猪CD163 基因的克隆及截短原核表达
2023-06-15刘雪晴张可唐青海薛姣雄隆泽桧高翠翠刘之睿苏荣唐斯萍
刘雪晴,张可,唐青海,薛姣雄,隆泽桧,高翠翠,刘之睿,苏荣,唐斯萍
(衡阳师范学院生命科学学院,南岳山区生物资源保护与利用湖南省重点实验室,动物微生物新型检测技术及生物制剂衡阳市重点实验室,湖南 衡阳 421008)
CD163 是一种富含B 族半胱氨酸的清道夫受体,是富半胱氨酸清道夫受体(scavenger receptor cysteine-rich domain,SRCR)超家族的重要成员之一[1],表达于外周血单核细胞和大多数组织巨噬细胞中[2]。CD163 为典型的Ⅰ型糖基化的跨膜蛋白,因富含半胱氨酸,内部可以形成9 个由二硫键连接构成的结构域[3]。CD163 在多种存在成熟巨噬细胞的组织中均高度表达。
研究表明,猪CD163(pCD163)是猪繁殖与呼吸综合征病毒(Porcinere productive and respiratory syndrome Virus , PRRSV) 感染宿主细胞的关键受体[4]。PRRSV 是一种单链RNA 病毒,包括欧洲型(PRRSV-1 型)和美洲型(PRRSV-2 型)两种基因型[5],在我国主要流行的是美洲型[6],主要引起母猪的流产和仔猪的呼吸障碍[7]。在PRRSV 感染过程中,pCD163 的SRCR 结构域与PRRSV 的糖基化膜蛋白GP2 和GP4 相互作用、促进病毒脱壳[8]。目前防控PRRSV的主要方式是疫苗免疫[9],但由于病毒存在免疫逃逸机制和抗体依赖性增强效应、现有疫苗免疫的效果并不十分理想[10],至今仍无有效的治疗措施[11]。Whitworth 等人利用基因编辑技术将pCD163 基因剔除,pCD163 基因缺失猪在整个研究期间未显示任何临床症状,不被现有的两种基因型PRRSV 的野毒株感染[12]。Wang 等通过删除猪的CD163 外显子7,赋予了该基因编辑猪对高致病性蓝耳病病毒(Highly pathogenic-Porcinere productive and respiratory syndrome Virus , HP-PRRSV)感染的抗性[13]。这些研究都证实了CD163 是PRRSV 感染宿主细胞的核心受体。
CD163 在未来药物的开发方面具有巨大的潜力,是目前已知的最佳药物开发靶点。本研究克隆猪CD163 基因,并进行体外截短表达,为进一步开展受体药物的研究奠定基础。
1 材料与方法
1.1 材料
1.1.1 细胞、质粒、菌株及实验动物
三元杂交猪的母猪与哺乳仔猪抗凝血全血由衡阳某养猪公司提供;感受态细胞DH5α、Transtta(DE3) 购自北京全式金生物技术股份有限公司;pMD18-T 载体购自宝生物工程(北京)有限公司;原核表达载体pGEX-4T-1 购自Pharmacia Biotech公司。
1.1.2 主要试剂、材料
RNA iso、PrimeScript II First Strand cDNA Synthesis Kit、DNA Ligationg Mix、GXL polymerase高保真酶、内切酶及IPTG 均购自宝生物工程(北京)有限公司;质粒DNA 提取试剂盒购自Axyprep 公司。Mouse anti-GST 单克隆抗体、HRP-羊抗鼠二抗均购自武汉三鹰生物技术有限公司;DAB 显色试剂盒购自北京中杉金桥生物技术有限公司。
1.2 方法
1.2.1 pCD163 基因的克隆、序列分析
采用TriZol 试剂提取三元杂交猪的抗凝血总RNA,取2 μg 总RNA,采用PrimeScript II First Strand cDNA Synthesis Kit 合成第一链cDNA,取2 μL 产物分别采用引物pCD163-orf-F1/R1 749、pCD163-orf-F1 717/R3348、pCD163-orf-F1/R3 348(根据pCD163 基因序列(GenBank No:OM321546 设计)进行PCR 扩增),扩增得到全长pCD163 基因及其两个截短基因片段,PCR 产物经纯化后克隆至pMD18-T 载体,转化DH5ɑ 感受态细胞进行抗性筛选,重组质粒进行酶切鉴定和序列测定。通过GenBank 中的BLAST 网址(http://www.ncbi.nlm.nih.gov/ blast)和DNASTAR MegAlign 软件包进行核酸序列比对分析,用MEGA 5.0 进行进化分析,引物序列见表1。

表1 引物序列及其反应条件Table 1 Oligonucleotide sequences of PCR primer sets and reaction conditions.
