干旱胁迫下大苞萱草bHLH转录因子家族鉴定与分析
2023-07-05陈丽飞刘云怡慧李嘉峻周蕴薇
陈丽飞,刘云怡慧,李嘉峻,白 云,孟 缘,周蕴薇
(1.吉林农业大学林学与草学学院,长春 130118;2.吉林农业大学园艺学院,长春 130118)
【研究意义】大苞萱草(Hemerocallismiddendorffii)隶属阿福花科(Asphodelaceae)萱草属(Hemerocallis)多年生宿根草本植物,是我国北方优良的露地宿根花卉[1-2]。因其花期较早,颜色以橘黄色为主,有“早金花”的美称。大苞萱草花色艳丽且具有较强的抗旱能力,可作为优良地被绿化及庭院观赏植物。干旱作为非生物胁迫主要因素之一,持续的干旱不仅会导致植物的生长和发育停滞甚至死亡,还能使植物进行抗旱基因的表达,通过分子机制的调控表达出植株的抗旱性[3-5]。bHLH(Basic helix-loop-helix,碱性螺旋-环-螺旋)转录因子是广泛存在于真核生物中的一类重要转录因子,家族成员数量仅次于MYB家族,在生长发育、次生代谢和环境胁迫中均具有重要的调控作用。通过研究植物耐旱机制,筛选参与抵御干旱胁迫的相关基因,对推动提高植物抗旱性育种的发展起到重要作用。【前人研究进展】1989年,Eurre等[6]在小鼠肌肉发育的研究中首次发现bHLH转录因子E12和E47,随后在动物、植物及菌类在内的所有真核生物中被鉴定及分离。bHLH转录因子的保守结构域由约60个保守氨基酸残基和2个保守基序组成,分为基本(Basic)区和螺旋-环-螺旋(HLH)区[7-8],除保守结构域外的其他序列具有较大差异。基本区具有特定DNA序列结合活性特点,螺旋区域在蛋白质相互作用中起作用,这种同时具备DNA和蛋白质相互作用的特点,且大多数以形成二聚体的方式发挥其生物学功能,使得bHLH转录因子在植物生长调节等方面非常重要[9-11]。研究表明,拟南芥中大部分bHLH蛋白功能特征已被鉴定出来,包括参与植物形态建成、种子萌发、激素信号及应激反应[12-16]。