穗期高温处理赣早籼58与周南稻杂交F2:3家系对农艺性状的影响
2023-06-30黄伦霄吴佳宏秦鱼河韩佳佳蒋菲魏红宇陈云澈彭春霖向警赵正武
黄伦霄 吴佳宏 秦鱼河 韩佳佳 蒋菲 魏红宇 陈云澈 彭春霖 向警 赵正武

摘要:为研究高温环境对抽穗开花期杂交水稻主要农艺性状的影响,以常规早稻赣早籼58号为母本和热敏感材料周南稻为父本构建杂交F2:3家系,在抽穗开花期对F2:3家系进行7 d高温胁迫处理,并在大田设置同期对照。结果表明,F2:3家系在抽穗开花期遭受高温胁迫后,高温组相较对照组株高、穗长、每穗实粒数、每穗总粒数、着粒密度和结实率平均值显著下降,每穗空粒数显著上升;产量相关农艺性状中亲优势提高,其中每穗实粒数中亲优势明显;每穗实粒数和结实率的变异系数明显上升,每穗空粒数变异系数下降。穗长、每穗总粒数、每穗空粒数与处理期相对湿度呈显著负相关,平均温度和时积温与相对湿度呈极显著负相关,湿度与温度共同影响抽穗开花期F2:3家系的生殖生长。以胁迫温度高于38 ℃的时积温作为高温危害阈值对F2:3家系进行热害状况评估,随时积温增加,热害程度加重,相对结实率下降。
关键词:杂交水稻;高温;农艺性状;抽穗开花期
中图分类号:S511.034文献标志码:A
文章编号:1002-1302(2023)05-0103-07
水稻是我国种植面积最大、单产最高、总產最多的第一大粮食作物,在我国的粮食生产中有着举足轻重的地位[1]。近年来,在全球气候变暖、温室效应加剧的大背景下,短期异常高温频繁发生,对我国水稻生产造成了严重的不利影响[2]。2006年6—8月,重庆市出现历史罕见高温热浪和百年一遇的伏旱,农作物受旱面积超过132.7万hm2,绝收面积为37.5万hm2,粮食减产500万t左右,直接经济损失达90.7亿元[3]。2003年,我国长江流域种植的杂交水稻超过40.5万hm2,因受持续高温影响导致水稻产量大幅度下降,平均减产2.25 t/hm2[4]。郭安红等分析1981—2016年早稻高温热害强度指数的变化趋势表明,江南和华南大部分地区6月下旬、7月上旬和整个抽穗至灌浆期高温热害强度指数呈现增强趋势,高温热害已成为影响水稻产量的主要因素之一[5]。
已有研究表明,水稻对高温最敏感的时期是抽穗开花期,此时遭到高温胁迫可能造成小花退化或花粉不育,以及花药开裂散粉受阻,引起受精率下降,从而造成水稻产量下降[6-11]。此外,水分和相对湿度在高温下同样影响水稻产量,且对耐热性不同的品种产生的影响程度具有差异[9,12-13]。王彤彤等研究表明,不同品种水稻各农艺性状受高温胁迫的变化程度不一,但都具有花期提前的特征,且热敏感品种受高温影响的幅度要大于耐热品种[14]。周浩等研究抽穗扬花期高温对水稻重组自交系群体RIL47结实率的影响,发现高温处理下RIL47群体结实率极显著下降,并以热害指数为指标筛选耐高温株系[15]。目前在耐高温相关种质资源的筛选和QTL定位等方面,常用人工气候室控制恒定高温进行胁迫处理[16-18];在自然高温对水稻的影响方面,则通常利用气象资料对高温地区的水稻产量进行统计分析[19-20];对高温诱导水稻不育机理进行研究时,则通常以特定耐热或热敏感品种作为材料[21-23]。本研究以赣早籼58号和周南稻作为亲本,杂交获得具有抽穗期早晚差异和耐热性差异的F2:3家系为供试材料,通过温室模拟自然高温,研究抽穗开花期高温对F2:3家系农艺性状的影响,分析高温胁迫下F2:3家系主要农艺性状的遗传变异及与温湿度的相关性,比较不同高温阈值参数与高温下产量相关农艺性状的关联程度,以期为水稻耐热品种评价及选育提供参考。
