滇东南珍稀古老植物蒜头果种子结构及子叶多样性研究
2023-06-20娟普
王 娟普 甜
(1. 西南林业大学绿色发展研究院,云南 昆明 650233;2. 西南林业大学林学院,云南 昆明 650233)
云南的东南部是古生代地层出露最丰富、地质历史最古老的地区之一,在全球广受第四纪更新世冰盖侵袭时,滇东南植物区系没有发生较大动荡,从而成为第四纪冰期全球生物界的冰期“避难所”,蕴藏了大量的古老、珍稀特有物种资源[1]。特殊的地理位置、古老的地质历史和优越的气候条件,为起源古老的生物物种的生存繁衍提供了得天独厚的条件,使得滇东南成为中国三大特有植物分布中心之一的“滇东南—桂西古特有中心”的重要组成部分。隶属被子植物中双子叶植物纲原始花被亚纲檀香目铁青树科(Olacaceae)蒜头果属(Malania)的蒜头果(Malania oleifera)也成为了这一区域保留下来的中国特有的单种属的孑遗植物,仅零星分布于云南的广南县和富宁县,以及邻近的广西西部那坡县等海拔300~1 500 m 喀斯特山地的常绿阔叶混交林中,现存野生资源量有限,被列为国家II 级保护物种之一。蒜头果种仁胚乳中富含油脂(51.85% 以上)[2],是当地居民传统食用油之一。蒜头果种仁油中含有高达40% 以上的神经酸,是迄今为止发现神经酸含量最高的木本植物[3]。神经酸能够促进受损伤神经组织修复和再生,对预防老年痴呆或辅助治疗老年性认知功能障碍有独特疗效[4-5]。因此,具有重要的保护价值和潜在的医药利用与经济价值。
蒜头果的外观特征、种子萌发与驯化栽培、营养价值、脂肪酸成分及其含量等已有不少研究[2,6-7],但有关蒜头果种子及其种胚的形态与结构的研究未见报道。种子是植物长期系统演化过程的产物,种胚作为种子最重要的组成部分,是幼小植物体的雏形被保护在种子的结构中,其结构特征是稳定和保守的。种胚一般由胚芽、子叶、胚轴和胚根4 部分组成,其中子叶数目及形态通常是种胚变异最小的性状之一[8],不仅对植物生长发育甚至产量具有重要作用[9-10],而且还能反映植物的进化及其系统发育。种子植物依据子叶数量划分为单子叶植物、双子叶植物和多子叶植物三大类群,多子叶结构一般只存在于裸子植物的种胚中。对处于被子植物基部位置古老的双子叶植物—蒜头果的多子叶现象的研究,对了解蒜头果的系统地位、揭示其种群构建等生态学规律具有重要的理论意义。蒜头果分类学研究历史显示,最早的报道曾将蒜头果置于茶茱萸科[11]和樟科[12]中,至1980 年才作为铁青树科一新属和新种,有了合法的名称和明确的系统位置。