不同植物中的bHLH基因家族成员数量各有不同,在拟南芥(Arabidopsisthaliana)、水稻(Oryzasativa)、结缕草(Zoysiajaponica)、红花(Carthamustinctorius)中分别发现bHLH家族成员167、178、141、41个[17-20],随着bHLH基因家族成员功能研究的深入,越来越多的bHLH转录因子被证明与植物抵抗干旱胁迫有关,如:OsbHLH148、OsbHLH120、GmbHLH125、EcbHLH57等转录因子。OsbHLH148能够通过参与茉莉酸信号途径对水稻的干旱耐受性起调节作用[21]。OsbHLH120可通过参与ABA信号通路,使植株在干旱胁迫下仍能保持一定的含水量,提高水稻的抗旱能力[22-23]。Zhai等[24]对小麦的研究中发现,过表达TabHLH39可增强小麦的抗寒、抗旱和抗盐能力。Dong等[25]对PebHLH35功能研究中发现,该基因可增强胡杨的抗旱性。【本研究切入点】由于目前对大苞萱草分子生物学方面的研究起步较晚,虽然bHLH转录因子在植物中的研究较为成熟,但是大苞萱草bHLH家族基因的研究鲜见报道。【拟解决的关键问题】本研究基于转录组数据,利用生物信息学方法对大苞萱草bHLH基因家族成员进行鉴定并构建系统发育进化树,通过对蛋白理化性质、保守结构域、表达模式的分析,为揭示bHLH转录因子在大苞萱草响应干旱胁迫的过程中发挥一定作用提供理论依据。
1 材料与方法
1.1 试验材料与胁迫处理
供试材料为生长健壮的大苞萱草一年生幼苗,种植于吉林农业大学植物资源圃。选取生长健壮、长势一致的大苞萱草,定植于相同基质(草炭∶珍珠岩=3∶1)的塑料花盆中进行为期35 d的缓苗工作。针对试验材料进行胁迫处理:采用PEG-6000模拟干旱法,分为对照组(CK)和干旱胁迫组(20%PEG-6000),每个试验组设置3个重复,每个重复4株,于同一时间浇入相同容量的超纯水和20% PEG-6000溶液。干旱胁迫24 h后,采集试材新鲜的根尖(R)及嫩叶(L)并用锡箔纸包装于液氮中速冻,随后置于-80 ℃冰箱中保存,将样品送至北京诺和致源科技股份有限公司进行转录组测序。
1.2 大苞萱草bHLH基因家族成员的鉴定及理化性质分析