1.2.2 重组原核表达载体的构建及鉴定
以pMD18-T-pCD163 载体为模板,参照GXL polymerase 高保真酶说明书,用表达引物pCD163-F1-BamH I/pCD163-R1749-Xho I、pCD163-F1717-BamH I/pCD163-R3348-Xho I 扩增pCD163 基因的截短片段,PCR 产物经BamH I 和Xho I 双酶切后与经同种酶双酶切的pGEX-4T-1 连接、转化Transetta(DE3)感受态细胞,阳性克隆采用双酶切鉴定和序列测定分析。
1.2.3 重组pCD163 蛋白的诱导表达及鉴定
将双酶切和测序结果正确的阳性菌株接种至新鲜LB 培养基中进行活化,扩大培养2.5 h 后,加入终浓度为1 mmol·L-1的IPTG,于37 ℃,220 r·min-1条件下诱导目的蛋白表达,同时设置未加IPTG 诱导剂组作为对照,具体操作参照Amersham Pharmacia Biotech pGEX-4T-1 操作手册进行。菌体采用超声破碎后离心,分别收集上清和沉淀。蛋白样品经SDS-PAGE 电泳后,转印到PVDF膜上,用2% 脱脂奶于37 ℃封闭1 h,PBST 漂洗3次后与一抗GST Tag-Mouse 单克隆抗体(1 :3 000稀释)孵育1 h,PBST 洗3 次,与二抗HRP-羊抗鼠(1 :3 000 稀释)孵育1 h,PBST 洗涤后,用DAB 显色试剂盒显色。
2 结果
2.1 猪pCD163 基因的克隆与序列分析
2.1.1 pCD163 基因的克隆
琼脂糖凝胶电泳结果显示,在1500 bp~2000 bp 之间分别有特异性条带,在约3000 bp 处有一条特异性条带,与预期结果相符(图1 泳道1-3)。阳性克隆菌DH5α-pMD-18T-pCD163-1-1749、DH5αpMD-18T-pCD163-1717-3348、DH5α-pMD-18TpCD163-1-3348 经BamH I 和Hind Ⅲ双酶切,电泳结果显示,均呈阳性(如图1 泳道4~6),表明目的基因已成功克隆到pMD-18T 载体中。
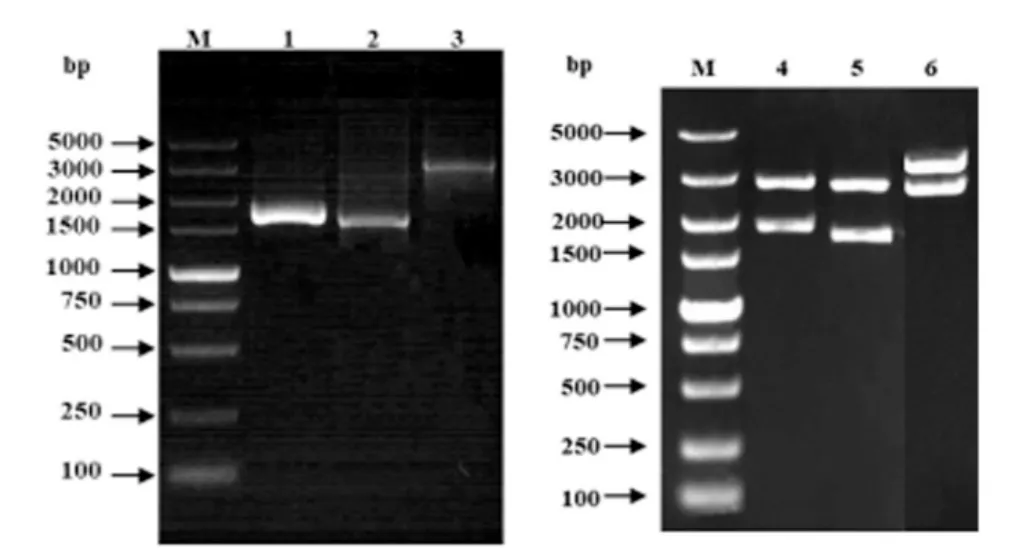
图1 pCD163 基因的PCR 扩增与DH5α-pMD-18T-pCD163-1-1749、DH5α-pMD-18T-pCD163-1717-3348、DH5α-pMD-18T-pCD163-1-3348 阳性克隆菌的筛选与酶切鉴定Fig1 Amplification of pCD163 gene by PCR and enzyme digestion identification of DH5α -PMD-18T-pCD163-1-1749、DH5α -PMD-18T-pCD163-1717-3348、DH5α-PMD-18T-pCD163-1-3348 positive clones