蒜头果的分类研究有文献记载仅40 余年历史,本研究通过对蒜头果果实特征与结构、种子结构及天然分布于不同海拔的蒜头果的种胚子叶数量等内容的研究,为蒜头果不同种源的良种选育、种子萌发及规模化培育造林成活率与保存率低等问题奠定基础,也为更深入认识蒜头果的系统分类地位和种子植物中子叶数量的演化规律或趋势提供了更丰富的信息和数据支撑。
1 材料与方法
1.1 材料来源及取样方法
供试蒜头果果实于2020—2022 年每年的10 月采自文山州富宁县和广南县有野生蒜头果分布的乡镇。
1.2 果实和种子基本特征观测
1.2.1 果实和种子结构特征观察
随机取100 粒蒜头果果实标记编号,用游标卡尺测量已编号果实的纵径(LD-F) 、横径(TD-F) ,称量单果质量(FW)。果柄着生部位至果实顶部的距离为纵径,垂直纵径果实最宽处为横径,横径按两个方向测量2 次,取平均值。将肉质果皮与果核(种子)分离,测量每粒种子的纵径(LD-S)和横径(TD-S) ,称量每粒种子质量(SW)。果柄的脱落处到种孔的距离为纵径,垂直纵径种子最宽处为横径,横径按两个方向测量2 次,取平均值作为种子横径。果实和种子的纵径、横径测量精度为0.01 cm;质量精度为0.01 g。果皮厚度计算方法参考文献[13]。
1.2.2 果实和种子各部分含水率测定
含水量测定按GB/T 3543. 6—1995[14],用四分法随机取20 个新鲜蒜头果果实作为1 个果样,称量鲜果质量(T-FW),重复5 次,共计100 粒果实。称量每个果样果皮总质量(T-PW)和种子总质量(T-SW)。剥出被坚硬的种壳包被的胚乳和胚,按果样编号称量种壳质量(T-ScW)、胚乳质量(T-EnW )和胚质量(T-EmW)。将5 个果样的果皮、种壳、胚乳和胚分别编号后放入75 ℃的烘箱中烘干至恒质量,记录每组果样的果皮、种壳、胚乳和胚的恒重质量,即为干果质量。
1.3 胚乳和种胚观测
取1.2.1 中已编号和称重的100 粒种子,用游标卡尺测量胚乳纵径长(LD-En)和横径长(TD-En),在徕卡502 体式显微镜下测量胚长和观测子叶数量,计算胚率。
在徕卡502 体式显微镜下观察统计已编号和称重的100 粒种子中的种胚的子叶数量,分别计数100 粒种胚中子叶数量的分布。
1.4 不同海拔蒜头果种子子叶数观察