在TAIR网站(https://www.arabidopsis.org/)下载拟南芥bHLH转录因子的基因序列及蛋白序列,利用本地BLAST和在线软件Smart(http://smart.embl-heidelberg.de/)进行成员筛选,去除不含bHLH结构域和重复的序列,得到具有bHLH保守域的大苞萱草bHLH转录因子。利用在线软件ExPASy ProtParam tool(https://web.expasy.org/protparam/)获得大苞萱草bHLH蛋白的理化性质,如氨基酸序列长度、相对分子质量、蛋白质等电点等。利用网站CELLO V2.5(http://cello.life.nctu.edu.tw/)对大苞萱草bHLH基因家族成员亚细胞定位进行预测。
1.3 大苞萱草bHLH蛋白质二级结构预测
利用在线软件SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa-sopma.html)对大苞萱草bHLH蛋白质二级结构进行预测。
1.4 大苞萱草bHLH基因家族保守基序分析及多重序列比对
通过在线软件MEME(https://meme-suite.org/meme/tools/meme)对大苞萱草bHLH蛋白保守基序进行搜索并使用TBtools软件将大苞萱草bHLH转录因子蛋白保守基序可视化。利用DNAMAN 6.0软件对大苞萱草bHLH家族基因编码的氨基酸序列进行多序列比对。
1.5 大苞萱草bHLH基因家族系统进化分析
以拟南芥作为参照,使用MEGA 7.0软件对大苞萱草bHLH转录因子构建系统发育进化树,采用邻接(Neighbor-Joining method)法,Bootstrap值设置为1000,其他参数选择默认值。使用在线软件iTOL(https://itol.embl.de/)对系统发育树进行美化处理。
1.6 大苞萱草bHLH基因家族在叶和根中的表达分析
利用北京诺禾致源科技股份有限公司提供的售后平台工具NovoMagic(https://magic.novogene.com/customer/main#/login)分别制作叶和根中55个大苞萱草bHLH基因家族成员在处理组与对照组的表达模式热聚图。
2 结果与分析
2.1 大苞萱草bHLH基因家族成员的鉴定及理化性质分析