2.1.2 pCD163 基因的序列分析
序列分析表明,pCD163 基因开放阅读框(ORF)全长为3348 bp,编码1115 个氨基酸(aa),蛋白分子量大小约为120.5 kDa。将该序列在NCBI 数据库提交后获得GenBank 登录号:OM321546。
目前NCBI 数据库的GenBank 中已经测定的CD163 基因序列有8 个物种:人、绿猴、野猪、牛、家鼠、褐家鼠、家鸡、狼,其他动物的CD163 基因mRNA 序列均为预测序列。该基因与GenBank 中35个物种(详细信息见图2)的核苷酸相似性在54.7%~91.1%之间,与抹香鲸(physeter catodon) CD163(XM_024129244)的相似性最高(91.1%),与人(Homo sapiens) CD163(DQ058615)的相似性为87.8%,与针尾雨燕(Chaetura pelagica)CD163(XM_009998806)的相似性最低(54.7%)。利用Mega5.0 软件的邻结法(Neighbor Joining)对所测定的CD163基因的序列与35 个物种的核苷酸序列进行进化分析,结果显示,猪CD163 与抹香鲸 (Physeter catodon CD163(XM_024129244))和牛(Bos taurus CD163(GQ184570))的CD163 在同一分支上,表明它们亲缘关系最近;猪CD163 与人(Homo sapiens CD163(DQ058615))和绿猴(Chlorocebus aethiops CD163(JF753553))等灵长类动物的亲缘关系也相对较近,而与几维鸟(Apteryx rowi CD163(XM_026080316))、针尾雨燕(Chaetura pelagica CD163(XM_009998806))、阿德利企鹅(Pygoscelis adeliae CD163(XM_009329868))和鹃(Leptosomus discolor CD163(XM_009947018))等鸟纲动物的亲缘关系相对最远,鸟纲独自分布在另一支中(图3)。

图2 猪CD163 基因核酸序列与其他物种CD163 基因核酸序列相似性比较Fig 2 Nucleic acid homologous comparison of porcine CD163 with that of other species