分 别 在海 拔400、600、800、1 000、1 300、1 400、1 500 m 处,各选择3 株野生蒜头果植株,每株采集果实100 粒,每株随机取10 粒,重复2 次,即每株共选取20 粒果实用于统计种胚子叶数量,每个海拔段共取样60 粒,统计每个海拔段种子中种胚的子叶数量占取样数的比例。
1.5 蒜头果种子石蜡切片制作
取蒜头果种子,剥去种壳确定种胚位置后,将包被着种胚的胚乳切成5~8 mm 见方的块段,迅速放入FAA 固定液中,并用注射器来回抽气几次,直至未见气泡产生后,放入准备好的固定液瓶中,固定液与材料体积比不小于20∶1,在固定液中固定48 h 以上。使用不同浓度的乙醇脱水,浓度由低到高,间隔为2 h,二甲苯透明6 h,浸蜡48 h 以后包埋。切片厚度为10~12 μm,展片用蛋白甘油,番红固绿二重染色,中性树胶封片。OlympusBX53 显微镜下观察并拍照。
2 结果与分析
2.1 蒜头果果实形态与结构
蒜头果果实由复雌蕊心皮发育而成核果,每年9 月底至10 月上旬果实成熟。成熟果实果皮表面光滑无毛或有时被白粉,呈绿色或灰绿色,果实内含1 粒种子,由果皮和种子2 部分组成。种子与果柄连接处的顶部形成凸起,形似一独蒜头,因此得名蒜头果(图1a~c)。种孔位于与种柄对应的另一端,种子萌发时,胚根从种孔突破坚硬的种壳。
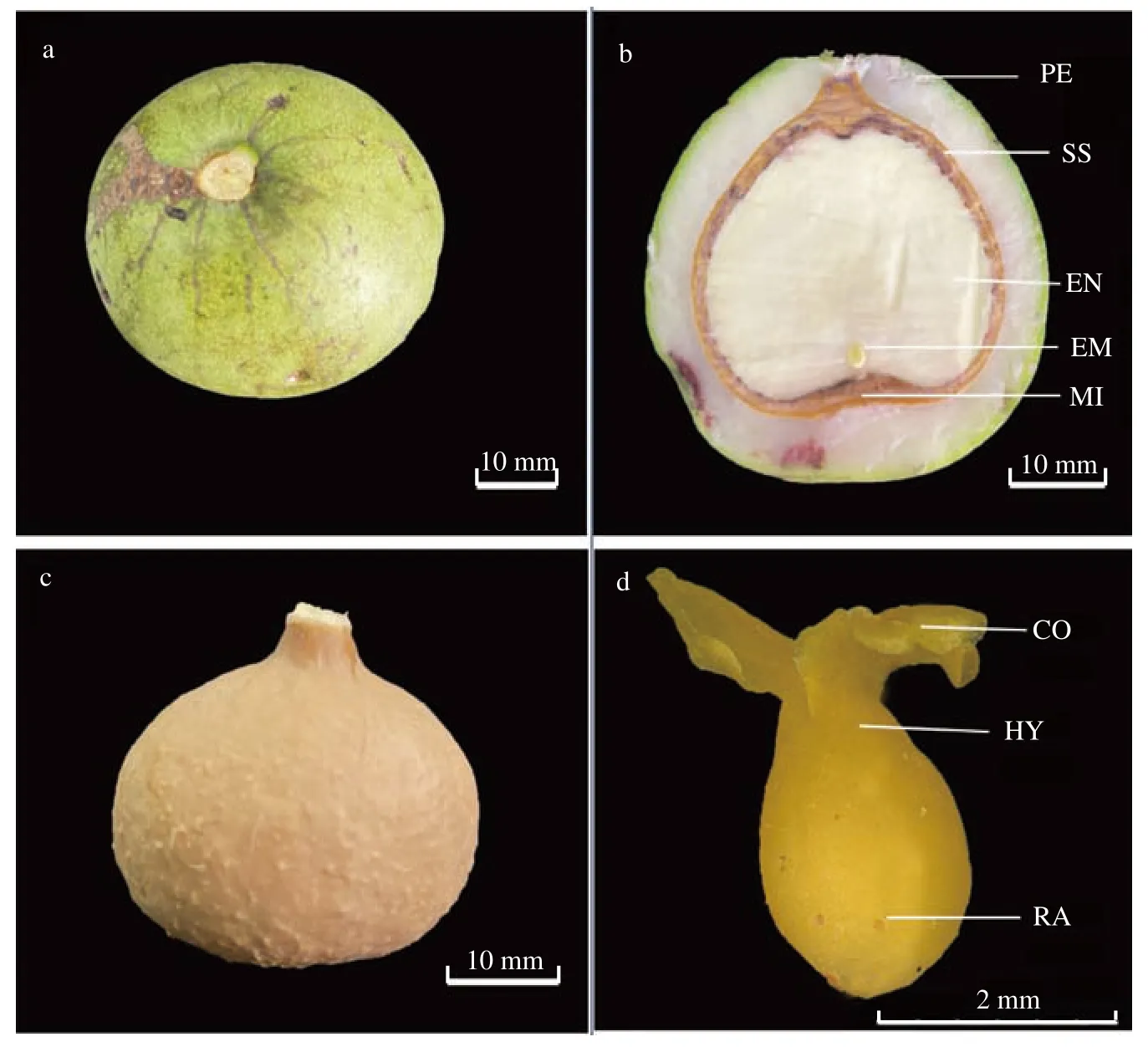
图 1 蒜头果果实与种子形态结构Fig. 1 Morphological structure of fruits and seeds in M. oleifera
果实质量29.72~59.01 g,平均质量39.71 g;果实纵径33.65~46.02 mm,横径38.30~50.16 mm,且变异系数均为0.05。表明蒜头果果实质量、形状和大小变异较大,且横径长略大于纵径,呈球形或扁球形。其中,果皮的厚薄不均,纵向果皮厚3.88~9.24 mm,平均厚5.6 mm;横向厚2.62~6.10 mm,平均厚4.76 mm,纵向果皮略厚于横向果皮;果皮鲜质量23.24 g,占果实鲜质量的58.52%(表1)。
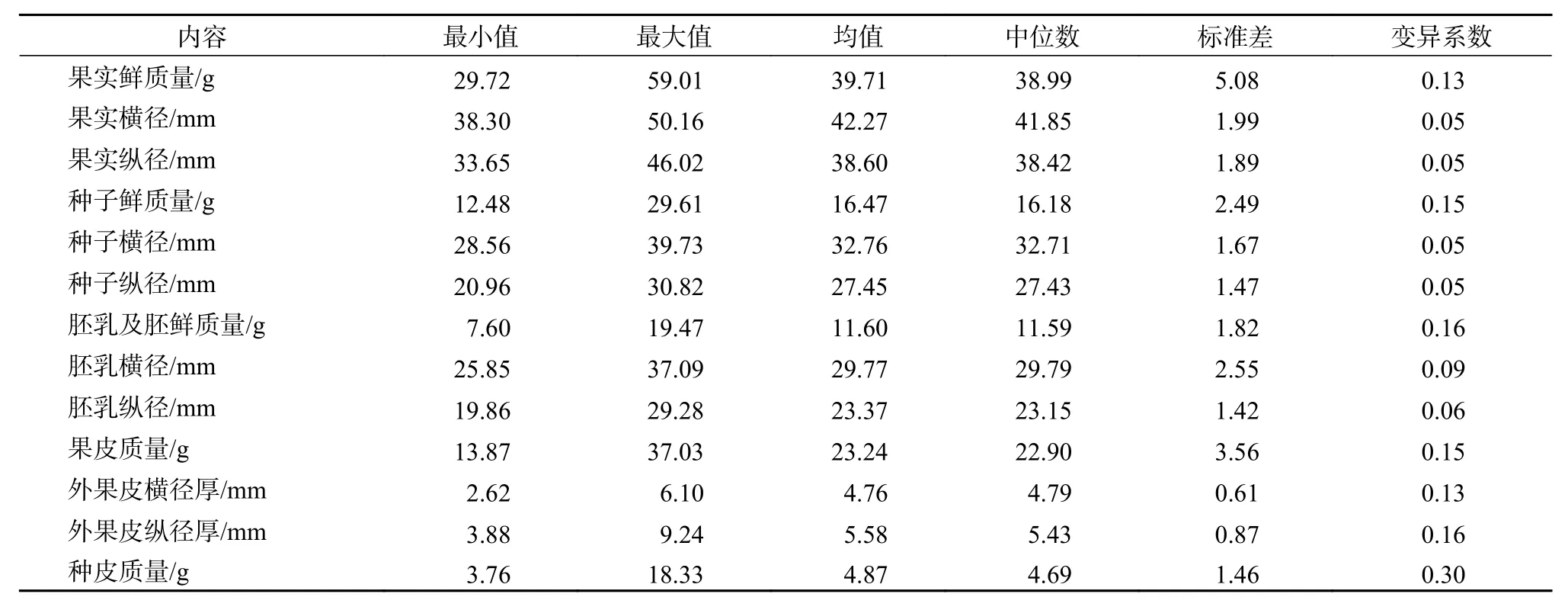
表 1 蒜头果果实结构组成Table 1 Fruit quality indexes of M. oleifera
2.2 蒜头果种子与胚的结构
