西藏不同林龄核桃林土壤理化性质及林下植被多样性特征1)
2023-07-10张绍聪高禾苗袁龙义任爱天
张绍聪 高禾苗 袁龙义 任爱天
(长江大学,荆州,434025)
林下植被是森林生态系统的重要组成部分,在改良土壤、促进养分循环、提高林地生物多样性、维持林地生态系统结构及功能等方面起着重要作用[1]。林下灌木、草本通过固氮及加快土壤养分循环的方式来改善土壤养分状况,其对林地的覆盖也能有效阻止雨水对林地的侵蚀,显著减少土壤水土及养分流失[2]。此外,灌木、草本的存在也可以增加土壤孔隙度,对土壤微生物及植物根系的生长、雨水入渗具有积极作用[3-4]。然而,相关研究表明,人工林林下植被物种多样性远低于天然林。由于人工林群落组成结构简单,外加不同程度的扰动导致其有机质质量分数和土壤酶活性降低,影响林下植被多样性和生态功能,因此造成人工经济林生态系统退化、林木生产力降低、病害严重。
核桃(JuglansregiaL.)是胡桃科胡桃属植物,属温带树种,其树体高大、枝叶繁茂、根系发达,在净化空气、涵养水源、保持水土等方面有不可忽视的作用[5]。西藏是中国六大核桃分布区之一,适宜的水热和光照资源使核桃在西藏南部和中部地区广泛分布。该地区核桃林种质资源丰富、种类繁多、林龄古老,是西藏特色人工经济林之一[6]。有研究表明,同种类人工林在长期培育过程中,随着林龄的增加,林地土壤有机质、全氮、全磷质量分数均呈先增大后减小的趋势[7],且养分的变化显著影响林下植被的组成和多样性[8]。此外,也研究表明,林下植被的发育与物种多样性、林分立地条件、林龄、林分密度密切相关[9]。但目前关于西藏高原核桃林龄对其林下植被及根际土壤养分、酶活性影响的相关研究较少,且鲜有对中龄林及古核桃林林下植被多样性、土壤理化性质的变化以及二者之间相互影响的研究。因此,本研究对西藏高原加查县不同林龄核桃林根际土壤理化性质的变化规律进行分析,同时对中龄林及古核桃林的林下植被多样性及土壤理化特性进行研究,探究了林下植被群落与土壤理化特性间的关系,为高原核桃林经济功能、生态功能的充分发挥提供参考。
1 研究区概况
研究区域位于西藏自治区加查县(92°14′43″~93°7′10″E,28°49′43″~29°43′16″N),年均气温8.8 ℃,年均日照时间2 700 h,年均降水量485 mm,主要集中在5月,年均蒸发量2 055.5 mm,平均海拔3 200 m。土壤母质为砂岩,土壤层厚度一般为60~100 cm,pH在6.4~7.5。该地区主要树种为核桃、光核桃(PrunusmiraKoehne.)、银白杨(PopulusalbaL.)