克隆整合对入侵杂草空心莲子草和本土莲子草生长及光合性能的影响
2023-07-01郭文锋李晓琼
宋 誉,郭文锋,李晓琼
(1 广西大学 林学院/广西森林生态与保育重点实验室,广西 南宁 530004;2 广西农业科学院/广西作物遗传改良生物技术重点开放实验室,广西 南宁 530007)
空心莲子草是一种原产于南美洲的克隆入侵植物[14],在世界范围内造成严重的经济和环境问题[15],被列为世界上最严重的入侵杂草之一[16]。空心莲子草可以忍受各种各样的环境条件,可在水生、半水生和湿生环境中生长[1]。在中国,空心莲子草表现出极低的遗传多样性,不能产生有活性的种子[17],而是通过茎和根的克隆生长产生后代分株[18]。当空心莲子草处于资源供应较高的同质环境中时,克隆整合能提高空心莲子草的生长性能[19];而有研究表明克隆整合并不能促进同质环境中空心莲子草整个克隆片段的生长,但可改变相连分株间的资源分配[14]。在我国,空心莲子草常常与本地同属种莲子草A.sessilis同域分布,但空心莲子草的竞争能力要显著高于莲子草[20]。然而,克隆整合在空心莲子草挤占莲子草生存空间中的作用机制尚不明确。空心莲子草能比莲子草更好地整合小尺度土壤异质性,从而在异质性大的地区(如氮沉降、施肥和干扰)入侵成功[21]。在竞争环境下,空心莲子草具有明显的劳动分工现象,并且克隆整合能够显著增强空心莲子草基端分株的生长及竞争能力,从而有利于其入侵[9]。本文通过比较克隆整合下空心莲子草和莲子草生长性能、根系分配、生物量分配及光合性能的差异,探究克隆整合与空心莲子草强竞争力间的关系。
1 材料与方法
1.1 试验材料
试验于广西大学林学院的苗圃内进行。参照陈燕丽等[22]的方法扦插育苗。选取数株长势良好的空心莲子草和莲子草,剪成若干长约4 cm、带有1 个茎节的茎段。将茎段插入装有混合灭菌土壤[V(壤土)∶V(草炭土)=1∶1]的苗盘中。30 d 后,选取长约25 cm、含有4 个茎节和1 个茎尖的空心莲子草和莲子草匍匐茎,将带有茎尖的2 个茎节(先端分株) 种植在1 个花盆(直径25 cm、高16 cm)中,将带有2 个较老茎节的基端(基端分株)种植在相邻花盆中。用0.15 mm 孔径的防虫网单独罩住所有花盆,以隔绝其他昆虫。
1.2 克隆整合处理
试验包括2 种植物(空心莲子草、莲子草),2 种克隆整合处理(连接、剪断)(图1);共4 个处理,每个处理重复5 次。克隆整合处理是将先端分株和基端分株保持连接,无克隆整合处理组则将先端分株和基端分株剪断,然后让植物生长60 d。试验期间,苗圃白天温度28~33 ℃,夜间温度24~28 ℃,相对湿度50%~75%。
图1 试验设计Fig.1 Experiment design
1.3 光合指标的测定
使用美国Li-6400 便携式光合测定仪,选取被测植株顶端完全展开且正常生长的第3~5 片叶进行光合指标测定,测定时间为晴朗天气的08:30—11:30,设定叶面温度为(26±3) ℃,测定指标包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci),并计算水分利用效率(WUE):
测定时,使用CO2气瓶控制CO2体积分数为0.04%,同时设定光合有效辐射(PAR)为1 500 μmol·m-2·s-1。每个植株测3 个叶片,取平均值。
同样条件测定植物的光响应曲线,测定时,使用CO2气瓶控制CO2体积分数为0.04%,同时设定光合有效辐射PAR 为0、50、100、200、400、600、800、1 000、1 200、1 500 μmol·m-2·s-1。每个植株测3 个叶片,取平均值。
光响应曲线的拟合采用非直角双曲线模型[23],公式为:
式中,α指初始量子效率;Pmax指最大净光合速率;θ指光响应曲线的凸度,取值范围是0≤θ≤1;Rd指暗呼吸速率。
本文旨在揭示数值格式的适度耗散可由色散-耗散条件确定. 作为演示验证, 将色散-耗散条件用于减弱以往发展的加权基本无振荡格式WENO-CU6-M2[15]所造成的伪波, 通过数值算例给出该格式的两种直接改进并加以对比确认.