1 材料与方法
1.1 试验地点与供试材料
本试验以热敏感籼稻品种周南稻作为父本(P1),常规早稻品种赣早籼58号作为母本(P2),亲本材料由江西省农业科学院提供,构建杂交F2代群体,从中筛选出206个F2:3家系,编号 1~206。
2018年3月10日,父本、母本和F2:3家系播种于重庆市璧山区来凤镇重庆师范大学水稻试验基地,4月26日,每个家系和亲本移栽于内径25 cm、高17 cm的盆钵,在重庆师范大学校内水稻温室基地室外进行培养,每个盆钵栽3个单株,分成2组,一组作为后续进行连续7 d高温处理组(high temperature treatment group,HT),另一组作为自然对照组(natural temperature treatment group,NT)。2个处理均按正常田间肥水管理,土壤肥力中等,在水稻全生育期严格控制病虫害,8月收获考种。
1.2 高温处理方法
在水稻进入抽穗开花期时(以稻穗顶部露出叶鞘外为标准)[11]将盆钵移入人工玻璃温室中,在抽穗开花期间对其进行为期7 d的高温胁迫处理,处理期间每天10:00—14:00使用空调加热升温营造高温环境,每日光照时间为12 h,水肥条件与对照组一致。随后将盆钵移出温室,按正常田间水肥管理,直至收获。由于F2:3群体内始穗期具有株系间差异,在达到抽穗开花期标准时依次进行高温处理,将处理日期相同的家系归为一组。
1.3 测定项目与方法
1.3.1 主要农艺性状测定
成熟后测量每个盆钵3株水稻的株高(plant height,PH)、有效穗数(productive ear number,PEN)、穗长(panicle length,PL)、每穗实粒数(filled grains per panicle,FGP)和每穗总粒数(total grains per panicle,TGP),每株随机选取3个有效穗考种取平均值。为便于讨论,高温组前缀HT(high temperature treatment)省略,对照组数据以NT(natural treatment)前缀作为区分,下同。
1.3.2 气象资料收集
高温处理期间,使用TH11R温湿度记录仪每隔30 min记录1次温室内的温度和相对湿度(RH)。对照组温度来源于国家气象科学数据中心璧山气象数据(http://www.nmic.cn)。
1.3.3 相关参数计算
每穗空粒数(empty grains per panicle,EGP)与着粒密度(spikelet density,SD)计算公式为
EGP(粒)=TGP-FGP;
SD(粒/cm)=TGP/PL。
結实率(seed setting rate,SSR)与高温相对结实率(heat relative seed setting rate,HRSR)计算公式为
SSR=FGP/TGP×100%;
HRSR=SSR/NTSSR×100%。
中亲优势(mid-parent heterosis)计算公式为
中亲优势=(F2:3性状值-亲本平均值)/亲本平均值×100%。
2 结果与分析
2.1 高温胁迫情况及F2:3家系主要农艺性状
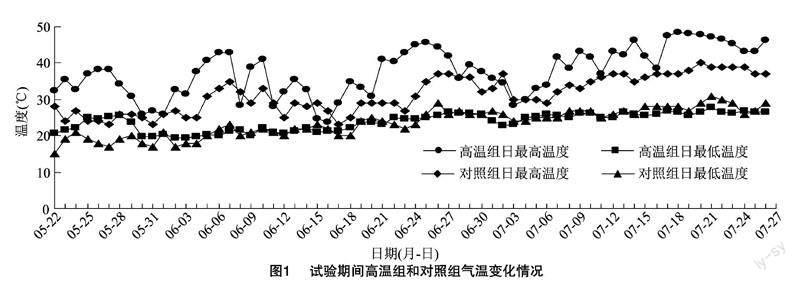
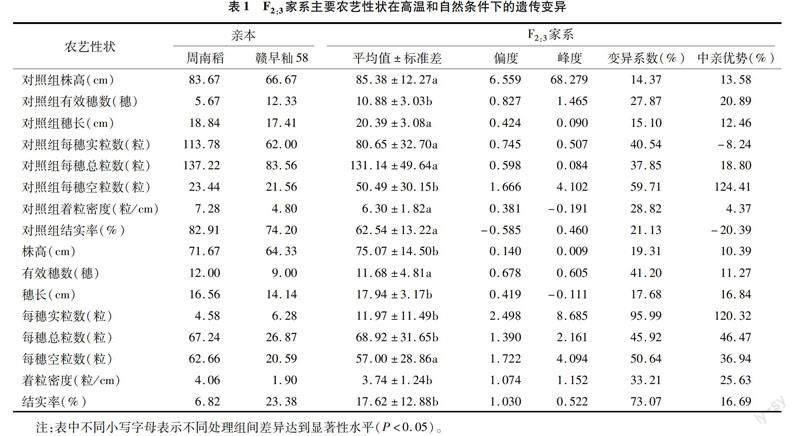