等。幼龄林林下植被为薹草(CarexLinn.);中龄林及古核桃林林下灌草植物主要为草麻黄(EphedrasinicaStapf.)、白莲蒿(ArtemisiastechmannianaBesser.)、矮生薹草(CarexpumilaThunb.)、牛蒡(ArctiumlappaL.)、委陵菜(PotentillachinensisSer.)、早熟禾(PoaannuaL.)等。
2 研究方法
2.1 样地设置
在西藏加查县选取核桃资源较丰富的3个区域(热推村、拉岗村、九堆村)设置样地,具体见表1。在每个区域选择立地条件、坡度基本一致的50 m×50 m人工幼龄林(林龄为0~10 a)、80 m×80 m人工中龄林(林龄为10~50 a)、100 m×100 m近自然林的古核桃林(林龄为100 a以上),共3个样地,并分别在每个样地随机设置3个采样点,采集0~20 cm土层土样,用密封袋封存,带回实验室进行风干处理。由于幼龄林林下植被只有少量覆盖的薹草属(1种),因此,本研究只对中龄林及古核桃林林分特征(如胸径、盖度、活下枝高度、郁闭度等)进行调查。分别在每个区域样地的中龄林设置11个、古核桃林设置15个25 m×25 m的代表性大样方,对3个区域的中龄林和古核桃林样地林下植被多样性进行分析。根据中龄林和古核桃林林冠下灌草存有情况,在每个大样方随机选择11个中龄林、8个古核桃林2 m×2 m的小样方,进行灌木层、草本层植物调查。
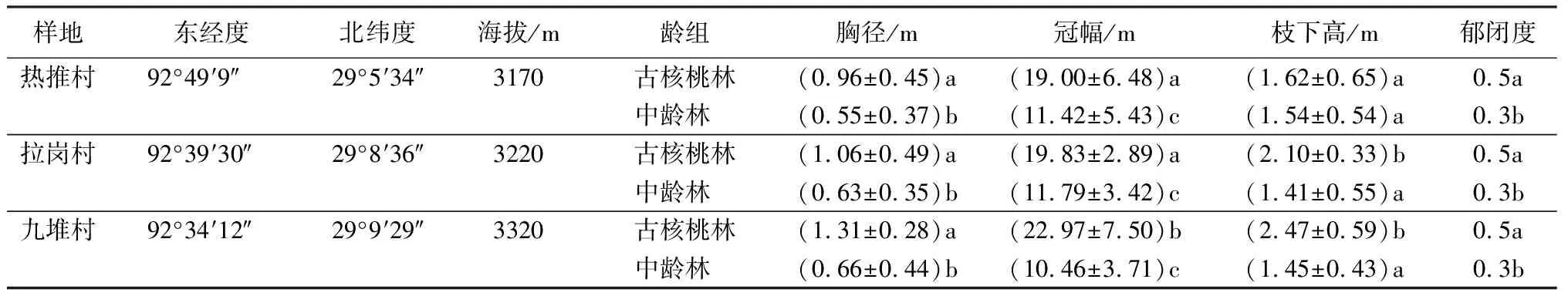
表1 不同样地核桃林林分特征
2.2 物种多样性计算
重要值计算[10]:重要值=(相对密度+相对频度+相对盖度)/3。
物种多样性指标[11]计算:
物种丰富度(S)=物种数;
Margalef丰富度指数Ma=(S-1)/lnN。
式中:Pi=Ni/N;Pi为抽样个体属于某一物种的概率;Ni为第i种物种个数;N为物种总数;S为样方内物种数。
2.3 土壤养分测定
2.4 土壤酶活性测定
土壤酶活性指标测定参照关松荫等[13]主编的《土壤酶及其研究法》。脲酶活性采用靛酚蓝比色法测定,以24 h后每克土壤中的NH3-N数表示其酶活性;β-葡聚糖酶活性采用硝基酚比色法测定,以每克土壤中产生的对硝基苯酚质量分数表示其酶活性;纤维素酶活性采用3,5-二硝基水杨酸比色法测定,以72 h后每克土壤中纤维素的消耗量表示其酶活性;酸性磷酸酶活性采用磷酸苯二钠比色法测定,以2 h后每克土壤中酚的质量分数表示其酶活性;过氧化氢酶活性采用高锰酸钾滴定法测定,以每克土壤消耗的标准高锰酸钾质量分数表示其酶活性;多酚氧化酶活性采用没食子素比色法测定,以每克土壤在2 h内紫色没食子素质量分数表示其酶活性。
