极小种群云南梧桐的群落特征研究
2023-06-20茶武婧石明杜凡王粤戴
茶武婧石 明杜 凡王 粤戴 蓉
(1. 西南林业大学林学院,云南 昆明 650233;2. 生态环境部南京环境科学研究所,江苏 南京 210042)
云南梧桐(Firmiana major)是梧桐科(Sterculiaceae)梧桐属(Firmiana)乔木,国家Ⅱ级重点保护野生植物与极小种群物种[1-2],分布于金沙江流域上游至中游河谷。1991 年出版的《中国植物红皮书》[3]中记载,由于云南中部地区持续不断的开发造成其生境被破坏,云南梧桐的野生植株几乎绝迹。1998 年被IUCN 确定为野外灭绝。2001 年后在云南的丽江、元谋、武定、禄劝、华坪、香格里拉及四川攀枝花均零星发现云南梧桐分布点[4-5]。
目前对云南梧桐的研究依然很少,仅有云南梧桐种群[4,6-7]、叶片结构[8]、叶绿体基因[9-10]和人工繁育[11]等少量文献。本研究根据近期发现于香格里拉的原生云南梧桐群落的调查数据,综合多年来各地的调查、相关文献和对标本的考证,归纳和分析了云南梧桐的地理分布、群落结构、区系成分、种群特征和濒危原因,为深入研究和有效保护云南梧桐提供参考依据。
1 研究区自然概况
2020 年5 月,在香格里拉洛吉乡东南部的尼汝峡谷发现了目前个体数量最多的云南梧桐种群。该区位于滇西北与川西交界处,地处北纬27°47′49.48″,东经100°19′11.33″,海拔1 798~2 050 m。峡谷呈西北—东南走向,纬度接近28°,谷深超过400 m,为深切的U 型谷,两侧山坡陡峭,谷底狭窄,河水湍急。在此环境下太阳正午才能短暂照进峡谷内,所以峡谷内具有日照短、蒸发量小而湿润环境。这里地处藏区人烟稀少,加之陡峭地形的限制,云南梧桐群落基本无人为干扰。云南梧桐集中分布于峡谷左岸,沿河谷分布长度约3.5 km,从河谷边沿悬崖上升约200 m,其株行距为2~10 m,估算群落面积约0.7 km2,云南梧桐个体数量超过10 000 株(图1)。

图 1 香格里拉尼汝峡谷云南梧桐生长情况Fig. 1 Growth of F. majorin Niru Valley of Shangri-La
2 研究方法
2.1 样方设置与调查
调查时间为2020 年5—6 月,共调查2 次。受陡峻悬崖生境的限制仅能选取3 个15 m ×15 m 相对平缓且可达的样方进行调查。记录样方环境因子、群落特征、云南梧桐种群特征及其开花、结实、更新情况、干扰因素,采用相机和无人机进行辅助调查。
2.2 重要值分析[12]
2.3 标本查阅