成熟的蒜头果种子由种壳和种仁(种仁含胚乳和胚)组成,种仁由胚乳和种胚构成。种子长 ×宽为(20.96~30.82)mm × (28.56~39.73)mm。其中胚乳饱满,白色,大小(长 × 宽)为(19.86~29.28)mm × (25.85~37.09)mm;种胚小,包藏于胚乳中,胚奶黄色,棒状体,剥离种壳和胚乳后,种胚长 × 宽为(2.9~6.0)mm × (0.5~0.9)mm,长仅占胚乳纵长的9.77%~19.86%,即种胚仅占胚乳全长的1/5~1/10(图1b、d)。种胚结构观测显示,胚轴短,子叶直立合生并包被胚芽环生于胚轴上端并紧接胚芽的顶端分生组织,下连胚根,子叶和胚芽约占整个胚体体积和解剖结构的1/3,胚根和胚轴约占2/3,形态上已发育形成子叶、胚轴和胚根3 个部分,胚芽仍为一团结构一致的分生细胞,且子叶与胚根、胚轴、胚芽处于同一条线上,但相连接部位界线不清。因此,蒜头果的种子具有丰富的胚乳,但种胚极小。
2.3 果实与种子各组成部分的干物质及其含水率特征
蒜头果种子中的胚乳含有高达51.85%以上的油脂及约25% 的神经酸[2],具有极高的经济价值。因此,其果实和种子各组成部分干物质及含水率的高低是测算经济利用与价值的重要基础数据。蒜头果单粒鲜果质量(39.71 ± 5.08)g(表1),果实的果皮、种壳和胚乳等各组成部分之间的干物质及含水量存在显著差异。其中果皮鲜质量(23.34 ± 3.56)g,具有浓郁的杏仁味,富含苯甲醛;种子鲜质量(16.47 ± 2.49)g。因此,果皮和种子分别占果实鲜质量的58.78% 和41.22%。蒜头果果皮中含水率高达(84.53 ± 0.52)%;果皮和种子干物质质量分别为3.61g 和9.76 g,占果实干物质质量的27.0% 和73.0%。因此,蒜头果果实鲜果和干果的出仁率分别为41.22%和73.0%。
蒜头果种子种壳坚硬木质,呈浅黄色或奶油色,表面有细小稀疏的疣状凸起,质量为(4.81 ±0.42)g,含水率(37.83 ± 1.56)%,约占种子鲜重和干物质量的29.57% 和20.71%。胚乳丰富,呈乳白色,质量约11.60 g,含水率41.42 %,约占种子鲜重和干物质量的70.41% 和79.29%。种胚小,位于胚乳基部(图1b)靠近种孔一侧,通体呈奶油色或浅黄色,重约0.0037 g,含水率高达75.51%,仅占种子鲜重和干物质量的0.022%和0.006%。因此,种子中的胚无论是胚率和胚的质量均极小。蒜头果种子质量大,胚乳丰富,种胚小,在种壳与胚乳之间可见密生细小短绒毛。