基于转录组测序得到232个大苞萱草bHLH转录因子。利用在线软件Smart和NCBI去除重复及不含有bHLH结构的大苞萱草bHLH转录因子,最终筛选得到55个大苞萱草bHLH转录因子并命名为HmbHLH1~HmbHLH55。
55个大苞萱草bHLH基因编码氨基酸的理化性质分析表明(表1),氨基酸序列长度最长和最短的基因分别为HmbHLH7和HmbHLH9,其肽链长度分别为824和100 aa,分子量分别为90.68和11.46 kD,平均氨基酸数为321.38 aa。平均亲水指数均为负值且具有较强的亲水性,是亲水性蛋白。等电点的范围为4.85~10.45,70.9%的HmbHLH家族蛋白的等电点小于7,属于酸性蛋白。55个大苞萱草bHLH基因家族成员的亚细胞定位预测(表2)表明,共有33个HmbHLH转录因子分布在细胞核内,21个HmbHLH转录因子分布在叶绿体和线粒体中,仅有HmbHLH25位于细胞质中。
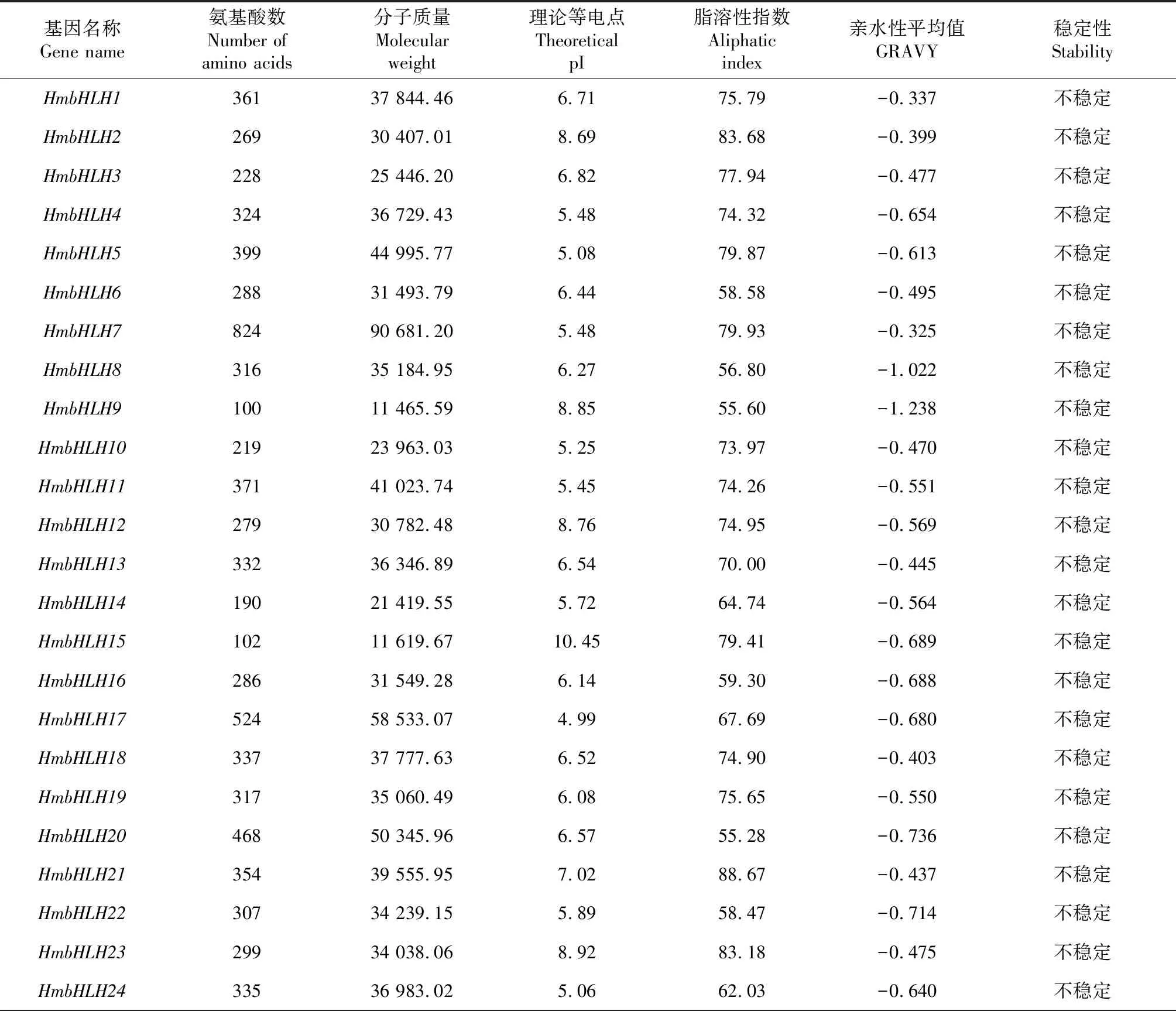
表1 HmbHLH转录因子的理化性质
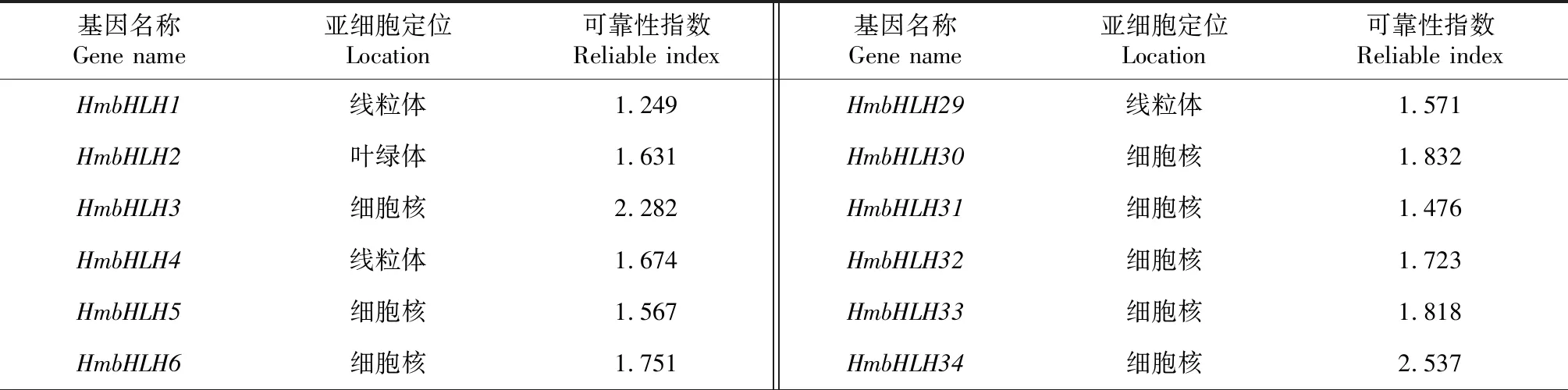
表2 HmbHLH转录因子亚细胞定位
2.2 大苞萱草bHLH蛋白质的二级结构预测
如表3所示,55个HmbHLH转录因子蛋白二级结构均含有α-螺旋、β-折叠、延伸链以及无规卷曲,且HmbHLH蛋白均呈现出无规卷曲数>α-螺旋数≥延伸链数>β-折叠数的特点。

表3 HmbHLH转录因子蛋白二级结构
2.3 大苞萱草bHLH基因家族聚类分析
将鉴定得到的55个大苞萱草bHLH基因家族成员与161个拟南芥bHLH基因家族成员构建系统发育进化树(图1)。根据预测的HmbHLH蛋白保守基序包含种类的相似性,对其进行分类。综合进化树分类情况和AtbHLH转录因子的分类系统,将HmbHLH基因家族成员分为11个亚家族。第6亚族成员数量最多,含有32个;第4亚族成员最少,含有5个。
2.4 大苞萱草bHLH家族蛋白保守基序(motif)分析及多重序列比对
