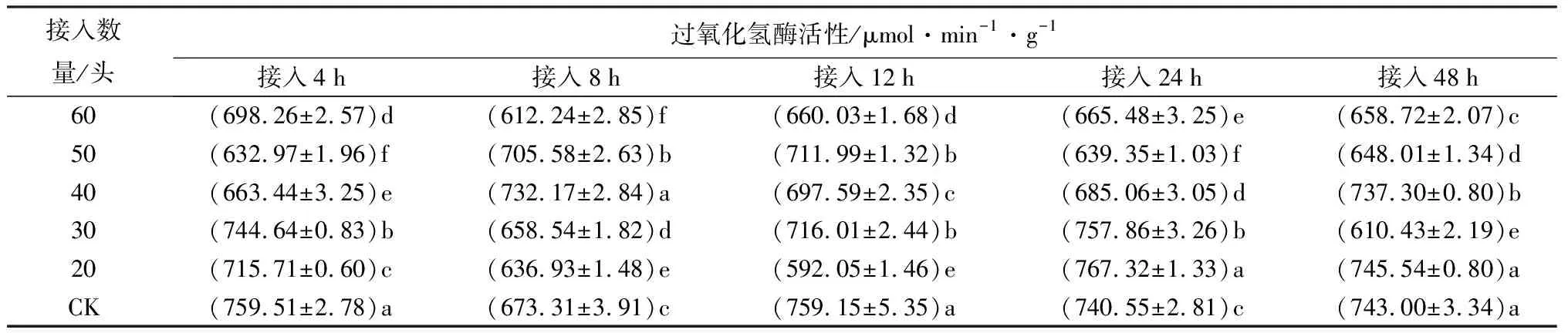
表11 接入不同数量中华甲虫蒲螨的冷杉梢斑螟过氧化氢酶活性
接20头中华甲虫蒲螨到冷杉梢斑螟老熟幼虫,48 h时,虫体内过氧化物酶活性显著高于CK(P<0.05),为CK的1.05倍(表12);4、8、12、24 h时,过氧化物酶活性显著低于CK(P<0.05)。表明接20头中华甲虫蒲螨时,过氧化物酶活性总体呈抑制-激活的趋势。接30头中华甲虫蒲螨处理,4、48 h时,虫体内过氧化物酶活性显著高于CK(P<0.05),分别为CK的1.73、1.22倍;8、12、24 h时,过氧化物酶活性显著低于CK(P<0.05)。表明接30头中华甲虫蒲螨时,过氧化物酶活性总体呈激活-抑制-激活的趋势。接40、50、60头中华甲虫蒲螨4、8、12、24、48 h时,虫体过氧化物酶活性均显著低于处理相同时间CK的个体(P<0.05),表明接40、50、60头中华甲虫蒲螨时,虫体过氧化物酶活性被明显抑制。接20头蒲螨时,随接入时间的延长,毒素在虫体内积累,过氧化物酶活性在后期被激活;接30头蒲螨,初期和后期过氧化物酶活性增强,分解虫体内多余的自由基,降低毒性,起到保护作用。当接蒲螨数量增多时,毒素在虫体内迅速积累,虫体在较短时间内麻痹,体内保护酶系逐渐被破坏,蒲螨取食虫体内营养物质,过氧化物酶活性始终低于对照。
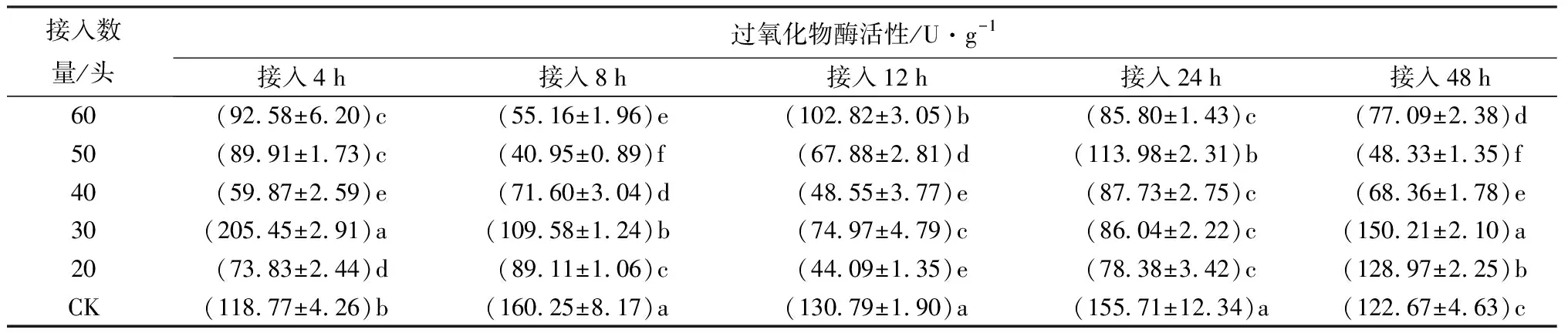
表12 接入不同数量中华甲虫蒲螨的冷杉梢斑螟过氧化物酶活性
3 结论与讨论
苏云金芽孢杆菌杀虫晶体蛋白具有高度专一性、只对靶标害虫有效、对其他昆虫无害、环保等特点[15]。本文采用饲料浸蘸苏云金芽孢杆菌鲇泽亚种的方法,室内处理冷杉梢斑螟,取得了良好的效果,且随着苏云金芽孢杆菌鲇泽亚种菌浓度的升高,梢斑螟的死亡率上升,菌浓度为109个/mL时,致死率最高(100%)。中华甲虫蒲螨毒力很强,繁殖周期短,被称为“蛀虫杀手”[11-12]。梢斑螟类害虫由于幼虫营隐蔽性生活,被害果梢产生保护性泌脂反应,常规药剂防治难以接触虫体[16],本文采用蒲螨防治,可达到更好的防治效果。随蒲螨接入数量的增加,梢斑螟死亡率升高,室内试验杀虫效果高达90%。中华甲虫蒲螨在冷杉梢斑螟老熟幼虫虫体上寄生效果较好。
昆虫对周围环境的变化极其敏感,环境胁迫会引起昆虫体内一系列生理变化,并伴随着内源毒素的产生。昆虫解毒酶系(如羧酸酯酶、乙酰胆碱酯酶、谷胱甘肽-S-转移酶等)的活性可以被各种外源、内源化合物诱导,这使得昆虫在受到环境胁迫时,体内酶能迅速作出反应,从而存活下来[17]。
Guo et al.[18]认为,羧酸酯酶是昆虫体内一类重要的解毒酶系,在内源或外源毒素代谢中不需要额外能量就能够催化酯类化合物水解,所以昆虫在不同环境胁迫时,均能通过提高自身羧酸酯酶活性来适应逆境。本研究中,经过苏云金芽孢杆菌鲇泽亚种或中华甲虫蒲螨处理后,羧酸酯酶活性多低于CK,呈现被抑制状态。表明苏云金芽孢杆菌鲇泽亚种和中华甲虫蒲螨都对冷杉梢斑螟羧酸酯酶活性有抑制作用。
