正交法优选调控秦艽次生代谢产物含量变化的条件
2023-05-22付欢欢何懿菡尹洋洋胡伟杨烨岳正刚
付欢欢 何懿菡 尹洋洋 胡伟 杨烨 岳正刚
摘 要:為优选可抑制秦艽次生代谢产物含量变化的条件,该文采用3因素4水平正交试验设计方法,共设计16组处理,研究洛伐他汀(MVA途径抑制剂)、膦胺霉素(MEP途径抑制剂)和取样天数对秦艽中马钱苷酸、獐牙菜苷、獐牙菜苦苷、龙胆苦苷4种主要环烯醚萜类化合物含量的影响。结果表明:(1)4种环烯醚萜类化合物含量变化受取样天数影响最大,其次为膦胺霉素浓度,次之为洛伐他汀浓度。(2)以最佳抑制条件处理后,马钱苷酸、獐芽菜苦苷、龙胆苦苷和獐芽菜苷含量分别下降了69%、36%、33%和4%。基于正交法优选的抑制条件,对4种化合物均可抑制。综上所述,可确定调控秦艽次生代谢产物含量变化的最佳抑制条件为膦胺霉素400 μmol·L-1,洛伐他汀50 μmol·L-1,取样天数6 d,该条件为进一步研究MEP和MVA途径在环烯醚萜类化合物代谢合成中的调控机制奠定基础。
关键词: 正交试验, MVA途径, MEP途径, 洛伐他汀, 膦胺霉素, 环烯醚萜
中图分类号:Q946
文献标识码:A
文章编号:1000-3142(2023)04-0777-08
Abstract:Gentiana macrophylla is a traditional Chinese herbal medicine for treating jaundice, hepatitis, and stomachic and choleretic ailments, and its main secondary metabolites are secoiridiod glycosides represented by gentiopicroside. The biosynthetic pathway of gentiopicroside is via methylerythritol phosphate pathway (MEP) and mevalonate pathway (MVA) pathways. Fosmidomycin and lovastatin are specific inhibitors of 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR) of the MEP pathway and hydroxymethylglutaryl-CoA reductase (HMGR) of the MVA pathway, and are widely used in the research of the biosynthesis pathway and regulation mechanism of secondary metabolites. The article aims to test and search for the optimal condition for decreasing the contents of four secoiridiod glycosides compounds (loganic acid, sweroside, swertiamarin and gentiopicroside) in G. macrophylla by using the orthogonal test designs. In this study, the concentration of fosmidomycin(A), concentration of lovastatin(B) and sampling days (C) were taken as three factors, and each factor was taken as four levels to design an orthogonal test, with 16 groups of treatment. HPLC was used to determine the contents of four secoiridiod glycosides, and the data were statistically analyzed. The results were as follows: (1) The contents of the four secoiridiod glycosides compounds in G. macrophylla were most affected by sampling time, followed by fosmidomycin concentration and then lovastatin concentration. (2) After treatment with the optimal inhibitory conditions, the contents of loganic acid, swertiamarin, gentiopicroside and sweroside decreased by 69%, 36%, 33% and 4% respectively. In conclusion, optimal inhibition conditions are determined to be fosmidomycin 400 μmol·L-1, lovastatin 50 μmol·L-1, samples are harvested on six days, which provides the reference for further research on the regulatory mechanism of MEP and MVA pathways in the metabolism and synthesis of secoiridiod glycosides.