查阅CVH 中国数字植物标本(http://www.cvh.org.cn/)、NSII 国家标本资源共享平台和Global Plants (http://plants.jstor.org/)网络资源。结果表明,云南梧桐标本收藏于国内外5 个标本馆,共17 号:昆明植物所(KUN)存7 号,北京植物所(PE)存6 号,华南植物园(IBSC)存1 号,西双版纳热带植物园(HITBC)存1 号,北京自然博物馆(BJM)存1 号,英国爱丁堡皇家植物园(BM)[13]存1 号为模式标本(G.Frest10820,1913 年8 月,中国长江流域)。除模式标本外其他标本多采集自路边、公园、庙内等生境,部分标本采集生境不详。
3 结果与分析
3.1 地理分布特征
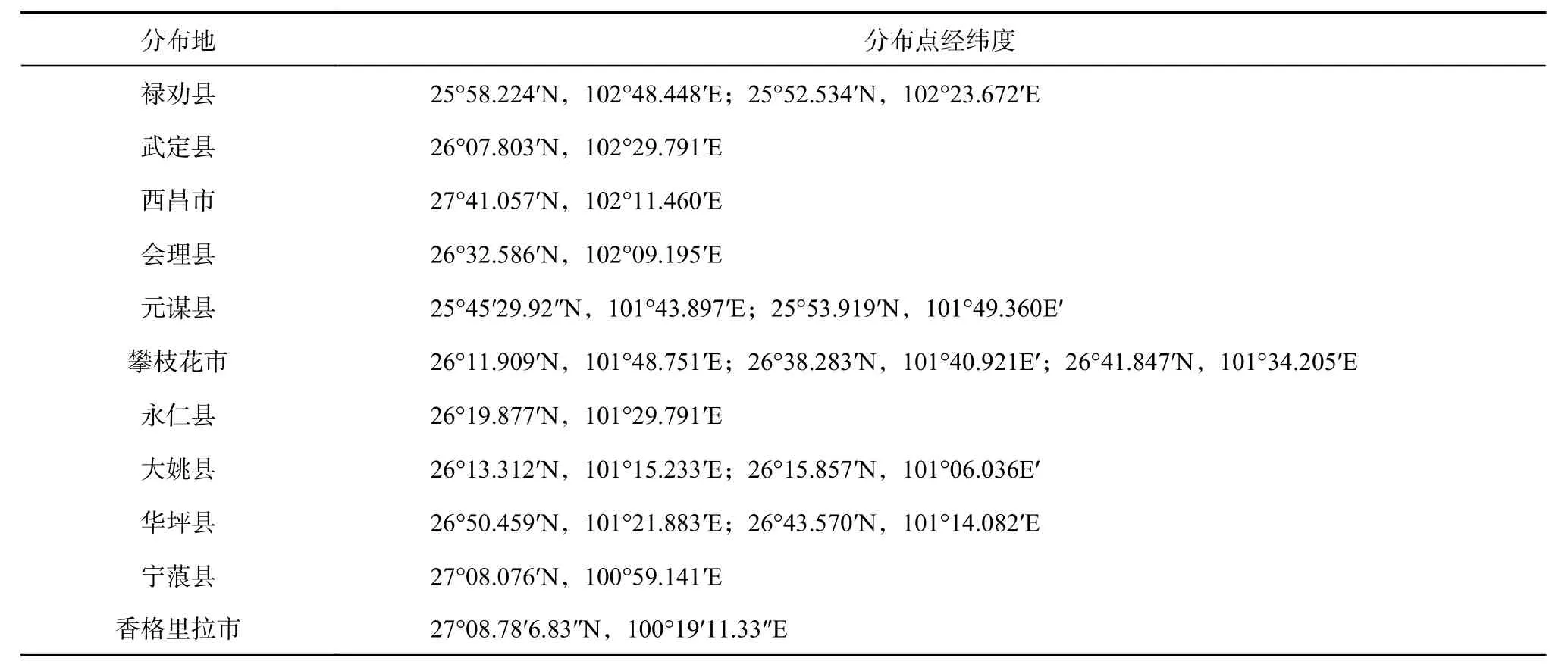
表 1 云南梧桐分布点Table 1 Distribution of F. major
3.2 群落区系组成与地理成分
香格里拉云南梧桐群落的物种较少,3 个样方共记录维管植物39 科70 属80 种,其中蕨类植物6 科8 属11 种;被子植物33 科62 属69 种。属的地理成分具有明显的热带性质,热带成分占58.06%,温带成分占40.32%。种地理成分则具有明显的温带性质,温带成分61 种,占88.41%。但从种的分布区类型看,植物区系受古地中海退却后的影响较大,该群落中的热带残遗分布类型中国—喜马拉雅分布、中亚至喜马拉雅和华西南分布的物种分别占群落种数的15.94%和1.45%。
3.2.1 属的数量和分布区类型分析
群落样方记录种子植物62 属,含2 种以上(包括2 种)的属计5 属含12 种,出现1 种的属有58 属,占群落全部属数的93.55%,全部种数的84.06%。说明出现1 种的单种属构成了本群落植物区系多样性的主体成分。
依照《中国种子植物属的分布区类型》[17],62 属可划分为13 个分布区类型和6 个变型,69种可划分为13 个分布区类型和8 个变型(表2)。
世界广布属1 属,占总属数的1.61%,为铁线莲属(Clematis);热带属36 属,占总属数的58.06%;温带属25 属,占总属数的40.32%。从属级水平看,群落与热带植物区系的联系以泛热带成分为主,与温带植物区系的联系以北温带成分为主。其中古地中海退却后的残遗属[17]计12 属,除梧桐属(Firmiana)外,有香薷属(Elsholtzia)、萱草属(Hemerocallis)、鹅绒藤属(Cynanchum )、 栎 属(Quercus )、 桑 属(Morus )、 葡 萄 属(Vitis )、 盐 肤 木 属(Rhus)、风铃草属(Campanula)、风轮菜属(Clinopodium)、黄精属(Polygonatum)、天南星属(Arisaema)。