55个大苞萱草bHLH蛋白序列的保守基序预测结果(图2)表明,49个HmbHLH蛋白含有motif1,51个HmbHLH蛋白含有motif2,推测motif1和motif2是特征保守基序,共同构成bHLH结构域(图3)。属于同一亚组的bHLH转录因子有着相似的motif类型和数量,同时也存在差异,亲缘关系较近的HmbHLH18和HmbHLH53包含motif种类和数量均相同,但近缘的HmbHLH55较以上2个蛋白少了motif14。在大苞萱草bHLH家族基因编码蛋白中,HmbHLH9、HmbHLH16、HmbHLH35、HmbHLH44的motif类型最简单且仅含有1种,HmbHLH18、HmbHLH53、HmbHLH55含有motif类型最多,为10种。
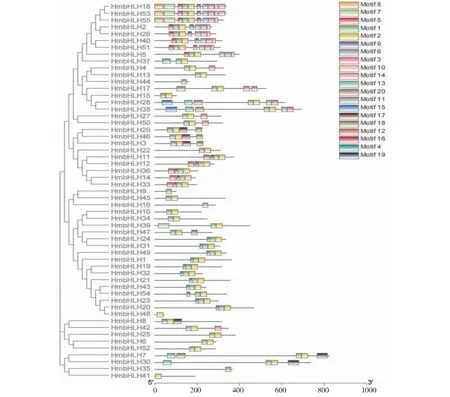
图2 HmbHLH转录因子家族保守基序

不同位点上的氨基酸字母的高度代表保守型的高低。
多重序列比对结果(图4)表明,55个HmbHLH蛋白均具有典型的basic-helix-loop-helix(bHLH)保守结构域,除loop区,其他区域均具有相对保守的氨基酸位点。如图5所示,在碱性区域(basic)中的氨基酸数量较少,第8和9位上具有高度保守的精氨酸。2个螺旋区中的第19和47位上具有高度保守的亮氨酸(L)。
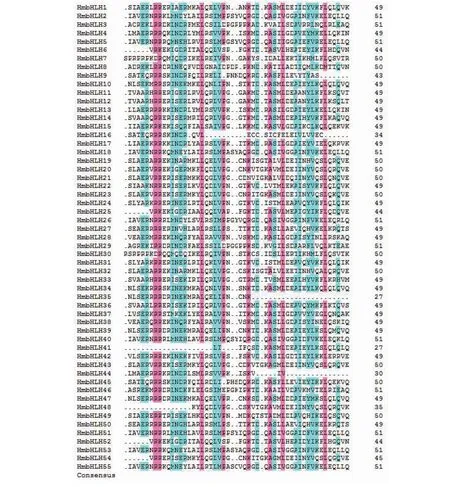
图4 HmbHLH结构域的氨基酸比对
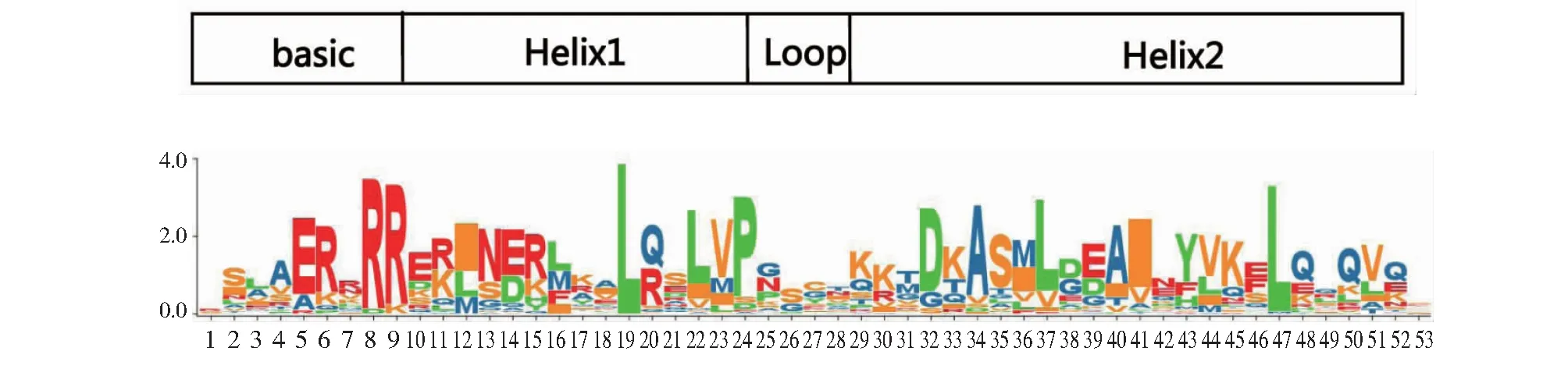
图5 HmbHLH转录因子家族的保守结构域
2.5 大苞萱草bHLH基因家族在叶和根中的表达分析
基于转录组数据的分析,发现55个HmbHLH转录因子在大苞萱草叶和根中具有不同的表达模式(图6)。与对照组相比,处理组的叶中有上调基因26个,下调基因26个,其中HmbHLH24和HmbHLH26在叶中无表达,HmbHLH45处理前后的表达量不变,HmbHLH50、HmbHLH20、HmbHLH21、HmbHLH43明显上调,HmbHLH14、HmbHLH55明显下调。处理组的根中有上调基因32个,下调基因22个,HmbHLH14在根中无表达,HmbHLH16、HmbHLH29明显上调,HmbHLH7、HmbHLH4、HmbHLH10明显下调。在受到干旱胁迫后,大苞萱草叶中HmbHLH基因家族成员的表达量总体呈上升趋势,根中HmbHLH基因家族成员的表达量总体呈下降趋势。从55个HmbHLH基因家族成员中共筛选得到6个差异基因(图7),叶中有4个差异基因,2个上调基因和2个下调基因;根中有2个差异基因,均为上调基因。