在PAR<200 μmol·m-2·s-1时,对光响应曲线PAR-Pn进行直线回归,计算各处理下空心莲子草和莲子草的光补偿点(LCP)和光饱和点(LSP)。
1.4 植物的收获与测定
收获前,分别统计植物先端和基端的叶片数和分株数,用卷尺和游标卡尺分别测定先端和基端的茎长和地径;然后分别收获先端和基端的地上和地下部分,收集地下部分时记录粗根数(直径≥2 mm)和细根数(直径<2 mm),并计算整个克隆片段(先端+基端)的指标。将收获的植株置于60 ℃的烘箱中烘干至恒质量;称量先端和基端分株的地上生物量和地下生物量,计算总生物量(地上生物量与地下生物量之和)和根冠比(地下生物量/地上生物量)。
1.5 数据统计与分析
采用双因素方差分析(Two-Way ANOVA)分别研究克隆整合处理、植物种类及两者交互作用对空心莲子草和莲子草先端分株、基端分株和整个克隆片段(先端+基端)的生长特性(叶片数、茎长、地径、分株数)、根系分配(粗根数、细根数、总根数)、生物量分配(地上生物量、地下生物量、总生物量、根冠比)、光合指标(净光合速率、气孔导度、胞间CO2浓度、蒸腾速率、水分利用效率)和光响应指标(最大净光合效率、暗呼吸速率、光饱和点、表观量子效率、光补偿点)的影响。植物种类和克隆整合处理作为固定因素,用Duncan’s 法进行多重比较。所有数据采用SPSS 22.0 进行统计分析,并用Sigmaplot 14.0 软件绘图。
2 结果与分析
2.1 克隆整合处理下2 种植物先端分株和基端分株地上部分生长及光合差异
植物种类和克隆整合处理对2 种植物先端分株、基端分株的叶片数及先端分株的分株数产生显著的交互影响(表1)。无克隆整合时,莲子草先端分株的叶片数以及基端分株的地径显著大于空心莲子草,然而克隆整合处理下,2 种植物先端分株的叶片数及基端分株的地径差异不显著;克隆整合显著增加了莲子草先端分株的分株数,从而使莲子草先端分株的分株数显著大于空心莲子草;克隆整合显著促进了空心莲子草先端分株的茎长(图2a~图2d)。植物种类及克隆整合处理并未对地径和茎长产生显著的交互影响(表1)。