取食经不同菌浓度苏云金芽孢杆菌鲇泽亚种处理的饲料和接入不同数量中华甲虫蒲螨的冷杉梢斑螟老熟幼虫,乙酰胆碱酯酶活性均显著高于CK,表明乙酰胆碱酯酶活性均被明显激活。苏云金芽孢杆菌鲇泽亚种处理后,虫体内谷胱甘肽-S-转移酶活性均高于CK。接不同数量中华甲虫蒲螨到冷杉梢斑螟,谷胱甘肽-S-转移酶活性均高于CK,表明蒲螨寄生虫体后,酶活性被显著激活。这是因为蒲螨寄生虫体后,注入神经毒素,虫体产生免疫反应,体内解毒酶系活性升高,发挥解毒作用,以维持昆虫正常生理活动。
超氧化物歧化酶、过氧化氢酶和过氧化物酶是生物体内普遍存在的3种防御氧化损伤的重要保护酶[19-20]。昆虫在环境胁迫时,体内活性氧的质量分数将增多,过氧化氢酶能将高浓度的有害物质H2O2分解为水和O2,从而阻止形成毒性更强的羟基自由基,将昆虫体内的H2O2控制在较低的水平,维持昆虫正常的生理活动。本研究中,苏云金芽孢杆菌鲇泽亚种处理的虫体,过氧化氢酶活性多呈抑制-激活-抑制的趋势,表明梢斑螟幼虫在取食初期,体内过氧化氢酶活性被显著抑制;取食中期,随毒素积累,过氧化氢酶被激活;取食后期,随毒素浓度积累以及作用时间的延长,超过过氧化氢酶解毒能力,虫体正常生理功能受到影响,取食递减,虫活性降低,酶活性降低。随蒲螨接入数量的增多,虫体过氧化氢酶被激活的时间点提前,当继续增加接入数量,虫体短时间内,迅速麻痹,酶活性被抑制,虫活性随之降低。
过氧化物酶是一类以H2O2为电子受体催化底物的氧化还原酶,在昆虫体内可以有效催化H2O2、氧化酚类和胺类化合物,从而起到清除H2O2和酚类、胺类毒性的双重作用[21]。本研究中,取食经苏云金芽孢杆菌鲇泽亚种处理后饲料的冷杉梢斑螟,过氧化物酶呈不同的变化趋势。高菌浓度苏云金芽孢杆菌鲇泽亚种处理的虫体,过氧化物酶活性始终低于CK;较低菌浓度苏云金芽孢杆菌鲇泽亚种处理前期,过氧化物酶活性增强,苏云金芽孢杆菌鲇泽亚种蛋白激发了虫体内解毒酶活性,过氧化物酶活性升高,起到保护作用,处理后期,过氧化物酶活性降低。蒲螨处理后,冷杉梢斑螟幼虫体内过氧化物酶呈相同的变化趋势。接入初期(4 h),酶活性被激活,起保护作用,随接入数量的增加以及接入时间的延长,毒素在体内累积,蒲螨吸食虫体血淋巴以及营养物质,虫体逐渐麻痹,虫活性降低,过氧化物酶活性也逐渐降低。
综上所述,苏云金芽孢杆菌鲇泽亚种作用于冷杉梢斑螟,对幼虫体内的解毒酶(乙酰胆碱酯酶、谷胱甘肽-S-转移酶)均有激活作用;对羧酸酯酶有抑制作用;过氧化氢酶和过氧化物酶随苏云金芽孢杆菌鲇泽亚种处理浓度的升高以及作用时间的延长,表现出逐渐抑制的作用。菌浓度为109个/mL的苏云金芽孢杆菌鲇泽亚种致死率最高,幼虫死亡率高达100%。中华甲虫蒲螨处理,对虫体内乙酰胆碱酯酶、谷胱甘肽-S-转移酶均有激活作用;虫螨数量比为1∶20、1∶30、1∶40、1∶50时,对冷杉梢斑螟幼虫过氧化氢酶活性表现出抑制-激活-抑制的作用,虫螨数量比为1∶60时,对过氧化氢酶活性表现为全部抑制作用;对羧酸酯酶和过氧化物酶多表现出抑制作用。接60头蒲螨到虫体的致死率最高,幼虫死亡率高达90%。