Key words: orthogonal test, MVA pathway, MEP pathway, lovastatin, fosmidomycin, secoiridiod glycosides
秦艽是龙胆科(Gentianaceae)龙胆属(Gentiana)多年生草本植物(彭美晨和艾晓辉,2021),是国家传统中药,味辛、苦,性平,归胃、肝、胆经(杨飞霞,2020)。秦艽以根入药,有祛风湿、清湿热的作用,可以止痹痛、退虚热,用于风湿痹痛、筋脉拘挛、湿热黄疸、小儿疳积发热等(中国药典2020年版)。秦艽主要含有环烯醚萜类成分,如龙胆苦苷(gentiopicroside)、獐牙菜苦苷(swertiamarin)、獐牙菜苷(sweroside)、马钱苷酸(loganic acid)等(杨飞霞,2020)。环烯醚萜类化合物有广泛的生物活性,如抗肿瘤和抗炎等(Tundis et al.,2008),龙胆苦苷和獐芽菜苦苷有良好的抗炎镇痛和保肝作用(吴昕怡和刘小莉,2017;李跟旺和王磊,2018;Muhamad Fadzil et al.,2021)。
萜类化合物的生源合成途径主要由位于细胞质中的甲羟戊酸途径(mevalonate pathway,MVA途径)和质体中的2-甲基赤藓糖醇-4-磷酸途径(methylerythritol phosphate pathway,MEP途径)组成(Hua et al.,2014;Liao et al.,2016)。结合长春花单萜吲哚生物碱合成途径的研究(Miettinen et al.,2014)以及秦艽转录组数据的挖掘(Cao et al.,2016),推测龙胆苦苷的生源合成途径(康恒等,2021)主要通过MVA和MEP途径分别合成焦磷酸异戊烯酯(isopentenyl diphosphate,IPP),IPP可异构化形成其同分异构体焦磷酸二甲基丙烯酯(dimethylallyl diphosphate,DMAPP),并一起缩合成焦磷酸香叶酯(geranyl pyrophosphate,GPP),GPP再经过多步酶促反应依次合成马钱苷酸、马钱苷、裂环马钱苷以及龙胆苦苷,并衍生出獐牙菜苦苷和獐牙菜苷等萜类化合物,而中间的反应步骤尚不清楚(Liao et al.,2016; Li et al., 2022)。