从5月22日第1批材料抽穗起至7月26日所有材料高温处理完毕,共计71 d。对温室内及对照组温度变化情况进行统计(图1),处理期间,高温组日均最高温度为37.7 ℃,日均最低温度为23.8 ℃,总平均温度为28.96 ℃;对照组日均最高温度为31.4 ℃,日均最低温度为23.6 ℃。相关性分析表明,处理期间高温组日均最高温度与对照组呈极显著正相关关系(皮尔逊相关系数为0.700,P<0.01),即高温组与对照组的同期最高温度走势一致,日均最高温度高温组超出对照组6.3 ℃。此外,处理期间高温组日极端高温大于32、35、38 ℃的天数分别为58、46、37 d。经统计,高温组和对照组共有30个株系未正常生长或抽穗,最终考种有效F2:3家系176个,2个亲本表现正常。对F2:3家系农艺性状的有效数据进行统计分析(表1),配对样本t检验(students test)表明,高温组株高(PH)、穗长(PL)、每穗实粒数(FGP)、每穗总粒数(TGP)、着粒密度(SD)和结实率(SSR)相对对照组均出现显著下降,有效穗数(PEN)和每穗空粒数(EGP)则显著上升;F2:3家系主要农艺性状在自然和高温条件下均具有明显的性状分离,对照组F2:3家系的株高(NTPH)、有效穗数(NTPEN)、穗长(NTPL)、每穗总粒数(NTTGP)、每穗空粒数(NTEGP)和着粒密度(NTSD)呈现中亲优势,其中NTEGP中亲优势最为明显,F2:3家系平均值达124.41%;高温组F2:3家系8个农艺性状均呈现中亲优势,且PL、FGP、TGP、SD、SSR中亲优势相较对照组提高,其中FGP中亲优势最为明显,F2:3家系平均值达120.32%。在8个农艺性状中,对照组和高温组的株高、穗长变异系数小于20%,其余农艺性状(PEN、FGP、TGP、EGP、SD、SSR)变异系数均超过20%。高温组与对照组相比,除EGP变异系数下降9.07百分点,其他农艺性状变异系数均有不同程度上升,其中FGP和SSR变异系数增加尤为明显,分别达到55.45百分点和51.94百分点。
由表1可知,高温引起空粒数、有效穗数显著增加,株高、穗长、每穗实粒数、每穗总粒数、着粒密度和结实率显著下降;高温胁迫下F2:3家系产量相关农艺性状包括穗长、每穗实粒数、每穗总粒数、着粒密度和结实率的中亲优势普遍提高,其中每穗实粒数中亲优势最明显;高温组F2:3家系内农艺性状差异普遍增大,其中每穗实粒数和结实率变异系数大幅上升。
2.2 F2:3家系主要农艺性状遗传相关分析
2.2.1 自然条件下F2:3家系主要农艺性状遗传相关分析
对F2:3家系对照组的农艺性状进行皮尔逊相关性分析,探讨自然条件下杂交后代农艺性状特征及其线性关系(表2)。自然条件下,8个农艺性状间有23对相关系数达到极显著水平,3对达到显著水平,其余2对性状无显著相关性。其中结实率(NTSSR)与每穗实粒数(NTFGP)呈极显著正相关,与每穗总粒数(NTTGP)、每穗空粒数(NTEGP)呈极显著负相关,与着粒密度(NTSD)呈显著负相关。
2.2.2 高温胁迫下F2:3家系主要农艺性状遗传相关分析
对高温组F2:3家系8个农艺性状以及高温相对结实率(HRSR)与高温处理期间平均温度(Ta)、高温处理期间高于38 ℃的时积温(acT38)和平均相对湿度(RH)进行皮尔逊相关性分析(表3)。高温组8个农艺性状间有15对相关系数达到极显著水平,4对达到显著水平,其余9对性状无显著相关性。其中结实率(SSR)与每穗实粒数FGP)呈极显著正相关,与每穗空粒数(EGP)呈极显著负相关。穗长(PL)、每穗总粒数(TGP)、每穗空粒数(EGP)、 着粒密度(SD)与平均温度[JP+2](Ta)和acT38均呈极显著正相关,PL、TGP、EGP与相对湿度(RH)呈显著负相关,此外RH与Ta和acT38均呈极显著负相关。