蒜头果胚乳中富含神经酸,其干物质重分别约占果实和种子的63.64% 和79.29%,即占种子干物质质量的近五分之四,反映了胚乳是果实和种子结构中重要的组成部分,既是最有潜在经济开发利用价值的主要部分又是提供种胚萌发营养的不可或缺的结构,在蒜头果的种群繁育中具有不可替代的重要作用。
2.4 蒜头果子叶多样性特征
2.4.1 蒜头果子叶的数量与形态
蒜头果虽然是双子叶植物,但其种胚的子叶数量不是恒定的2 枚,而是呈2~5 枚不等。取样统计100 粒果实的子叶显示,具有2、3、4 枚子叶的胚分别占观察数的 1%、56%和43 %,说明多子叶胚在野生蒜头果种子中占绝大多数,且是普遍现象。在体式显微镜下可见,种胚中子叶间相互紧贴不分开,肉质稍肥厚,呈淡黄色或奶油色;在外形上,种胚萌发前子叶因数量2、3、4、5 枚,分别呈“一字型”(2 枚子叶)、“Y 字型”(3 枚子叶)、“X 型”(4 枚子叶)和“口字型”(5 枚子叶)(图2a1、b1、c1),这些特征有别于其他的双子叶植物,也是目前双子叶植物中首次报道的多子叶现象。
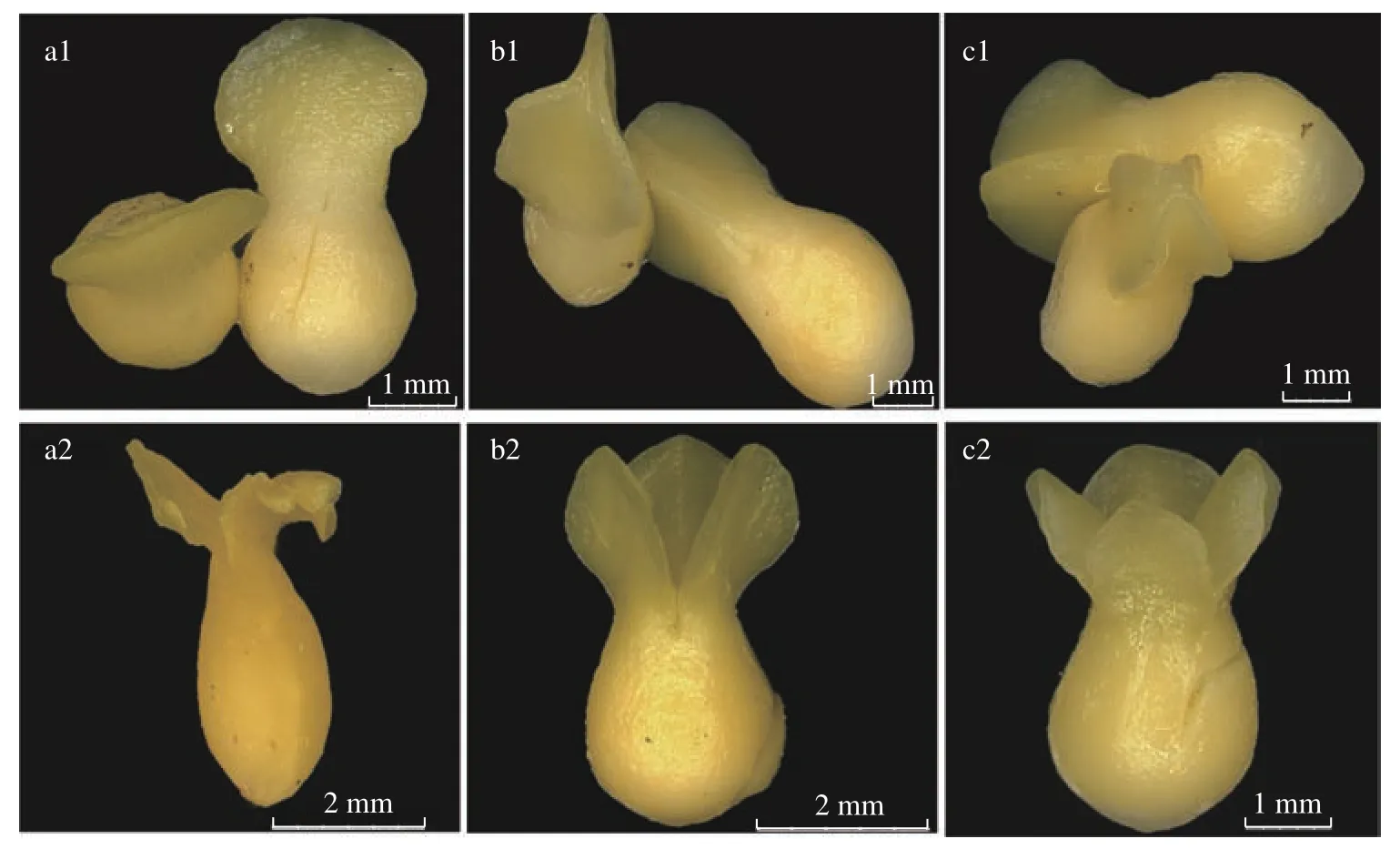
图 2 蒜头果种胚中的子叶数量Fig. 2 Number of cotyledons in seed embryo of M. oleifera
2.4.2 不同海拔分布的野生蒜头果种胚子叶数量的变化
为考察种胚子叶数量随海拔的变化情况,采集蒜头果分布区内最低海拔(400 m)到最高海拔(1 500 m)之间7 个海拔段蒜头果果实,观察子叶数,结果表明蒜头果种胚的子叶数量以3~4 枚占绝大多数,2 者合计占87%以上(图3)。说明蒜头果种胚中多子叶现象可能是其系统发育中形成的,且随蒜头果分布海拔升高,3~4 枚子叶种子的数量呈增加趋势,而2 枚子叶种子的数量减少。作为双子叶植物的蒜头果,其子叶数量以3~4 枚居多,甚至个别出现5 片子叶,是蒜头果种子中的普遍现象,这一现象对研究蒜头果遗传变异的表征和被子植物系统发育具有重要的研究价值和学术意义。
2.5 成熟蒜头果果实中种子及其子叶的解剖特征
通过石蜡包埋切片观察显示,果实成熟时胚长仅2.9~6.0 mm,包含胚根、胚轴、胚芽和子叶4 部分,处于同一条线上,但连接部位界线不清(图4a)。子叶约占整个种胚长度的50%,而胚芽、胚根和胚轴合占约50%,整个胚未见导管等维管组织的分化,呈幼嫩含水量高的肉质结构。子叶为幼嫩含水量高的肉质片状结构,因子叶数量不同,横切面呈不同的形状,如4 枚子叶为X 型,子叶间上表皮相互间紧贴无间隙(图4b);子叶上下表皮及其之间的细胞均排列紧密,由结构一致的核大、质浓的分生细胞组成,仅见维管束原基,且无叶绿体,未见角质层和蜡被等附属物、未见气孔器(图4c)。