目前,有关MVA和MEP途径在代谢产物合成中作用机制备受关注。膦胺霉素和洛伐他汀分别为MEP途径1-脱氧-D-木酮糖-5-磷酸还原异构酶(1-deoxy-D-xylulose-5-phosphate reductoisomerase,DXR)和MVA途径HMG-CoA还原酶(hydroxyme-thylglutaryl-CoA reductase,HMGR)的专一抑制剂(魏洁书,2013),被广泛应用于次生代谢产物的生源合成途径及调控机制的研究。为了解穿心莲内酯的生物合成途径,Sinha等(2018)在穿心莲的悬浮细胞中添加洛伐他汀和膦胺霉素,结果发现MEP和MVA通路受多种水平调控,DXR和HMGR对穿心莲内酯的贡献率分别为76.6%和77.6%,这两种通路对穿心莲内酯的产生都有重要贡献。刘丽(2012)在人参发根中添加洛伐他汀和膦胺霉素,通过检测其次生代谢产物含量推断人参总皂苷生物合成的主要通路,结果发现洛伐他汀和膦胺霉素均可抑制人参总皂苷生物合成,并推测MVA途径在人参皂苷的生物合成中占主要作用。Rather等(2019)在青脆枝的发根组织中添加膦胺霉素和洛伐他汀,通过评估次生代谢产物含量以及生物合成通路的主要基因的变化,研究MVA和MEP途径的相互作用,发现膦胺霉素和洛伐他汀可显著降低青脆枝的发根组织中DXR和HMGR的转录水平,并推测MEP途径是喜树碱合成前者的主要供给者。Yang等(2012)和Yang等(2019)在丹参毛状根中添加洛伐他汀和膦胺霉素,探讨MVA和MEP通路在丹参毛状根细胞生长和次生代谢产物积累中的作用,发现MVA途径在丹参毛状根细胞生长中发挥着重要作用,而MEP途径是丹参次生代谢产物含量的积累的主要途径。但是,有关MVA和MEP途径在秦艽次生代谢产物生物合成途径中的作用机制尚缺乏相关的研究。基于此,本研究依据抑制剂膦胺霉素和洛伐他汀的抑制原理,采用3因素4水平正交试验的方法,通过设置不同浓度的膦胺霉素和洛伐他汀处理一月生秦艽幼苗,选取不同的取样天数,通过评估其次生代谢产物含量的变化,拟筛选可抑制秦艽中马钱苷酸、獐牙菜苷、獐牙菜苦苷、龙胆苦苷含量的最优条件,以期为进一步研究秦艽MEP和MVA代谢途径调控机制和解析环烯醚萜类化合物的合成机理奠定基础。
1 材料与方法
1.1 材料和主要仪器
秦艽种子购自甘肃庆阳正宁县,于温室撒播种子给予16 h光照,8 h黑暗处理,覆膜出苗后培养一个月(叶片2对生,高度2 cm左右)用于试验。LC-20XR分析型高效液相色谱仪(日本岛津公司),色谱柱(Welch,4.6 mm×250 mm,5 μm),精密电子天平(赛多利斯科学仪器有限公司),超声波清洗仪(小美仪器有限公司),OBS-2100油浴锅(上海爱朗仪器有限公司)。
1.2 试剂
马钱苷酸(批号:K17S11B124207,纯度≥98%)、獐牙菜苦苷(批号:Y25J1G1429,纯度≥98%)、龙胆苦苷(批号:Y29A11Q112202,纯度≥98%)、獐牙菜苷(批号:P25O10F101344,纯度≥98%)、洛伐他汀(批号:Y23D6C7375,纯度≥98%)均购自上海源叶生物科技有限公司。膦胺霉素(批号:lX040552,CHEMEGEn公司),甲醇(成都市科隆化学品有限公司),乙腈(上海霍尼韦尔贸易有限公司),磷酸(天津市科密欧化学试剂有限公司)。