2.3 高温胁迫阈值的比较分析及F2:3家系热害情况评估
F2:3家系抽穗日期从5月22日至7月20日不等,将同一日期进行高温处理的株系归为一个处理组,共计17组,对各组的高温胁迫情况统计见表4。F2:3家系内的抽穗期有较大差异,67个株系与父本周南稻同期抽穗,16个株系在2个亲本前抽穗,其余93个株系在2个亲本后抽穗。本试验中,F2:3家系间由于抽穗时间不同,高温处理期间所遭受的热胁迫程度也有所差异,计算不同温度阈值(30、32、35、38 ℃)下的时积温(acT)或高温胁迫累计小时数(CHT)与各处理组产量相关农艺性状的皮尔逊相关性,比较用于评估F2:3家系受胁迫程度的温度指标(表5)。
结果表明,处理期间平均温度和最高温度均与每穗空粒数、每穗总粒数呈显著正相关。30、32、35 ℃ 时积温均与每穗空粒数、每穗总粒数呈极显著正相关,38 ℃时积温与每穗空粒数呈极显著正相关,与每穗总粒数呈显著正相关。30、32、35 ℃高温胁迫小时数与每穗空粒数、每穗总粒数呈显著正相关,38 ℃高温胁迫小时数与每穗空粒数、每穗空粒数呈极显著正相关。其中38 ℃时积温与每穗空粒数的相关性最佳,皮尔逊相关性系数达0.779,显著性达0.003。以38 ℃时积温为自变量,每穗空粒数为因变量,使用步进法进行线性回归,回归拟合方程为y=0.107 6x+51.678 0,r2=0.607 1(图2)。
以高温处理期间平均温度(Ta)、平均最高温度(Tmax)和acT38作为参数,使用离差平方和法(ward.D2函数)对17个处理组进行层次聚类(图3),将各组按高温胁迫情况分为3个等级(表6):轻度脅迫(S1)包含父本周南稻以及122个F2:3家系,38 ℃时积温为5.46 ℃·h,平均每穗空粒数为66.64粒,平均高温相对结实率为29.61%;中度胁迫(S2)包含母本赣早籼58号以及43个F2:3家系,38 ℃时积温为46.12 ℃·h,平均每穗空粒数为75.35粒,平均高温相对结实率为28.40%;重度胁迫(S3)包含11个F2:3家系,38 ℃时积温达320.18 ℃·h,平均每穗空粒数为96.76粒,平均高温相对结实率为24.00%。可见,在抽穗扬花期,38 ℃以上的高温持续时间会显著增加F2:3家系每穗空粒数,并降低高温相对结实率。
3 结论与讨论
水稻抽穗开花期受到高温影响会使水稻花粉活力、花药开裂率、花粉萌发率以及花粉管长度显著降低,导致水稻结实率明显下降,从而影响水稻产量,并且水稻抽穗开花期被认为是水稻产量对高温最敏感的时期[24-25]。本研究以早稻赣早籼58号和热敏感材料周南稻杂交构建F2:3家系,在抽穗扬花期进行高温胁迫试验,发现水稻在抽穗开花期遭受持续高温胁迫会引起主要农艺性状包括株高、穗长、每穗实粒数、每穗总粒数、着粒密度和结实率的平均值显著下降,并显著增加每穗空粒数从而影响水稻产量,与李兴华等的研究结果[26-27]一致。高温胁迫下F2:3家系穗长、每穗实粒数、每穗总粒数、着粒密度和结实率中亲优势普遍提高,每穗实粒数中亲优势明显增大,暗示F2:3家系在高温耐受性方面表现出杂种优势;高温组高温胁迫引起F2:3家系内农艺性状差异普遍增大,其中每穗实粒数和结实率变异系数大幅上升,这可能是由于F2:3家系抽穗开花期所受胁迫程度和耐热性差异不同造成的。