胚轴极短,胚芽由一团还未分化的分生细胞组成,细胞核大,细胞质浓,细胞近方形,未见叶原基、腋芽原基和维管柱的分化(图4a、d),组成胚芽的分生细胞被环生于胚轴上端的子叶包被。胚芽下连胚轴但未见明显的分界线,胚轴下接胚根,胚根的顶端分生组织与根冠的界线明显,已分化维管束原基(图4a、f)。从胚根的基本分生组织、根冠及其初见的维管束原基可以看出,根的表皮、皮层及维管束原基均来自共同的分生细胞,且分生细胞未见分层现象(图4f)。因此,蒜头果胚根的分生细胞通过分裂,不断产生一些细胞加入到根中成为新的根部的细胞,同时这些不断产生细胞的分生细胞仍然保留在分生组织中,这些经过不断更新始终保留在分生组织中具有分生能力的细胞,是产生根的其他所有细胞的原始细胞。
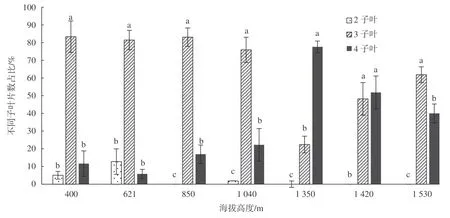
图 3 蒜头果不同海拔不同子叶枚数占比Fig. 3 Percentage of the number of cotyledons of M. oleiferaat different altitudes
成熟蒜头果果实中胚的解剖结构显示,种胚长度仅占胚乳长度的1/5~1/10,虽然种胚已有胚根、胚轴和子叶的分化,但胚芽和胚轴均是由细胞核大、细胞质浓的分生细胞组成,未见维管束原基、叶原基和腋芽原基的分化,仅子叶和胚根中可见维管束原基的分化,说明此时种胚还未达到生理和形态成熟。成熟果实离开母体后,其种子尚需要时间完成生理和形态后熟。种子须湿砂层积后,种胚才能完成组织的分化及维管束的形成,进而从环境中吸收水分和营养完成萌发。
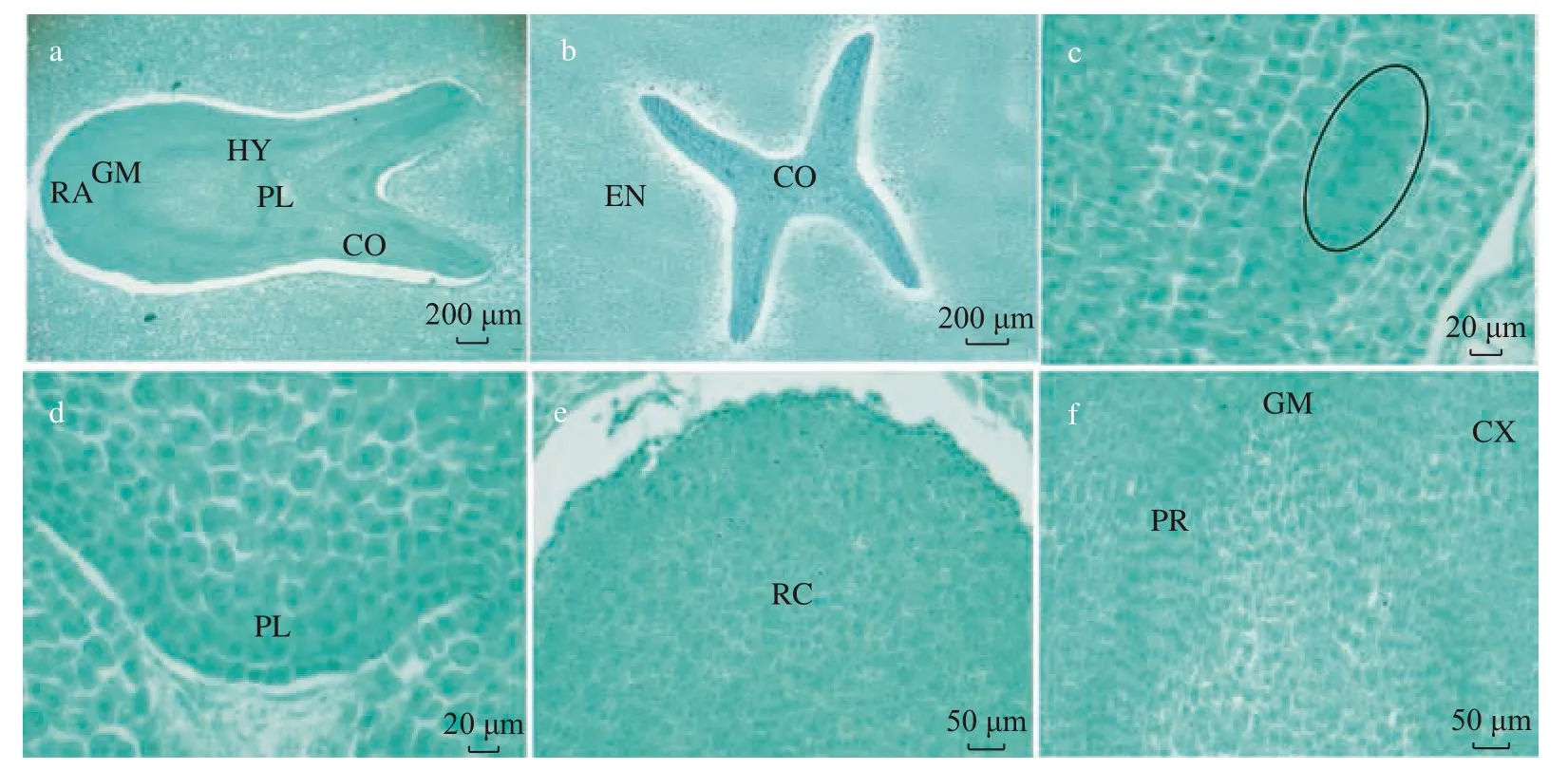
图 4 蒜头果种胚解剖结构Fig. 4 Anatomical structure of M. oleiferaembryos
a. 胚的纵切面, 示胚被胚乳包围的格局和子叶、胚芽、胚轴和胚根; b. 叶中部横切面,示4 枚子叶被胚乳包围的格局; c. 图 a 中子叶的局部放大图, 示叶脉; d. 图 a 中胚芽的局部放大图, 示胚芽的基本分生组织; e. 图 a 中胚根的局部放大图, 示根冠; f. 图 a 中胚根的局部放大图, 示基本分生组织、原形成层和皮层,图中圆圈代表维管束。GM 为基本分生组织; PR 为原形成层; CX 为皮层; PL 为胚芽; RC 为根冠。