1.3 溶液配制
称取0.010 4 g洛伐他汀于50 mL容量瓶中,加入2 mL无水乙醇,混匀后加入20 μL 0.6 mol·L-1的NaOH,静置30 min,加入适量1 mol·L-1 HCl調节pH至7,加纯化水定容至50 mL,得500 μmol·L-1的洛伐他汀母液,4 ℃备用。
称取0.010 0 g膦胺霉素于50 mL容量瓶中,加纯化水定容至50 mL,超声溶解,得到浓度为546 μmol·L-1的膦胺霉素母液,4 ℃备用。
称取马钱苷酸0.95 mg、獐牙菜苦苷1.26 mg、龙胆苦苷2.52 mg和獐牙菜苷1.24 mg的对照品,加甲醇定容至1 mL容量瓶中,混匀,得到质量浓度为马钱苷酸0.95 mg·mL-1、獐牙菜苦苷1.26 mg·mL-1、龙胆苦苷2.52 mg·mL-1和獐牙菜苷1.24 mg·mL-1的母液,4 ℃备用。
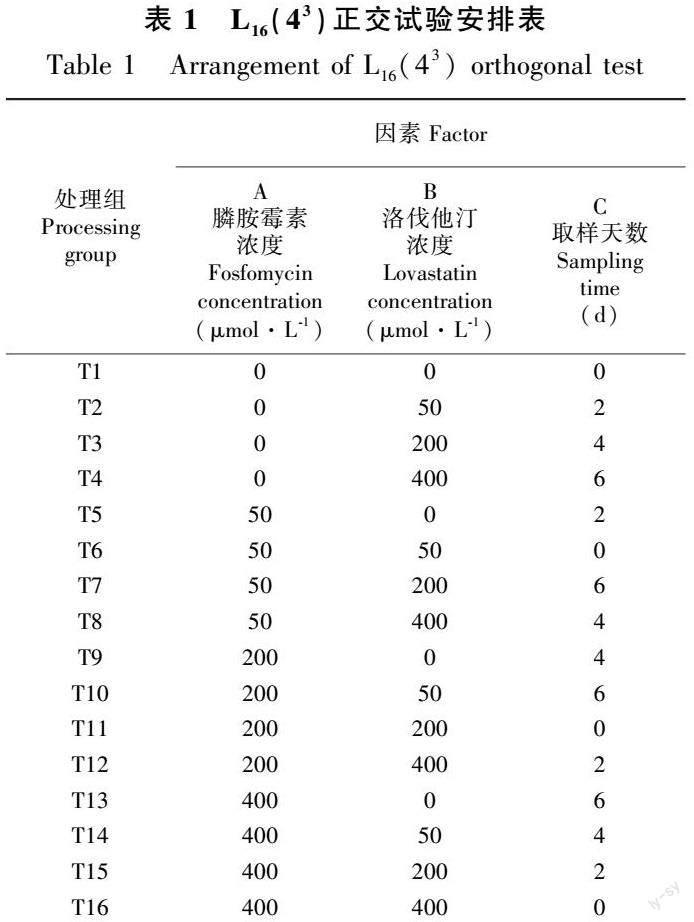
1.4 正交试验及样品处理
参照(于淑池,2013;罗云云,2019)按表1的因素、水平进行试验设计喷施处理16组秦艽幼苗,每组处理30株。处理后放置26 ℃下光照16 h·d-1。取不同处理条件下的幼苗,用纯水冲洗干净幼苗上的土壤和杂质,滤纸吸干水分后置于旋蒸瓶中,加入少量甲醇减压1.5 h抽干水分。将干燥后的秦艽幼苗称重后加入1.5 mL离心管中,加入1 mL甲醇超声提取3次,每次1.5 h,过滤得到绿色的提取液,减压浓缩,得到灰绿色的固体物质,黏附在瓶壁,称重。加入1 mL甲醇溶解,过0.22 μm滤膜,滤液为样品溶液。
1.5 HPLC色谱条件及标准曲线绘制
流动相为0.1%磷酸水∶乙腈,洗脱梯度为0~10 min,12%乙腈;10~13 min,10%乙腈;13~30 min,12%乙腈,流速为0.8 mL·min-1,检测波长254 nm,进样量10 μL,柱温为30 ℃。
将配制好的标准品溶液分别用甲醇按照倍数稀释,制备不同浓度梯度的标准品溶液。马钱苷酸浓度梯度为0.950 0、0.475 0、0.237 5、0.118 8、0.059 4、0.029 7、0.014 9、0.007 5、0.003 8、0.001 9 mg·mL-1;獐牙菜苦苷浓度梯度为1.260 0、0.630 0、0.315 0、0.157 5、0.078 8、0.039 4、0.019 7 mg·mL-1;龙胆苦苷浓度梯度为2.520 