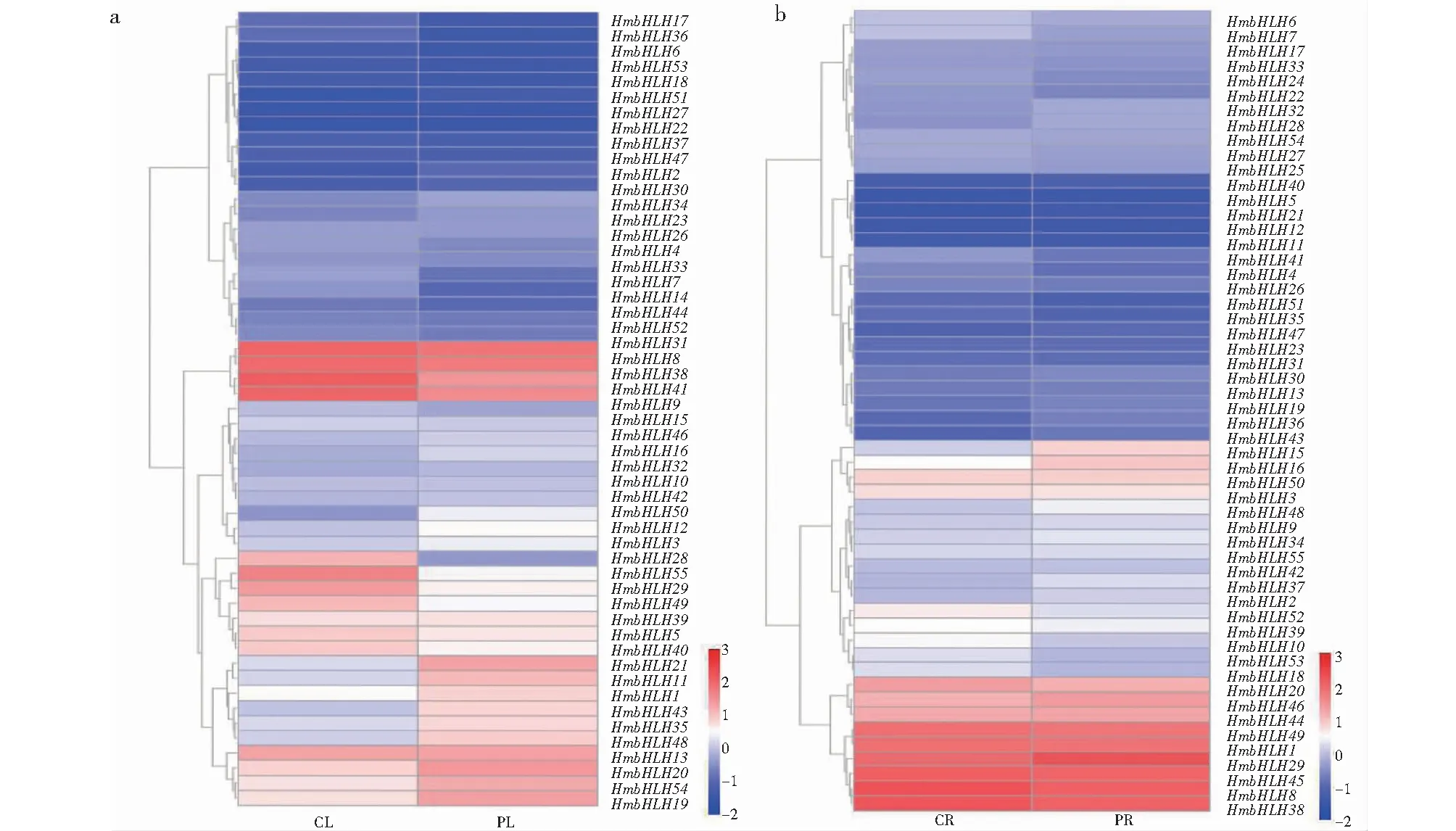
CL/CR为对照组,PL/PR为处理组,下同。

图7 不同处理下叶(L)和根(R)中HmbHLH差异基因的韦恩图
3 讨 论
植物bHLH转录因子的功能主要包括参与调节各种信号转导及合成,如在干旱胁迫中参与调控植物的信号转导来发挥重要作用。研究表明,干旱胁迫下水稻OsbHLH148可通过参与调节茉莉酸信号途径,来调节植株对干旱的耐受程度[21]。过表达PebHLH35基因会显著降低拟南芥叶片的气孔密度和开度,降低自身的蒸腾速率,从而维持干旱胁迫下植株的湿度,提高植物的抗旱能力[26]。bHLH转录因子可以通过特定的氨基酸残基与靶基因结合来调节相关基因的表达,调控植物抗旱能力[27]。
本研究共鉴定出55个大苞萱草bHLH基因家族成员,通过对HmbHLH转录因子保守结构域分析发现,有22个氨基酸残基保守位点且保守性均超过50%,其中Leu-19氨基酸残基完全保守。Atchley等[28-29]认为bHLH碱性区能够辨别并结合靶DNA,螺旋区则在形成同源或异源二聚体方面发挥作用。由此,HmbHLH碱性区的特殊位点Glu-5、Arg-8和Arg-9可能在DNA结合中发挥重要作用,螺旋区的Leu-19和Leu-47高度保守,可能在形成二聚体中发挥作用。大苞萱草bHLH基因家族划分为11个亚族,相同亚族的大苞萱草和拟南芥成员之间存在相似的基因结构及生物学功能。拟南芥中第6亚族的转录因子,如AtbHLH4、AtbHLH5、AtbHLH6、AtbHLH17等可通过参与ABA合成、茉莉酸等激素信号途径来提高植株抗逆性[30-31]。因此,同亚族的HmbHLH17、HmbHLH28、HmbHLH38等可能与大苞萱草的抗旱性相关。拟南芥中第10亚族的AtbHLH15和AtbHLH24能够参与植物形态建成及种子萌发,表明HmbHLH47和HmbHLH39可能在大苞萱草种子的萌发过程中发挥一定作用。