表1 植物种类和克隆整合处理对空心莲子草和莲子草先端分株、基端分株及整个克隆片段生长及光合特性影响的双因素方差分析结果1)Table 1 Two-way ANOVA results of effects of plant species and clonal integration on growth and photosynthesis of apical ramets,basal ramets,and whole clone fragments of Alternanthera philoxeroides and A. sessilis
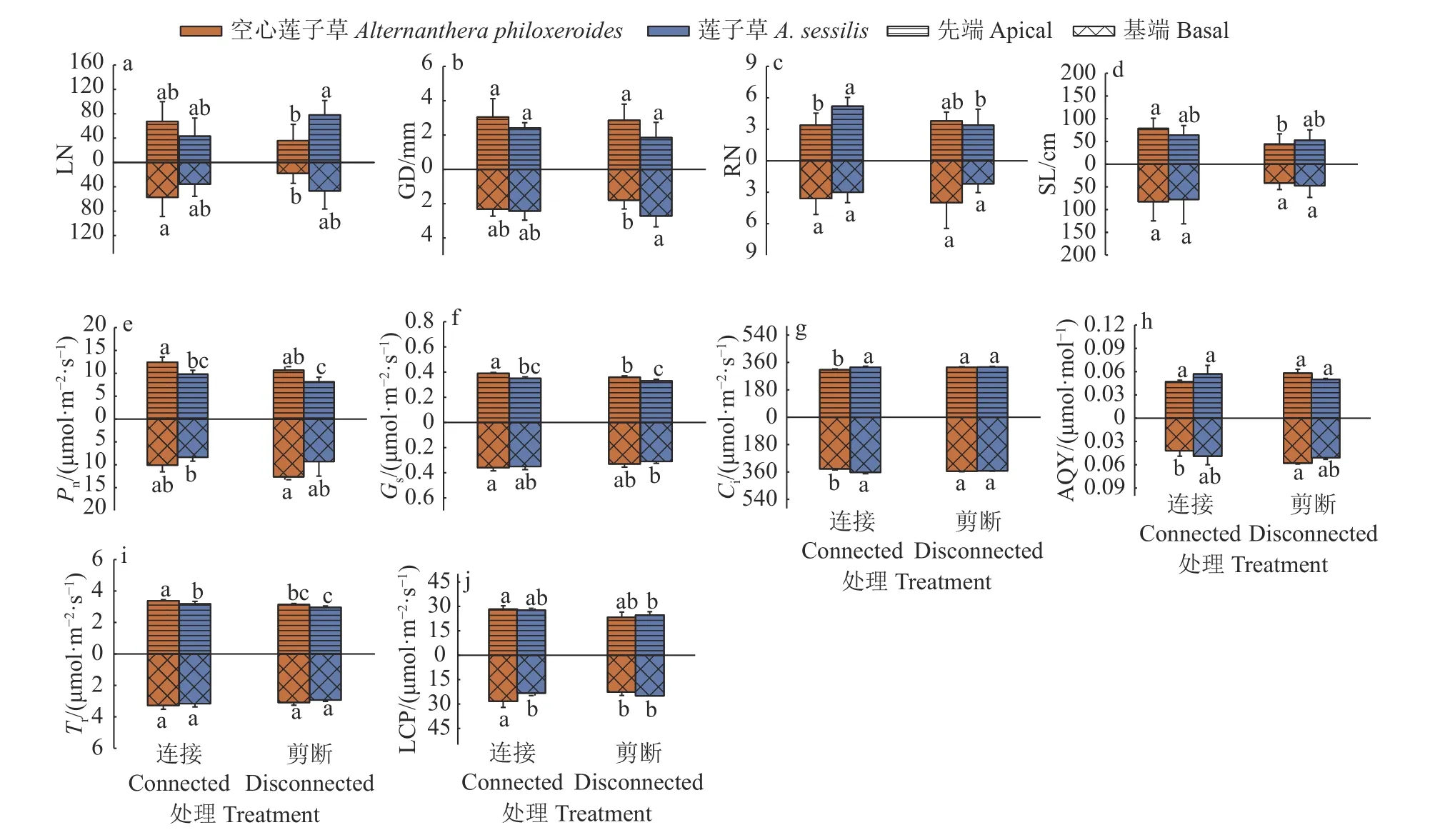
图2 克隆整合处理对2 种植物先端分株和基端分株地上部分生长及光合性能的影响Fig.2 Effects of clonal integration on the aboveground growth and photosynthesis of apical and basal ramets of two plants
植物种类和克隆整合处理对2 种植物先端分株、基端分株的胞间CO2浓度,先端分株的表观量子效率及基端分株的光补偿点产生显著的交互影响(表1)。克隆整合显著增加了空心莲子草先端分株的气孔导度,且无论有无克隆整合,空心莲子草先端分株的净光合速率和气孔导度均显著大于莲子草(图2e、2f)。无克隆整合时,2 种植物先端分株、基端分株的胞间CO2浓度及基端分株的表观量子效率无显著差异,而克隆整合显著降低了空心莲子草先端分株、基端分株的胞间CO2浓度及基端分株的表观量子效率,使其显著低于莲子草(图2g、2h)。无克隆整合下,2 种植物先端分株的蒸腾速率无显著差异,克隆整合虽显著增加了2 种植物先端分株的蒸腾速率,但空心莲子草的蒸腾速率显著大于莲子草(图2i)。克隆整合显著提高了空心莲子草基端分株的光补偿点,使其显著大于莲子草(图2j)。
2.2 克隆整合处理下2 种植物先端分株和基端分株根系及生物量分配情况