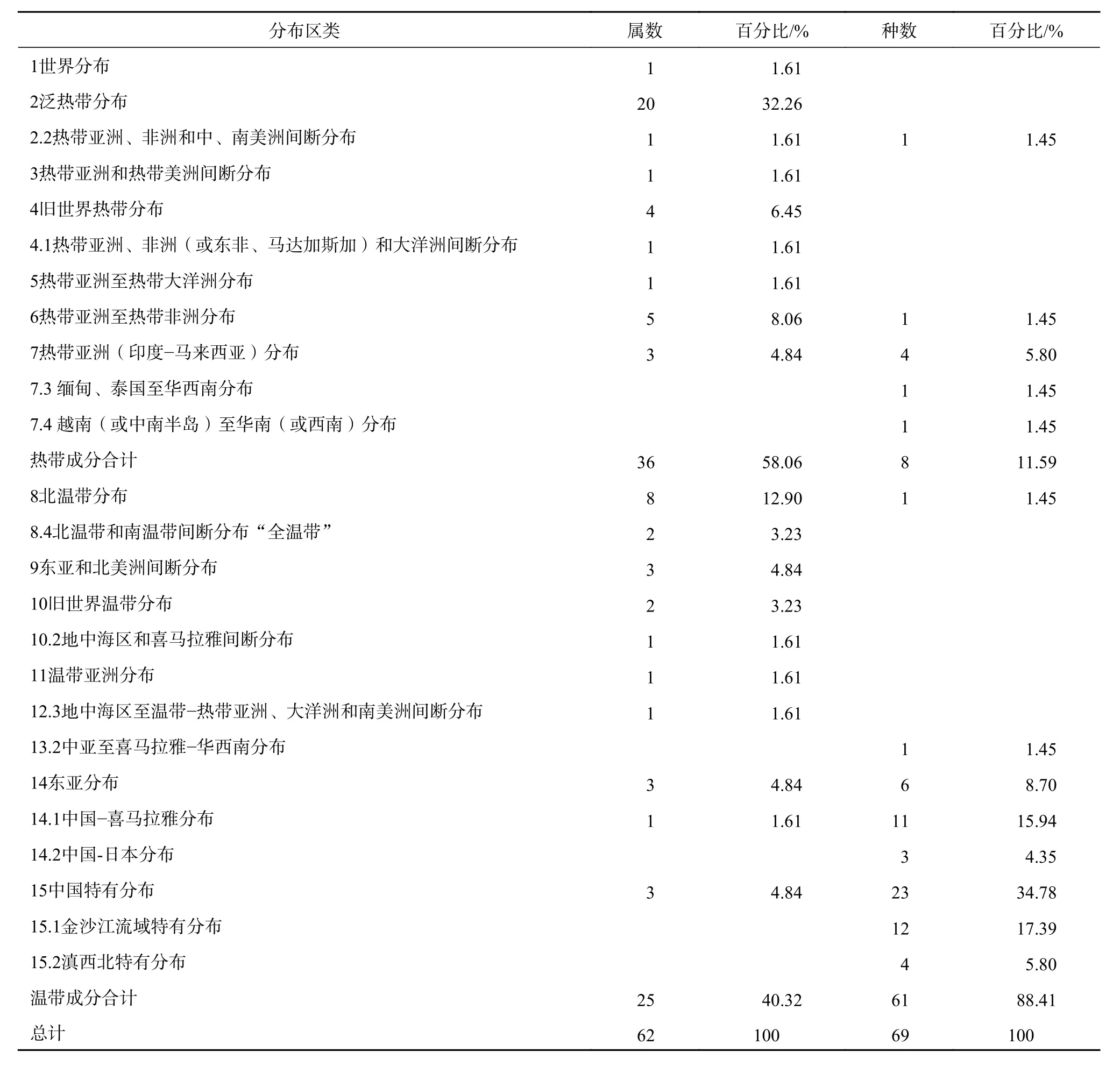
表 2 香格里拉云南梧桐群落种子植物属和种的分布区类型Table 2 The distribution types of genera and species of seed plants in F. majorcommunity in Shangri-La
3.2.2 种的分布区类型
1)群落无世界广布种。热带种8 种,占群落种数的11.59%。以热带亚洲(印度—马来西亚)分布较多,共4 种,为茎花苎麻(Boehmeria clidemioides)、鸡矢藤(Paederia scandens)、假杜鹃(Barleria cristata)、尼泊尔芒(Miscanthus nepalensis)。其余为热带亚洲至热带非洲分布、泛热带分布变型和热带亚洲分布变型,即臭灵丹(Laggera pterodonta)、长穗小草(Microchloa indicavar. kunthii)、心叶秋海棠(Begonia labordei)、茶条木(Delavaya yunnanensis)。
2)温带种61 种,占群落种数的88.41%。以中国特有分布及其变型最多,计39 种。其中正型23 种, 如 扫 把 竹(Drepanostachyumfractiflexum)、毛叶柿(Diospyrosmollifolia)、雅致雾水葛(Pouzolziaelegans)、铁橡栎(Quercus cocciferoides)、红麸杨(Rhuspunjabensisvar.sinica)、丽 江 铁 苋 菜(Acalyphaschneideriana)、齿唇铃子香(Chelonopsisodontochila)、西南铁线莲(Clematispseudopogonandra)、西南木蓝(Indigoferamairei)等;变型金沙江流域特有种12 种,即云南梧桐(Firmiana major)、云南金合欢(Acacia yunnanensis)、丽江羊蹄甲(Bauhinia bohniana)、滇 榄 仁(Terminaliafranchetii)、银叶铁线莲(Clematisdelavayi)、光柱旱地木槿(Hibiscusaridicolavar.glabratus)、灰 岩 木 蓝(Indigoferacalcicola)、滇 荆(Vitexyunnanensis)、云南龙眼独活(Araliayunnanensis)、干生铃子香(Chelonopsissiccanea)、小叶鹅绒藤(Cynanchumanthonyanum)、 丁 茜(Trailliaedoxagracilis);另一变型滇西北特有种4 种,即网脉唐松草(Thalictrum reticulatum)、云南巴豆(Croton yunnanensis)、西南杭子梢(Campylotropis delavayi)白毛野丁香(Leptodermis rehderiana)。