图3 各个物种CD163 基因进化树Fig3 Each speciesCD163 gene evolutionary tree
2.2 pCD163 截短基因重组表达载体的鉴定
重组子Transetta(DE3)-pGEX-4T-1-CD163-1-1749 和重组子Transetta(DE3)-pGEX-4T-1-CD163-1717-3348 经BamH I 和Xho I 双酶切,进行1.2%琼脂糖凝胶电泳,分别得到片段为1750 bp 的目的条带、1632 bp 的目的条带和4900 bp 的载体片段,表明目的基因已经成功克隆到pGEX-4T-1 载体中(图4)。
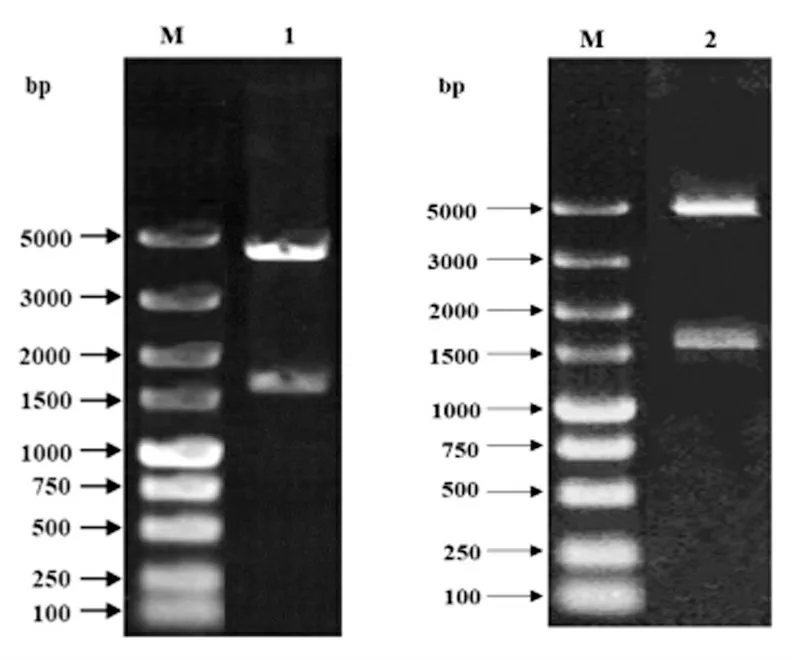
图4 重组表达载体Transetta (DE3)-pGEX-4T-1-pCD163-1-1749 和Transetta (DE3)-pGEX-4T-1-pCD163-1717-3348的酶切鉴定Fig4 Identification of recombinant plasmid Transetta (DE3)-pGEX-4T-1-pCD163-1-1749 and Transetta (DE3)-pGEX-4T-1-pCD163-1717-3348 by enzyme digestion
2.3 重组pCD163 蛋白的诱导表达与鉴定
阳性菌株Transetta(DE3)-pGEX-4T-1-pCD 163-1-1749 经诱导,与未诱导的样品(泳道3、4)对比分析,可看出在泳道2 的70 kDa~100 kDa 有一条目的条带(图5-A),与预期大小相符。
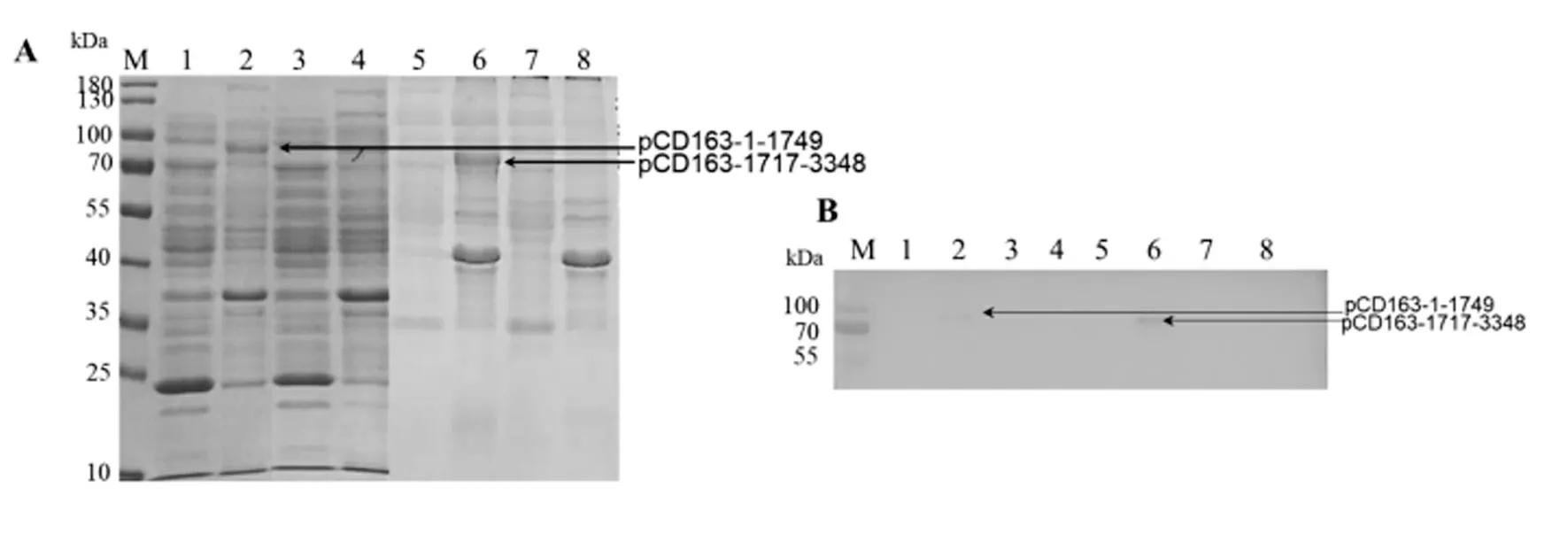
图5 重组蛋白pCD163 的SDS-PAGE 检测和Western blot 鉴定(GST 单克隆抗体)Fig5 Detection of recombinant protein pCD163 by SDS-PAGE and Western blot (GST mAb)
阳性菌株Transetta(DE3)-pGEX-4T-1-pCD163-1717-3348 经诱导,与未诱导的样品(泳道7、8)对比分析,可看出在泳道6 的70 kDa~100 kDa 有一条目的条带(图5-A),与预期大小相符。
将重组pCD163 蛋白进行Western blot 检测,结果显示,pGEX-4T-1-pCD163-1-1749 包涵体蛋白和pGEX-4T-1-pCD163-1717-3348 包涵体蛋白分别在92 kDa 处有一条目的条带、85 kDa 处有一条目的条带(图5-B),说明目的蛋白成功表达。