此外,穗长、每穗总粒数、每穗空粒数、着粒密度与高温处理期间Ta和acT38均呈极显著正相关,王亚梁研究表明穗分化期高温影响促进叶片生长,引起穗长、株高和穗粒数减少;另一方面穗长、每穗总粒数、每穗空粒数与相对湿度呈显著负相关,Ta和acT38与相对湿度呈极显著负相关,湿度与温度共同影响抽穗开花期F2:3家系的生殖生长[28]。胡达明发现,高温高湿环境下,杂交水稻结实率提高[29];胡声博则发现高温高湿会阻碍花粉受精[9];潘永地等在不同湿度条件下水稻受精率的研究说明高温下适宜的湿度对提高颖花受精率有利[13,30]。持续高温下不同温湿度对抽穗期水稻生长的影响还有待进一步研究。
对于抽穗扬花期高温热害,通常以日平均气 温≥ 30 ℃ 或32 ℃、日最高气温≥ 35 ℃、极端高温≥38 ℃ 以及高温持续天数3 d以上等作为临界指标[5,31-33],另有研究表明,以热积温作为指标能够更好地说明高温热害对结实率的影响[20-21,34]。本研究分析了以30、32、35、38 ℃为高温危害阈值与结实率的斯皮尔曼相关性,比较发现以胁迫期间 38 ℃ 时积温(acT38)作为指标能够较好地描述高温对F2:3家系产量相关农艺性状的影响,acT38与每穗空粒数呈极显著正相关;以处理期间平均温度、平均最高温度和acT38作为指标可将F2:3群体所受热害程度分为轻度、中度和重度胁迫,随acT38增加,热害程度加重,每穗空粒数和高温相对结实率增加。
参考文献:
[1]方福平,程式华. 论中国水稻生产能力[J]. 中国水稻科学,2009,23(6):559-566.
[2]万丙良,查中萍. 气候变暖对水稻生产的影响及水稻耐高温遗传改良[J]. 中国农学通报,2012,28(36):1-7.
[3]中国气象局. 中国气象灾害年鉴[M]. 北京:气象出版社,2007.
[4]吴 超,崔克辉. 高温影响水稻产量形成研究进展[J]. 中国农业科技导报,2014,16(3):103-111.
[5]郭安红,何 亮,韩丽娟,等. 早稻高温热害强度指数构建及气候危险性评价[J]. 自然灾害学报,2018,27(5):96-106.
[6]李万成,朱启升,王云生,等. 高温胁迫条件下水稻生理生化指标与产量性状的相关性研究[J]. 中国农学通报,2013,29(9):5-10.
[7]林翠香,倪大虎,宋丰顺,等. 高温胁迫下水稻生理特性变化及适应机制研究进展[J]. 安徽农学通报,2020,26(24):37-42.
[8]赵正武,曾卓华,陈 旭,等. 自然高温胁迫对杂交水稻花粉育性和主要农艺性状的影响[J]. 江西农业学报,2009,21(8):19-21.
[9]胡声博. 水稻高温诱导颖花不育品种间差异及其机理研究[D]. 北京:中国农业科学院,2013:17-44.
[10]宋有金,吴 超. 高温影响水稻颖花育性的生理机制综述[J]. 江苏农业科学,2020,48(16):41-48.
[11]Li M M,Li X,Yu L Q,et al. Identification of QTLs associated with heat tolerance at the heading and flowering stage in rice (Oryza sativa L.)[J]. Euphytica,2018,214(4):70.
[12]段 骅,唐 琪,剧成欣,等. 抽穗灌浆早期高温与干旱对不同水稻品种产量和品质的影响[J]. 中国农业科学,2012,45(22):4561-4573.
[13]潘永地,朱世杨,胡 凝,等. B68-1转基因水稻花粉活力的温湿响应规律[J]. 中国农学通报,2014,30(30):21-27.
[14]王彤彤,张凤鸣,白良明,等. 温度对水稻生长的影响[J]. 黑龙江农业科学,2016(2):17-20.