利用角色动画表现这样一种引领式的交互方式将设计理念中重要的部分呈现出来,当进入到室内环境中,角色的任务也就此完成,从而让用户成为唯一的角色。
东亚分布及其变型计20 种,占温带种的32.79%,以中国—喜马拉雅分布的种占多数,计11 种,如绣球藤(Clematis montana)、海漆(Excoecaria acerifolia)、西南叶下珠(Phyllanthus tsarongensis)、毛叶合欢(Albizia mollis)、飞蛾藤(Porana racemosa)、印缅石蝴蝶(Petrocosmea parryorum)、长毛香薷(Elsholtzia pilosa)、旱茅(Schizachyrium delavayi)等;正型东亚分布6 种,即叶底珠(Flueggea suffruticosa)、黑弹树(Celtis bungeana)、蒙桑(Morus mongolica)、清香木(Pistacia weinmannifolia)、匍匐风轮菜(Clinopodium repens)、薯蓣(Dioscorea opposita);中国—日本分布变型3 种,即莸(Caryopteris divaricata)、天南星(Arisaema heterophyllum)、三毛草(Trisetum bifidum)(表1)。群落中北温带分布和中亚至喜马拉雅-华西南分布各1 种,分别为黄茅(Heteropogoncontortus)和西南风铃草(Campanula pallida)。
群落热带属的分布区类型比热带种的分布区类型多,说明群落物种区系深受古地中海退却、喜马拉雅抬升的影响而保留较多热带属的残遗。
3.3 群落特征
调查的群落分布于峡谷两侧陡峭的石灰岩峭壁,岩石裸露度达90%以上,坡度大于45°。群落没有乔木层,灌木层盖度35%~45%,高1~5 m,种类37 种,优势种为扫把竹(Drepanostachyum fractiflexum)和云南梧桐(Firmiana major),其重要值分别为7.45 和5.06。云南梧桐多为丛状,树干弯曲。其他重要值较大的是滇榄仁(Terminalia franchetii)、清香木(Pistacia weinmannifolia)、丽江羊蹄甲(Bauhinia bohniana)。草本层盖度20%~25%,高0.1~1 m,种类30 种,以禾本科的黄茅(Heteropogon contortus)为优势,其重要值为4.57。