3 结论与讨论
3.1 蒜头果果实与种子的结构
蒜头果属有胚乳种子。蒜头果花期为3~5 月,5 月下旬出现幼果,10 月果实成熟。成熟果实由果皮和种子2 部分构成,分别占果实鲜质量的58.52%和41.47%,占干物质质量的27.0%和73.0%,说明果皮含水量高达73%。
蒜头果种子由种壳、胚乳和种胚3 个部分构成,种子鲜重12.48~29.61 g。种胚极小,位于胚乳基部近种孔一端,长仅2.9~6.0 mm,占胚乳长的 1/5 ~1/10;胚和胚乳分别占种子干物质量的0.006%和79.29%。因此,无论是胚率和胚的干物质质量均极小,丰富的胚乳为蒜头果种子的萌发提供了营养。胚极小,是蒜头果原始性特征的体现。
3.2 成熟果实的种子因种胚未完成细胞分化而成为生理和形态后熟种子
蒜头果种子种胚有胚根、胚轴、胚芽和子叶4 个部分,但胚芽和胚轴均由分生细胞组成,未见维管束原基、叶原基和腋芽原基的分化,意味着此时种胚还未达到生理和形态成熟。因此,蒜头果成熟果实离开母体后,其种子还要经历生理后熟后才能萌发,这与已有的研究结果认为蒜头果种子的胚需要经历约75 d 的生理和形态后熟才能萌发的结论一致[6]。因此蒜头果种子的生理后熟是由于种胚还未完成细胞分化和组织形成所致,种子砂藏后才能完成生理和形态后熟进而萌发。结合蒜头果的果实特征,推测在自然条件下,蒜头果果实成熟后,在其含水量高达58.52%果皮的包裹保护下,种胚才能完成后熟过程是其对生境的适应特征。
3.3 种胚多子叶现象的发现
蒜头果种胚的子叶数量以3~4 枚居多,是蒜头果种子中的普遍现象。对不同海拔的种子统计,说明这种现象不是自然变异或基因突变产生,推测是系统发育中形成的,可能与蒜头果处被子植物基部较原始的位置及其古老特性有关。该发现为研究种子植物系统演化进程中种胚子叶数量演化的规律提供了新案例,也反映了蒜头果在双子叶植物系统演化中所处的古老地位。
子叶是植物的重要器官,是植物进化的一个重要标志,是植物变异最小的性状之一[8]。因此,子叶数目是种子植物划分为被子植物(包括双子叶植物和单子叶植物)和裸子植物(多子叶植物)两大类群的依据之一。被子植物更进化,认为单子叶是由双子叶的1 个子叶退化形成。有研究认为双子叶植物中的多子叶现象属于自然突变,也可人工诱变产生[15-18],但出现几率极小。