2.5 数据处理
采用IBM SPSS 20.0和Excel 2016软件对数据进行处理分析;采用单因素方差分析对不同林龄西藏核桃林林下土壤理化性质和酶活性进行显著性分析(P=0.05);采用T检验对中龄林和古核桃林林下植被多样性数据进行处理;采用Pearson相关分析对土壤主要养分指标、酶活性、林下植物物种多样性指数、群落组成进行分析;利用Bray-Curtis相异性法对林下植被群落组成的差异进行非度量多维测度排序分析,在二维排序图上表示林下植被群落组成在不同林龄间的差异。
3 结果与分析
3.1 不同林龄核桃林林下科、属、种数量及重要值
研究区域内林下草本层共发现植物38种,属于35科49属(表2),其中,幼龄林占1种,隶属于1科1属;中龄林占26种,属于17科22属;古核桃林占28种,隶属于17科26属。林下灌木层共发现植物6种,属于3科6属(表3),其中,中龄林占4种,隶属于2科4属;古核桃林占5种,隶属于3科5属。

表2 林下草木层主要物种组成及重要值
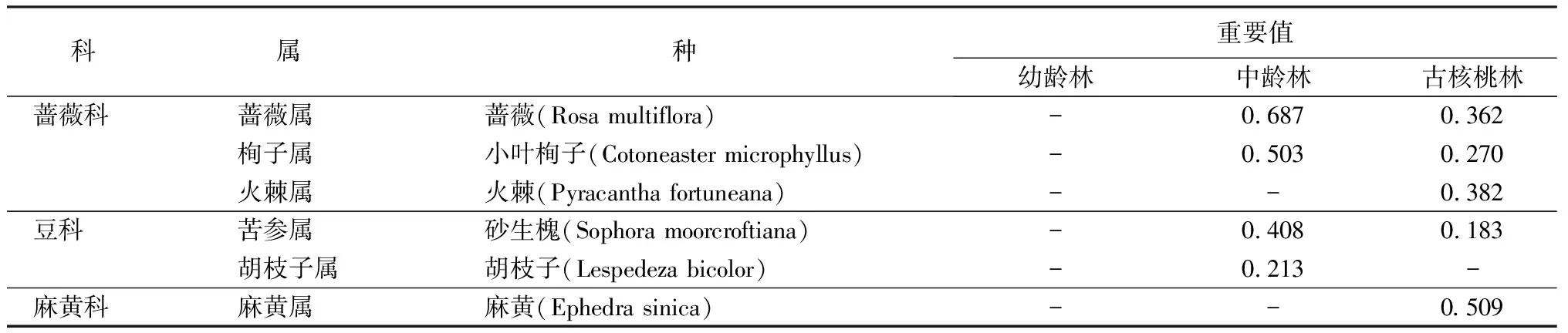
表3 灌木层主要物种组成及重要值
3.2 林龄对核桃林林下土壤养分及酶活性的影响
3.2.1 不同林龄核桃林林下土壤酶活性变化特征
林龄变化显著影响土壤酶活性(表4)。除过氧化物酶和β-葡聚糖酶外,其余酶活性均随核桃林龄的增加呈先增加后降低的趋势,中龄林土壤酶活性显著高于幼龄林及古核桃林(P<0.05)。古核桃林林下土壤脲酶活性、酸性磷酸酶活性、纤维素酶活性、碱性磷酸酶活性、多酚氧化酶活性、葡萄糖苷酶活性、氨基葡萄糖苷酶活性较中龄林分别显著降低了13%、21%、14%、34%、10%、39%、37%。β-葡聚糖酶活性随林龄的升高而升高,其中,古核桃林较中龄林、中龄林较幼龄林差异显著(P<0.05)。过氧化物酶活性随林龄升高呈先降低后升高的趋势,其中,古核桃林较中龄林、中龄林较幼龄林差异显著(P<0.05)。

表4 不同林龄核桃林林下土壤酶活性
3.2.2 不同林龄核桃林林下土壤养分变化特征
西藏高原不同林龄核桃林的林下土壤养分质量分数变化见表5。林龄显著影响核桃林林下0~20cm土层土壤有机质、速效磷、硝态氮质量分数(P<0.05),土壤有机质、速效磷、硝态氮质量分数均随林龄的增加呈先升高后降低的趋势。古核桃林的有机质、速效磷、硝态氮质量分数较中龄林分别显著的降低了30%、36%、29%。