其他重要值较大的草本植物是印缅石蝴蝶(Petrocosmea parryorum)、粉背蕨(Aleuritopteris pseudofarinosa)、三毛草(Trisetum bifidum)、 缘 毛 卷 柏(Selaginella compta)。层间层种类13 种,高0.1~2 m,以绣球藤(Clematismontana)占优势,其重要值为1.59。其他重要值较大的是红素馨(Jasminum beesianum)、云南粘山药(Dioscorea yunnanensis)、钝萼铁线莲(Clematis peterae)、崖爬藤(Tetrastigma obtectum)。表3 为香格里拉云南梧桐群落中各层重要值较大的前5 种物种。
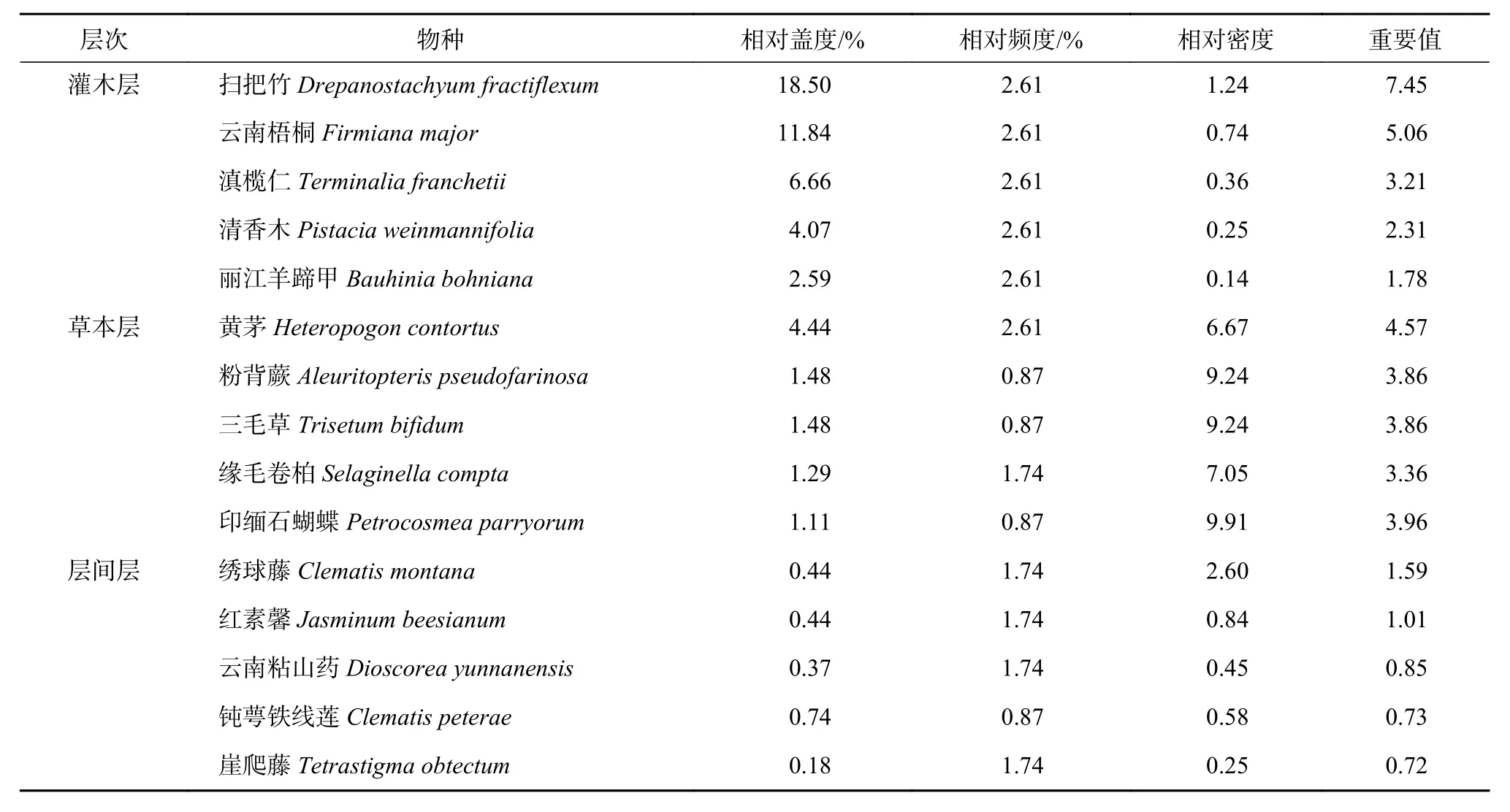
表 3 香格里拉云南梧桐群落物种重要值Table 3 Species importance of F. majorcommunity in Shangri-La
3.4 种群开花结实状况
云南梧桐野生种群开花结实量少,自然更新率极低。野外调查表明,云南梧桐5 月开花、6 月结实。云南梧桐为离生雌蕊,每个雌蕊有5 个离生心皮,每心皮有5~6 个胚珠,但最终仅有1~3 粒发育成种子,其果实为典型的蓇葖果,成熟时沿腹缝线开裂;每100 株云南梧桐植株有1~2 株开花。开花植株中每株有花枝1~2 枝,每花枝着花约50 朵,每株50~100 朵,每花枝最终结果3~4 个,每株结果3~8 个,每花最终形成1~3 个蓇葖果,每株最终有3~24 个蓇葖果,每个蓇葖果中约有1~3 粒成熟种子,每株约有3~72 粒种子;每100 株云南梧桐开花约200 朵,结果16 个,座果率为8%,每100 株云南梧桐最终有48 个蓇葖果,每个蓇葖果中有1~3 粒成熟种子,最终每100 株云南梧桐产生144 粒种子,结实率为1.44%。其种子球形,直径6~7 mm,主要靠重力散布。调查中仔细检查群落下方区域均未发现掉落的云南梧桐种子和实生苗。
4 结论与讨论
4.1 结论
本研究综合作者多年调查、标本记录、《云南植物志》[14]《Flora of China》[15]《中国植物志》[16]、Li 等[4]、王大绍[5]及中国科学院昆明分院官网等报道,得出云南梧桐零星分布于25°68′18″~27°78′6.83″N 和100°19′11.33″~102°44′E 之间,从东向西分布县域依次为禄劝、武定、西昌、会理、元谋、攀枝花、永仁、大姚、华坪、宁蒗、香格里拉、木里。同时根据对云南梧桐种群特征的研究得出:云南梧桐在群落中重要值最大达5.06,为该群落的优势树种。但云南梧桐野生种群座果率为8%,结实率仅1.44%,种群表现出自然更新率低的特点。因此,云南梧桐种群表现出自然更新率低且在金沙江流域内呈零星分布的特点。