自然突变的如草本植物的四季豆(Phaseolus vulgaris)中发现2~7 片多子叶幼苗[19];野萝卜(Raphanus raphanistrum)种子中约占4.8% 萌发的植株子叶数量少于或多于2 片[8];甘蓝型油菜(Brassica napus)的三子叶幼苗[20],茄(Solanum melongena) 和红茄(S.Aethiopicum)[21],仙客来(Cyclamen persicum)[22],向日葵(Helianthus annuus)品种‘种富阳’幼苗中也有单子叶和三子叶 2 种形态,但所占比例仅分别为2.3%和0.9%[23]。此外,木本双子叶植物中的枫树(Acerspp.)也发现有三子叶和四子叶的幼苗[24];桑树(Morus alba)的20 个杂交组合获得的种子中也发现3 枚和4 枚子叶的幼苗,但出现概率仅为0.33~4.5 株/g[25]。可见,已有报道无论是草本还是木本的双子叶植物中自然突变产生的多子叶现象所占比例都非常低,不超过5%,属零星发生。而蒜头果的种子中具有3~4 枚子叶的种子数却占种子数的87% 以上,并随蒜头果分布海拔的升高,3 枚和4 枚子叶的种子数呈增加趋势,甚至零星还可见5 枚子叶的种子。因此,多子叶现象在蒜头果种子中具有普遍性,这与双子叶植物类群中已有报道的多子叶现象不同。蒜头果作为被子植物基部原始花被亚纲檀香目铁青树科蒜头果属的单种属植物,具有如此高比例的多子叶的种子存在,故推测不是自然变异或基因突变引起,而是在其系统发育中形成的。
结合已有的研究认为部分植物的三子叶是从双子叶演变而成,如裸子植物的松柏类的多子叶现象衍生于双子叶的分裂[26];也有研究认为双子叶植物中一些种子会有极少比例出现的三子叶现象是一种“返祖现象”[23];对桑树种子的多子叶突变的研究也推测属于“返祖现象”[25]。结合蒜头果种胚的多子叶特征,与已有报道的裸子植物中的南洋杉科南洋杉属的植物有相似性,南洋杉属中的Eutacta 组植物具有4 枚分离的子叶,而其他种均为2 枚子叶[27]。推测蒜头果可能是被子植物演化环节中的一个重要物种,但需要通过胚胎学对其多子叶形成进行研究,为深入了解单子叶植物和双子叶植物的进化过程提供基础数据和参考。蒜头果是铁青树科蒜头果属的单种属植物,科中还有26 属多达200 余种植物[28]未见多子叶现象的报道。因此,蒜头果种胚中多子叶的形成机制、途径与其他近缘种关系的研究,能为进一步认识蒜头果的系统分类地位及其演化关系提供重要信息和证据。
致谢: 感谢赵瑞同学在富宁县和广南县不同海拔区域采集的蒜头果果实,为种胚子叶数量随海拔的变化提供了观测材料。