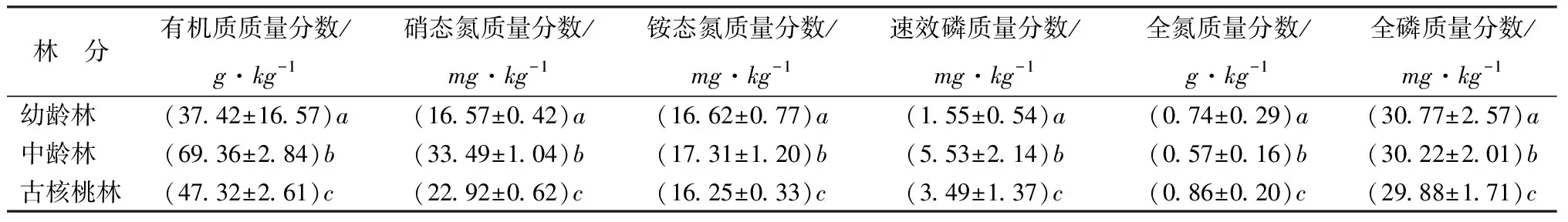
表5 不同林龄核桃林林下土壤养分质量分数
3.3 林龄对林下植物多样性指数及物种组成的影响
西藏高原核桃林林龄对林下植被的多样性及群落组成有显著影响(表6)。古核桃林林下植被Shannon-Wiener多样性指数、Margalef丰富度指数较中龄林显著降低(P<0.05),但Pielous均匀度指数、Simpson优势度指数与中龄林差异不显著。此外,林龄对西藏高原核桃林林下植物群落组成有显著影响(P<0.05)。非度量多维尺度分析得出,中龄林核桃林与古核桃林群落组成的布雷柯蒂斯距离较远(图1),进一步进行安东尼分析得出P=0.02(P<0.05),由此说明,林龄对西藏核桃林林下物种组成具有显著性影响。
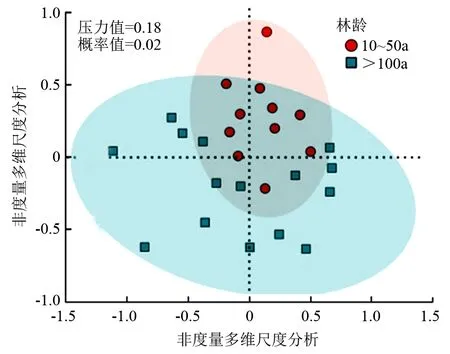
图1 中龄林及古核桃林林下物种组成

表6 中龄林及古核桃树林林下植被多样性指数
3.4 中龄林及古核桃林林下土壤养分质量分数、酶活性与植物群落的相关性
中龄林相关性分析表明,Margalef丰富度指数,与碱性磷酸酶活性、脲酶活性、氨基葡萄糖苷酶活性呈显著负相关(P<0.05),与过氧化物酶活性呈显著正相关(P<0.05)。林下植被群落组成与纤维素酶活性、多酚氧化酶活性呈显著正相关(P<0.05,表7)。

表7 中龄林林下土壤养分质量分数、酶活性与植被多样性指数的相关系数
古核桃林相关性分析表明,Pielous均匀度指数与氨基葡萄糖苷酶活性、β葡聚糖酶活性呈显著正相关(P<0.05);Shannon-Wiener多样性指数、Simpson优势度指数、Margalef丰富度指数与多酚氧化酶活性、氨基葡萄糖苷酶活性呈显著正相关(P<0.05);Shannon-Wiener多样性指数、Pielous均匀度指数与纤维素酶活性呈显著负相关(P<0.05);林下植被群落组成与土壤养分、酶活性均无显著相关性(表8)。
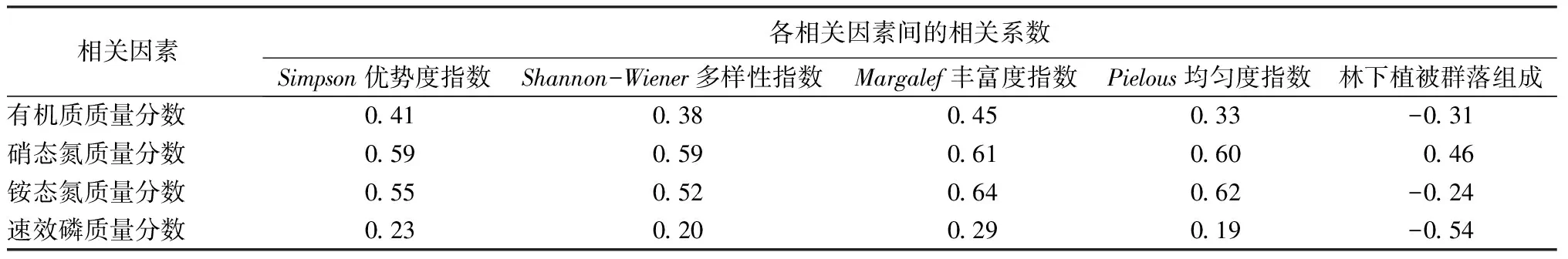
表8 古核桃树林林下土壤养分质量分数及酶活性与植被多样性指数的相关系数
4 结论与讨论
研究表明,随着林龄的增加,森林生态系统的组成结构、内部环境、土壤性质都会随之发生改变[14],进而影响土壤养分质量分数。本研究发现,西藏核桃林随着林龄的增加,土壤有机质质量分数、速效磷质量分数、硝态氮质量分数均呈先升高后降低的趋势,且有显著差异(P<0.05)。出现该现象的原因是随着核桃林龄增长,至旺盛时期时,其盖度增加,导致凋落物随之增加。