植物种类和克隆整合处理对2 种植物先端分株、基端分株的粗根数产生显著的交互影响,但对细根数和总根数仅产生单独影响(表1)。克隆整合显著增加了2 种植物先端分株、基端分株的细根数,总根数及莲子草先端分株、基端分株的粗根数,且无论有无克隆整合,莲子草先端分株、基端分株的粗根数、细根数和总根数均显著多于空心莲子草(图3a)。

图3 克隆整合处理对2 种植物先端分株和基端分株根系分配及生物量分配的影响Fig.3 Effects of clonal integration on root system allocation and biomass allocation of apical and basal ramets of two plants
植物种类和克隆整合处理对2 种植物先端分株的地上生物量产生显著的交互影响,但仅对地下生物量、总生物量或根冠比产生单独影响(表1)。无克隆整合时,2 种植物先端分株的地上生物量、总生物量及基端分株的地下生物量无显著差异。
与无克隆整合处理相比,克隆整合显著增加了空心莲子草先端分株的地上生物量、总生物量和基端分株的总生物量,从而使其显著大于莲子草;克隆整合显著增加了空心莲子草基端分株的地上生物量和总生物量,且无论有无克隆整合,空心莲子草基端分株的地上生物量和总生物量均显著大于莲子草;克隆整合虽显著增加了莲子草先端分株的地下生物量,但与空心莲子草相比仍无显著差异(图3b)。
2.3 克隆整合处理下2 种植物整个克隆片段根系及生物量分配情况
植物种类和克隆整合处理对2 种植物整个克隆片段的粗根数产生显著的交互影响(表1)。克隆整合显著增加了2 种植物整个克隆片段的细根数、总根数及莲子草的粗根数,但无论有无克隆整合,莲子草整个克隆片段的粗根数、细根数和总根数均显著大于空心莲子草(图4a)。植物种类和克隆整合处理对2 种植物整个克隆片段的地上生物量产生显著的交互影响(表1)。克隆整合显著增加了空心莲子草整个克隆片段的地上生物量和总生物量,从而使空心莲子草整个克隆片段的地上生物量和总生物量显著大于莲子草;克隆整合显著增加了空心莲子草的地下生物量(图4b)。
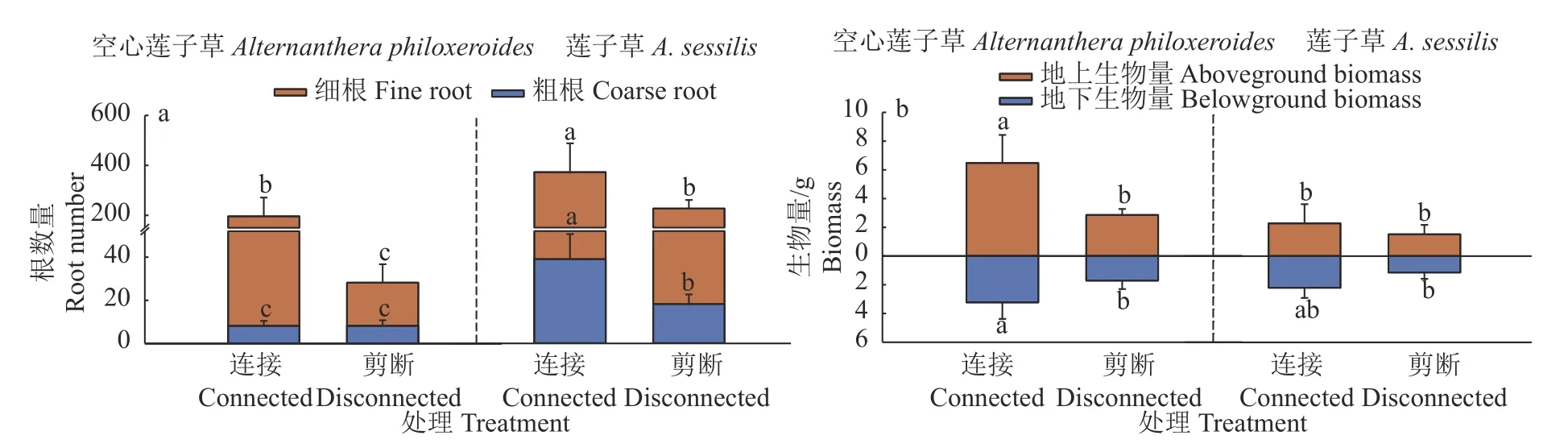