4.2 讨论
基于云南梧桐自然更新率低和在金沙江流域内呈零星分布的特点,其濒危原因讨论如下。
1)云南梧桐特殊的残遗性是造成其在流域内分布区域零星分布的主要原因。梧桐属为白垩—老第三纪古热带的山地亚热带孑遗[18],第三纪古青藏高原隆起后,原来旱生的地中海成分随着青藏高原的隆升分化形成了中亚成分、地中海—西亚—中亚成分、旧世界温带成分、北温带成分、中国喜马拉雅成分等,并形成了间断、残遗现象[19-20]。群落中的绣球藤、海漆、毛叶合欢、飞蛾藤、印缅石蝴蝶、旱茅等中国—喜马拉雅成分占比达15.94%,且位居第二,也说明该群落起源古老为地中海的残遗类型。青藏高原的隆升起到了屏障作用,使其受第四纪冰川的影响较小,促使云南梧桐在流域内适宜生境得以保存下来。
2)云南梧桐开花结实量少加之啮齿动物取食是使其种群自然更新率低的原因,同时也是其濒危原因之一。云南梧桐种子产量少,座果率为8%,结实率仅1.44%。其次,云南梧桐种子成分与梧桐相似,富含蛋白质、油脂等而被松鼠、老鼠等啮齿动物取食[7]。种子圆球型,豌豆大小,靠重力传播,散布范围有限。调查中在群落下方和周边开阔地带均未发现种子和实生苗。分析原因可能是滚落散布到平坦开阔地带的种子均被动物捡食,仅少数落入石缝中、动物取食不了的种子才有机会萌发更新。这是在云南梧桐成年植株下方和周边未能发现其种子和实生苗的主要原因,也是云南梧桐主要生存于石缝中的原因。同时遗传多样性下降、授粉成功率不足导致的生殖力下降,种子质量差、种子损失导致的种子向幼苗的转化率低也是造成云南梧桐自然更新率低的原因之一[7]。在雷彻虹等[7]对攀枝花苏铁保护区的云南梧桐种群研究中发现,保护区的野生云南梧桐几乎均为萌蘖后的植株,且无性繁殖已成为该种群现阶段保存和延续的重要方式。但作者此次调查中在香格里拉云南梧桐种群中未发现萌蘖后的植株,香格里拉云南梧桐种群是否存在无性繁殖现象,在后续研究中需完善补充。
3)云南梧桐的适宜生境在金沙江流域内较少,其生长及分布受生境限制。有关研究[6,11]虽然云南梧桐幼苗具有胡萝卜状的肉质根便于其短期对应岩石生境的干旱[7],但云南梧桐叶还不具备耐旱特征。同时许云蕾等[8]对云南梧桐叶片结构进行解剖,研究结果显示云南梧桐叶片长宽达30 cm,叶呈现出上表皮较厚,栅栏组织排列整齐,海绵组织排列疏松,气孔分布于下表皮的特点,所以从叶片结构来看云南梧桐为中生性植物而非典型的旱生性植物。此外,云南梧桐分布的金沙流域随着喜马拉雅的抬升使金沙江河谷不断深陷,焚风效应加强、干旱逐渐加剧,流域内湿润生境逐渐减少。目前金沙江流域年降雨量700~800 mm,而蒸发量是降水量的3~6 倍,在高温干旱季节可达10~27 倍[21],生境以干热河谷为主。所以流域内普遍的干热河谷并非云南梧桐生长的适宜生境,其仅能退缩于生境湿润的局部区域。