同时,此阶段核桃根系分泌物对土壤微生物影响较小[15],但此时,对土壤微生物影响较大的有机质质量分数增加,提高了微生物活性,加快了凋落物分解速率,进而增加了土壤氮、磷质量分数,使核桃生长至中龄期时呈现养分升高的趋势。但是对于古核桃林,由于其长期对土壤养分的消耗,加之当地土壤不施加任何肥料,使得土壤养分质量分数逐渐降低[16]。此外,本研究发现,由于古核桃树冠层较大、林内地面光照较差、西藏高原温度较低,使得凋落物分解速率减慢,进而导致土壤养分质量分数降低。核桃是典型含有化感物质的树种,其凋落物和根系分泌物的化感作用对土壤微生物具有显著影响[17],因此导致微生物活性下降,进一步影响土壤养分质量分数[18]。土壤获取磷元素的主要途径来自于岩石风化和淋洗,方式比较单一[19],且土壤中磷元素主要以无机磷的形态存在,而无机磷中能被植物直接利用吸收的磷占比很少,所以土壤全磷质量分数丰富,但植物可吸收的有效磷亏缺[20]。因此,土壤总磷质量分数随林龄的增加其变化趋势并不明显。林龄对铵态氮无显著影响,这是由于硝态氮可以被还原成铵态氮[21],而铵态氮较硝态氮而言,更利于核桃地下根系生长[22]。因此,核桃根际土壤周围的铵态氮质量分数保持动态平衡,不随林龄增加发生显著变化。
土壤酶是土壤生态系统的重要组成部分,参与土壤有机物的合成、分解、转化等过程,在森林生态系统土壤养分循环过程中发挥重要作用[23]。在西藏高原不同林龄核桃林中,参与氮、磷转化的酶活性均随着林龄增加呈先升高后降低的趋势,其酶活性在核桃中龄林中达到峰值。造成这种现象一方面是由于随着林龄升高,核桃林分郁闭度逐渐增加,其凋落物的数量和质量也随之升高,凋落物分解改变了土壤微生物数量[24],从而提高了核桃中龄林林下土壤酶活性;另一方面,核桃中龄林生长旺盛,地下根系需要吸收更多的氮、磷以供植物生长,促使氮、磷转化酶的活性增强[25]。此后,随着林龄不断增长,古核桃树周围土壤肥力下降[26],郁闭度继续增加,光照降低,林内温度及湿度发生变化,导致微生物数量及活性降低,枯落物分解转化强度减弱,土壤酶活性随之降低[27]。β-葡聚糖酶是土壤碳循环过程中的关键酶,其活性随林龄增长而升高,与全氮的变化趋势相同。出现该现象的原因是随着林龄升高,氮质量分数增加,微生物通过同化氮素促进自身生长,激发了土壤有关微生物分泌酶,因此,β-葡聚糖酶活性增加[28]。