3 讨论
本研究所测定的猪pCD163 基因与预测的马来穿山甲的CD163(GenBank No. XM017667238)核苷酸相似性最高(88.7%)。结合进化树图(图3),牛与抹香鲸在同一分支上,CD163 亲缘关系最近,提示我们这两个物种对PRRSV 的病毒易感性可能更强;一般来说,动物分类学上属于同目或同科的物种其CD163 基因在进化树上也处于同一分支;但也有例外,牛和抹香鲸的CD163 基因进化树中处于同一分支,但牛属于偶蹄目,而抹香鲸属于鲸目,土豚和牛均属于偶蹄目,土豚却在另一分支中,所以土豚和牛的CD163 亲缘关系相对较远;此外,鸟纲动物单独在另一分支中,与猪的CD163 亲缘关系相对较远,Dong 等在PRRSV 感染试验中发现CD163 基因中的所有单核苷酸多态性(Single nucleotide polymorphism , SNP) 与所评估的遗传背景具有显著的相互作用效应[14];所以相对来说,鸟纲动物应当不易感染此种病毒。Li 等将猪的CD163全长基因分成三段(160~798 bp、790~2046bp、2143~3 084 bp),并去掉信号肽和跨膜区,与原核表达载体pET-28a(+)重组进行诱导表达,其重组蛋白均以包涵体形式表达[15],其三个片段Y-P1 (160~798 bp)、Y-P2 (790~2046 bp)和Y-P3 (2143~3084 bp),分子量较小,相对于本研究的较大分子量片段更易于表达,并且其病毒阻断试验表明,Y-P2 蛋白和抗Y-P2 小鼠血清均能阻断PRRSV 感染,且呈剂量依赖性,Y-P3 蛋白能改善病毒感染,而其试验的未去除信号肽和跨膜区的CD163 片段P1 (1~798 bp)、P2 (790~2046 bp)、P3 (2023~3345 bp)的细胞对PRRSV 不敏感,本研究将CD163 全长基因截短,同样未去除信号肽和跨膜区,结果蛋白表达量较少,原核细胞没有核膜,缺少细胞器,多余的信号肽不能被细胞识别,信号肽有可能影响活性蛋白质的正确折叠;这表明去掉信号肽和跨膜区能够更好地表达CD163 蛋白。李秀媛等选择了亲水性和抗原指数较高的第45~297 位氨基酸编码区进行原核表达试验,去掉信号肽和跨膜区,与含CTLA-4 IgV编码序列的表达载体pQE-IgV 连接,融合在重组大肠杆菌中获得了高效表达,但表达产物也几乎全部以包涵体形式存在[16],而本研究通过将猪CD163截短成两段(1~1749 bp 及1717~3348 bp),与原核表达载体pGEX-4T-1 重组进行诱导表达并在92 kDa 和85 kDa 处可见包涵体形式的目的蛋白,由此可见,CD163 各截短片段在原核表达系统中虽载体等不尽相同,但其表达形式均为包涵体形式,这提示我们猪的CD163 蛋白特性使其更易于以包涵体形式表达。