大苞萱草bHLH蛋白保守结构域分析发现,同一亚家族大多数成员具有的保守基序种类和数量相似,表明每个亚家族编码蛋白的功能相对稳定。在大苞萱草bHLH蛋白中发现了20种保守基序,几乎所有蛋白均包含motif1和motif2且基序motif1和motif2含有保守氨基酸精氨酸(R)和亮氨酸(L),推测motif1和motif2是大苞萱草bHLH转录因子的保守基序,共同构成了bHLH结构域,在HmbHLH基因行使功能时起重要作用。在植物的进化过程中亚组高度保守,相同的亚组蛋白保守基序相似。特定的亚组存在着特殊的基序,表明这些保守基序对亚族特异的保守功能起着关键作用。大苞萱草bHLH蛋白保守基序的分析为bHLH基因的分类与进化提供了重要的参考依据。
在干旱胁迫下HmbHLH转录因子的表达模式中,叶和根的基因表达均发生差异性变化。叶中有4个差异基因,分别为HmbHLH17、HmbHLH21、HmbHLH28和HmbHLH50,根中有2个差异基因,分别为HmbHLH16和HmbHLH29。经干旱胁迫前后的对比发现,叶中HmbHLH21和HmbHLH50的表达量均升高且HmbHLH50表达量变化显著,HmbHLH17和HmbHLH28的表达量均下降。根中HmbHLH16和HmbHLH29的基因表达量均升高。Aftab等[32]研究发现,干旱条件下过表达的AtbHLH106能有效减少气孔开度来减少植株的水分蒸腾,以此来提高植株的抗旱能力。AtbHLH106转录因子可以结合G-box和E-box顺式作用元件,抑制下游逆境基因的表达进而增加ABA的积累,提高拟南芥的抗旱能力。通过本研究构建的系统发育进化树发现,HmbHLH50与AtbHLH106具有较近的亲缘关系,间接表明HmbHLH50可能通过促进其表达来参与大苞萱草响应干旱胁迫的过程,但还需进一步验证其生物学功能。
4 结 论
bHLH转录因子在植物的逆境调节中发挥着重要作用。本研究基于转录组测序数据,共鉴定出55个大苞萱草bHLH基因家族成员并分为11个亚家族,HmbHLH蛋白的保守基序和理化性质具有特异性和多样性。通过对不同处理下叶和根中bHLH基因的表达分析,表明大苞萱草bHLH转录因子家族基因可能与大苞萱草的抗旱性相关,为深入研究bHLH转录因子在大苞萱草响应干旱胁迫过程中的生物学功能及分子机制提供理论基础。