林下植物物种多样性是反映林下植物群落结构的组成、功能、演替形态、稳定性的重要指标[29]。本研究中,西藏高原不同林龄的核桃林林下植物群落Shannon-Wiener多样性指数、Margalef丰富度指数差异显著(P<0.05),Pielous均匀度指数、Simpson优势度指数差异不显著。出现该现象是因为随着林龄的升高,林分演替到达中后期,林分趋于成熟,群落稳定导致林下物种更新减缓[30],影响了物种优势度,使物种数降低且逐渐趋于均匀化[31],最终导致多样性减少。非度量多维尺度分析显示,中龄林与古核桃林林下植物群落组成差异显著(P<0.05),随着林龄增加,林下植物的物种相似度随之变小。出现这一现象一方面是由于在林龄增加的过程中,林分郁闭度逐渐趋于0.5。大量研究表明,郁闭度在0.5左右时林下植物群落种类较为稳定[32],且西藏地处青藏高原,其强太阳辐射及光质、低温、低气压等因素使植物长期演化,形成了独特的适应机制[33]。另一方面,物种多样性发生变化是环境因子、人为干扰、物种竞争力综合作用决定的[34]。西藏高原人工林在初期造林时,当地人多使用实生苗,林分郁闭度不高,出现了大量随机入侵物种,而随着林龄增长,适应性较强的物种逐渐转为优势种[35],使物种组成发生变化并趋于稳定[36]。因此,在人工林的造林过程中应注重合理的伴生植物配置,营造良好的养分循环微气候。
核桃中龄林的Margalef丰富度指数与脲酶活性、碱性磷酸酶活性、过氧化物酶活性、氨基葡萄糖苷酶活性存在显著或极显著相关关系,其中,碱性磷酸酶活性与Margalef丰富度指数呈极显著负相关(P<0.01)。研究表明,土壤碱性磷酸酶活性与土壤微生物的活动显著相关[37],这是由于中龄林较高的林下植物丰富度促进了土壤微生物活动,而微生物活动频繁使根际土壤中可利用的有效磷达到饱和状态,降低了碱性磷酸酶活性,导致土壤中碳氮比发生改变[38],从而改变了其他碳氮循环相关酶的分泌。
古核桃林多项参与碳氮循环的酶活性(除纤维素酶外)均与林下植物多样性指数呈显著正相关,这是因为土壤纤维素酶参与土壤养分循环的方式主要是分解纤维素来为微生物提供能源物质[39]。古核桃林林下土壤肥力较中龄林弱,纤维素酶活性升高可为微生物的活动提供更多能源,以维持正常土壤养分循环。总体来看,西藏古核桃林林下植物多样性与根际土壤各项养分指标的相关性高于中龄林。
不同林龄核桃林林下土壤养分、酶活性会随林龄升高发生显著变化[40]。本研究表明,不同林龄核桃林林下植物群落物种组成差异显著,古核桃林林下植物多样性与土壤养分酶活性相关性更高。因此,100a林龄以上的古核桃林要注重林下物种组成和群落结构的配置,可以考虑人工改善选择出最适合的核桃林林下伴生植物,营造多样性高、稳定性强的群落环境,发挥西藏高原核桃林最优的经济功能和生态功能。