4)人类活动加剧了云南梧桐种群濒危。目前除香格里拉群落外其他种群均有一定的人为影响。现存的云南梧桐均为残存种群,延续至今的人类活动是造成云南梧桐生境进一步破坏、种群进一步减少的原因之一[3]。作者于2020 年9 月在元谋弯腰树水库发现的云南梧桐因其陡峭的地势,无法开荒耕种,在周围均是耕地的情况下保留了面积不足1 亩,约84 株,为残存次生群落片段;其次调查中发现该样地及周边存在放牧现象,怀疑在放牧中羊群取食其种子,对云南梧桐更新有一定影响。放牧取食种子还是啮齿动物取食
综上,云南梧桐为古地中海退却喜马拉雅抬升、青藏高原隆起后形成的金沙江河谷的残遗种[18],主要生存于岩石夹缝中。因为自身繁殖能力差、种子数量少,种子质量差等多重因素,加之啮齿动物对其种子取食,使其更新困难,是造成云南梧桐种群数量少且濒危的主要原因。同时因适应生境的限制和人类活动对其生境的破坏加剧了其种群濒危,现今云南梧桐在金沙江流域内属于夹缝中生存,生存现状十分担忧。
4.3 建议
云南梧桐是我国珍稀濒危保护植物,需加强对云南梧桐种群和种质资源的研究与保护。香格里拉云南梧桐种群的野外调查表明,其开花结实量少且自然更新率极低,同时在金沙江流域呈零星分布。因此,需要采取合理的保护策略,云南梧桐的种群数量和种群规模才有望得到恢复。
云南梧桐地理分布呈零星碎片化,且种群数量少。这与其所需生境有很大关系,云南梧桐主要分于金沙江流域内相对湿润的生境中,而金沙江流域绝大部分为干热河谷生境。所以对云南梧桐的保护更有效的是保护云南梧桐原有的生境和种群。保存其原有的生态面貌,是解决云南梧桐保护的首要任务。因此,对云南梧桐的保护必须从单一的物种保护转移到物种栖息地及生态系统的保护。
优先就地保护云南梧桐现有种群,重点关注没有出现在自然保护区中的保护空白地区,优先处理好这些地区经济与环境的关系,加大这些自然保留种群的管理力度。如本次调查的香格里拉云南梧桐种群是目前发现数量最多、面积最大、保存完整且无人影响的群落,具有重要的研究和保护价值。建议当地政府及相关部门可以建立保护小区,加大宣传和保护力度,并给予一定的专项资金支持。
加强科研,鼓励科研人员对云南梧桐开花传粉的研究,掌握云南梧桐繁殖过程。目前余志祥等[11]已对云南梧桐进行了人工繁育,并取得了一定的成果。接下来还可以进一步加大对云南梧桐的人工繁育,在流域内适宜生境开展种群回归,最大限度的保护云南梧桐种质资源。