本研究Western blot(图5-B)结果还表明,pCD163 蛋白包涵体形式与GST-mAb 呈现良好的反应活性。张留君等将CD163 受体SRCR5 结构域蛋白进行表达、纯化,并免疫大白兔制备其特异性多克隆抗体IgG,当CD163 受体SRCR5 结构域被其特异性抗体封闭后,能够显著抑制体外PRRSV 感染PAMs 细胞的抗体依赖性增强作用(Antibody dependent enhancement , ADE)[17]。该研究的重组蛋白也均以包涵体形式表达,通过使用尿素法纯化后获得了高纯度的目的蛋白,这提示我们猪的CD163 蛋白特性虽可能使其更易于以包涵体形式表达,但在纯化后仍然可以制备高滴度的抗体。段盈伊等制备了CD163 单克隆抗体用于检测巨噬细胞极化表型,效果良好[18]。Xu 等观察到CD163-3D4/21 细胞能够支持所有测试的PRRSV2菌株的感染,包括中国PRRSV 株的流行谱系NADC30 样PRRSV[19]。这说明,在中国流行的毒株也能通过CD163 作为靶点开发有效药物。各研究证明CD163+巨噬细胞的特异性靶向已被证明是一种很有前途的治疗炎症和恶性疾病的药物传递策略,有助于目前的药物治疗[20]。Wu 等发现pCD163-MARC 细胞具有较高的PRRSV 临床样本的分离率,并且更有效地繁殖PRRS 减毒活疫苗株[21]。在头颈部鳞状细胞癌(Squamous cell carcinoma of head and neck , SCCHN) 中评估CD163+TAMs 具有未来临床应用的潜力[22]。Stoian 等提出一个潜在的疫苗靶点是诱导抗体,破坏巨噬细胞CD163 受体与病毒粒子表面突出的GP2、GP3 和GP4 异源三聚体之间的相互作用[23]。Etzerodt 等利用遗传和药理学工具选择性耗竭大网膜中的CD163+Tim4+巨噬细胞可防止肿瘤进展和疾病转移扩散[24]。CD163 在巨噬细胞中的非脱落膜形式构成了药物靶向炎症巨噬细胞的靶点,该方法已用于动物炎症模型,以显著提高抗炎糖皮质激素药物的表观治疗指数,该药物与抗CD163 抗体偶联[25]。这些研究表明,CD163 不仅在抗病毒方面有较大潜力、在其他疾病的治疗中也有一定的应用前景。
综上所述,本研究成功克隆了pCD163 基因及其截短基因片段,并表达其截短基因片段的蛋白,为后续基因功能研究、抗体制备及药物开发研究奠定了基础。□