图4 克隆整合处理对2 种植物整个克隆片段根系分配及生物量分配的影响Fig.4 Effects of clonal integration on root system and biomass allocation of the whole clonal fragments in two plants
2.4 克隆整合处理下2 种植物整个克隆片段地上部分生长及光合差异
植物种类和克隆整合处理对2 种植物整个克隆片段的叶片数产生显著的交互影响(表1)。无克隆整合时,莲子草整个克隆片段的叶片数显著多于空心莲子草,然而克隆整理处理下,2 种植物整个克隆片段的叶片数无显著差异(图5a)。克隆整合显著增加了空心莲子草的茎长(图5b)。
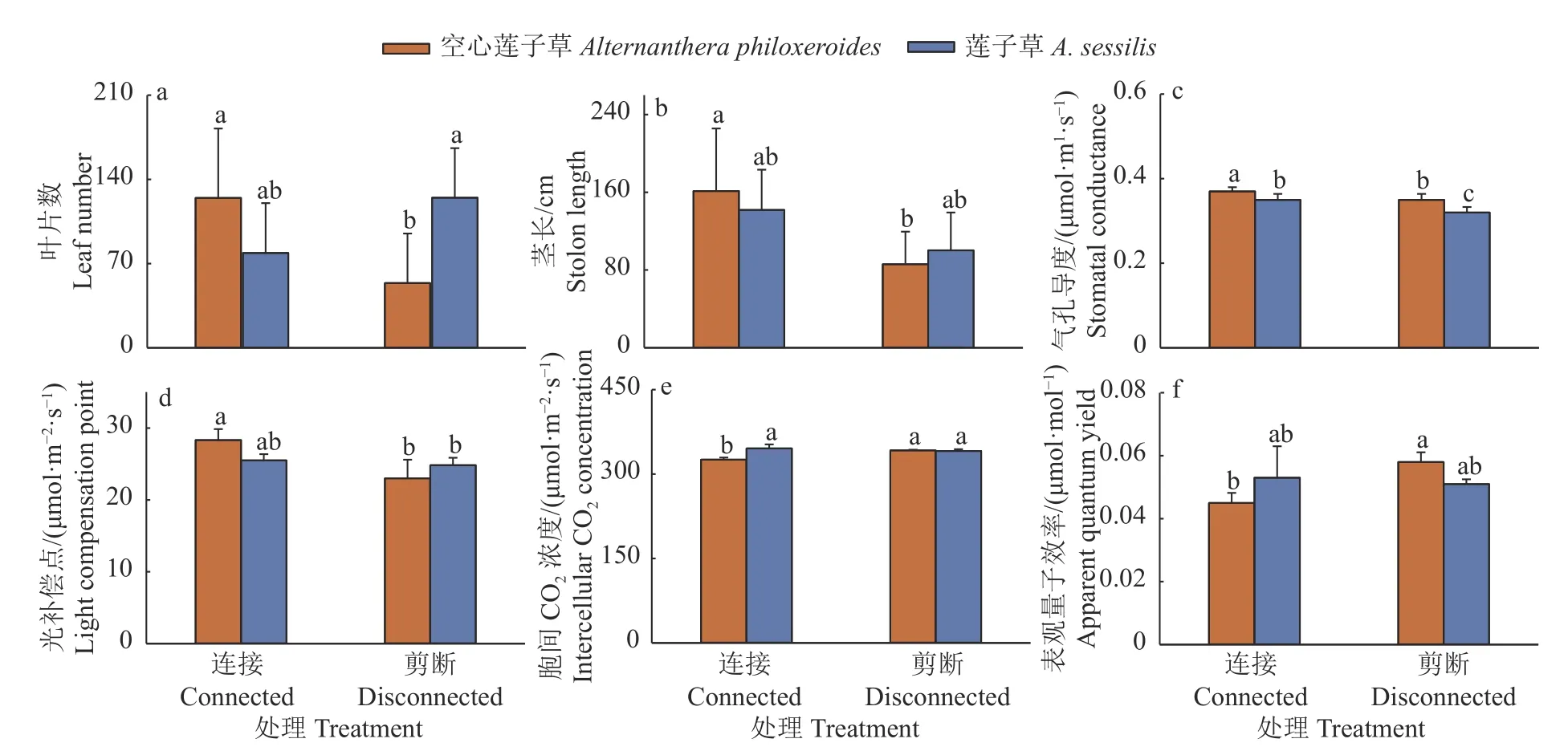
图5 克隆整合处理对2 种植物整个克隆片段地上部分生长及光合的影响Fig.5 Effects of clonal integration on growth and photosynthesis of aboveground part of clone fragments in two plants
植物种类和克隆整合处理对2 种植物整个克隆片段的胞间CO2浓度、表观量子效率和光补偿点产生显著的交互影响(表1)。克隆整合显著增加了2 种植物整个克隆片段的气孔导度和空心莲子草整个克隆片段的光补偿点(图5c、5d),且无论有无克隆整合,空心莲子草整个克隆片段的气孔导度均显著大于莲子草(图5c)。克隆整合显著降低了空心莲子草整个克隆片段的胞间CO2浓度和表观量子效率,从而使空心莲子草整个克隆片段的胞间CO2浓度和表观量子效率小于莲子草(图5e、5f)。克隆整合处理对2 种植物整个克隆片段的净光合速率无显著影响(表1),但无论有无克隆整合,莲子草整个克隆片段的净光合速率要显著低于空心莲子草,分别低19.3%和25.3%。
3 讨论与结论
许多研究都表明克隆整合对克隆植物的生长有促进作用[9-10]。本研究中,克隆整合显著增加了空心莲子草先端分株的茎长、细根数、总根数、地上生物量、总生物量以及莲子草先端分株的粗根数、细根数、总根数、地下生物量,说明空心莲子草和莲子草都可以在一定程度上从克隆整合中受益。