[15]周 浩,胡文彬,王作平,等. 抽穗扬花期高温对水稻重组自交系群体RIL47结实率的影响[J]. 中国生态农业学报,2011,19(1):69-74.
[16]文绍山,张 林,焦 峻. 抽穗开花期耐热性水稻恢复系种质的筛选[J]. 贵州农业科学,2018,46(3):7-10.
[17]陈 雷,王 强,张晓丽,等. 不同水稻基因型花期耐热性鉴定与评价[J]. 南方农业学报,2021,52(10):2641-2649.
[18]陈庆全,余四斌,李春海,等. 水稻抽穗开花期耐热性QTL的定位分析[J]. 中国农业科学,2008,41(2):315-321.
[19]陈升孛,张天圣,黄秋如,等. 海南早稻抽穗乳熟期高温热害对结实率的影响[J]. 中国热带农业,2019(3):65-69.
[20]叶 佩,刘可群,申双和,等. 湖北中稻抽穗开花期高温热害风险分析及区划[J]. 中国农学通报,2022,38(8):110-117.
[21]梁天锋,王 强,陈 雷,等. 基于时积温的水稻耐热性鉴定方法研究[J]. 南方农业学报,2016,47(4):537-541.
[22]王华运,周桂香,万谦千,等. 花期高温对水稻穗、叶生理特性的影响[J]. 农业灾害研究,2020,10(4):129-131.
[23]张 彬,芮雯奕,郑建初,等. 水稻开花期花粉活力和结实率对高温的响应特征[J]. 作物学报,2007,33(7):1177-1181.
[24]Matsui T,Omasa K,Horie T. The difference in sterility due to high temperatures during the flowering period among Japonica-rice varieties[J]. Plant Production Science,2001,4(2):90-93.
[25]Jagadish S V K,Muthurajan R,Oane R,et al. Physiological and proteomic approaches to address heat tolerance during anthesis in rice (Oryza sativa L.)[J]. Journal of Experimental Botany,2009,61(1):143-156.
[26]李興华,张 盛,周 强,等. 抽穗期高温对不同品种水稻产量的影响及差异[J]. 中国农学通报,2019,35(9):1-6.
[27]曹云英,段 骅,杨立年,等. 减数分裂期高温胁迫对耐热性不同水稻品种产量的影响及其生理原因[J]. 作物学报,2008,34(12):2134-2142.
[28]王亚梁. 高温对水稻穗发育及穗部性状的影响[D]. 北京:中国农业科学院,2016:32-42.
[29]胡达明. 籼型三系不育系水稻花时滞候解除研究 Ⅱ. 花时的温湿生态因子效应[J]. 湖北农学院学报,1996(4):254-257.
[30]Julia C,Dingkuhn M. Variation in time of day of anthesis in rice in different climatic environments[J]. European Journal of Agronomy,2012,43:166-174.
[31]江 敏,金之庆,石春林,等. 长江中下游地区水稻孕穗开花期高温发生规律及其对产量的影响[J]. 生态学杂志,2010,29(4):649-656.
[32]杨 军,章毅之,贺浩华,等. 水稻高温热害的研究现状与进展[J]. 应用生态学报,2020,31(8):2817-2830.
[33]刘 佳,陈 超,张玉芳,等. 四川单季稻抽穗扬花期和灌浆结实期高温热害时空特征[J]. 中国农业气象,2018,39(1):46-58.
[34]陈 雷. 水稻花期耐热性与耐热QTL定位及候选基因分析[D]. 南宁:广西大学,2021:30-75.
收稿日期:2022-04-27
基金项目:国家自然科学基金(编号:31670326);重庆市技术创新与应用发展专项(编号:cstc2019jscx-msxm X0353);重庆高校优秀成果转化项目(编号:KJZH17114)。
作者简介:黄伦霄(1996—),男,四川攀枝花人,硕士研究生,主要从事水稻生理学研究,E-mail:1164158792@qq.com;共同第一作者:吴佳宏(1996—),女,重庆人,博士研究生,主要从事生物进化研究,E-mail:wujiahong231310@gmail.com。
通信作者:赵正武,博士,研究员,主要从事水稻遗传育种研究。E-mail:zhaozhengwu513@sina.com。