在高土壤养分条件下,克隆整合增加了薇甘菊Mikania micranthaKunth 先端分株和基端分株的生物量[10],而其他研究发现克隆整合只能促进克隆植物先端分株的生长[24-25]。这可能是因为克隆植物通过克隆整合将碳水化合物从基端分株运输至先端分株,相对于没有克隆整合的植株获得更大的生长优势,从而促进整个分株的生长[9,24-25]。
本研究发现克隆整合显著增加空心莲子草先端分株的地上生物量及其基端分株的地下生物量。在具有充足资源的同质环境中,莫邪菊Carpobrotus edulis的先端分株和基端分株之间存在类似的分工:在克隆整合处理下,先端分株将更多的生物量分配给地上部分,而基端分株将更多的质量分配给根系[26]。胡安安[27]的研究也表明在低光照、高养分处理下,空心莲子草的基端分株将更多的生物量分配到地下部分。这表明克隆整合在同质环境下也能使克隆植物受益[26]。但本文中的莲子草并未发现类似的现象,说明不同的克隆植物所用的克隆整合策略是不同的。
气孔控制着叶片与大气间的H2O、CO2交换,植物可根据环境因素调整孔径[28]。气孔导度是气孔张开程度的量度,在植物光合作用、呼吸作用、蒸腾作用中扮演重要角色[29-30]。多数情况下,蒸腾速率以及气孔导度都与净光合速率呈极显著正相关[31]。本文中,克隆整合显著增加了空心莲子草先端分株的气孔导度、蒸腾速率以及莲子草的蒸腾速率,说明克隆整合在一定程度上增加了空心莲子草和莲子草的光合性能,从而获得更高的生长速度和定植能力。有研究发现克隆整合植株的光合性能优于无克隆整合植株[26]。此外,本文还发现克隆整合处理下,空心莲子草先端分株的蒸腾速率显著大于莲子草,说明空心莲子草比莲子草更能从克隆整合中获益。有研究也提出外来入侵克隆植物与本地克隆植物相比具有更强的克隆整合能力[5]。
本文结果表明空心莲子草和莲子草均能在一定程度上从克隆整合中受益。但在克隆整合处理下,空心莲子草先端分株的地上生物量和总生物量以及基端分株的地上生物量、地下生物量和总生物量均显著增加,而莲子草的这些指标均无显著变化,说明空心莲子草的克隆整合能力要强于莲子草,在自然生境中能通过克隆整合作用挤占莲子草的空间生态位,从而形成竞争优势。