0、1.260 0、0.630 0、0.315 0、0.157 5、0.078 8、0.039 4、0.019 7 mg·mL-1;獐牙菜苷浓度梯度为1.240 0、0.620 0、0.310 0、0.155 0、0.077 5、0.193 8、0.009 7、0.004 8、0.002 4 mg·mL-1的对照品溶液,依据上述色谱条件依次进样测定。以进样浓度(X,g·mL-1)和峰面积(Y)绘制标准曲线。马钱苷酸的线性回归方程为Y=4 571 241.60X-11 205.29,线性范围为0.002 0~0.475 0 g·mL-1;獐芽菜苦苷的线性回归方程为Y=6 499 223.38X-24 589.43,线性范围为0.003 0~0.630 0 g·mL-1;龙胆苦苷线性回归方程为Y=14 344 350.83X-442 328.21,线性范围为0.005 0~2.520 0 g·mL-1;獐芽菜苷线性回归方程为Y=6 047 666.40X-40 600.10,线性范围为0.002 0~1.240 0 g·mL-1。
1.6 数据统计
使用Excel 2019对数据进行处理,使用Minitab 18(Minitab公司)对正交试验结果做极差分析,SPSS Statistics 26(IBM公司)对数据进行方差分析和相关性分析,运用GraphPad Prism 5(GraphPad公司)软件作图。验证结果数据采用x±s表示。抑制剂对代谢产物的抑制率计算公式:抑制率=(处理组-空白组)/空白组×100%。
2 结果与分析
2.1 次生代谢产物含量测定结果分析
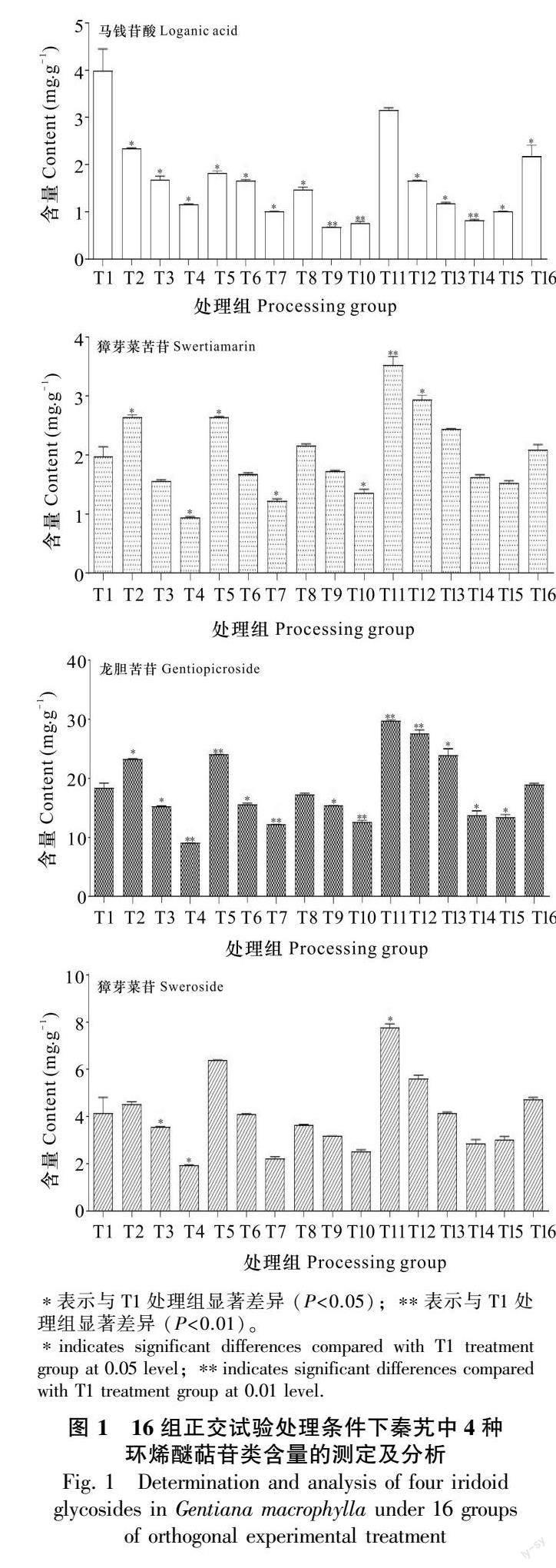
从表1和图1可以看出,除T1(空白组)外,T2至T16处理组马钱苷酸、獐牙菜苦苷、龙胆苦苷和獐牙菜苷的含量变化趋势类似。与T1相比,不同处理条件下马钱苷酸的含量均呈降低的趋势,其中T9、T10、T14处理组的马钱酸苷含量降低最为显著,分别降低了83%、81%、79%,与T1组相比有极显著差异。经比较发现,无论是单一抑制剂还是两个抑制剂共同作用,均可抑制马钱苷酸的合成且在抑制剂浓度相同的条件下膦胺霉素对马钱苷酸的抑制率高于洛伐他汀的作用。与T1组相比,不同的是大部分处理条件下獐牙菜苦苷、龙胆苦苷和獐牙菜苷含量均有所降低,其中T4组降低最为显著,3种代谢产物分别降低了52%、50%、54%,而T2、T5、T12处理组的獐牙菜苦苷、龙胆苦苷和獐牙菜苷的含量不降反升。推测引起此结果的原因可能与代谢途径酶的活性、酶基因的表达等响应抑制剂的强度存在差异有关。
2.2 基于正交试验结果进行多因素方差分析
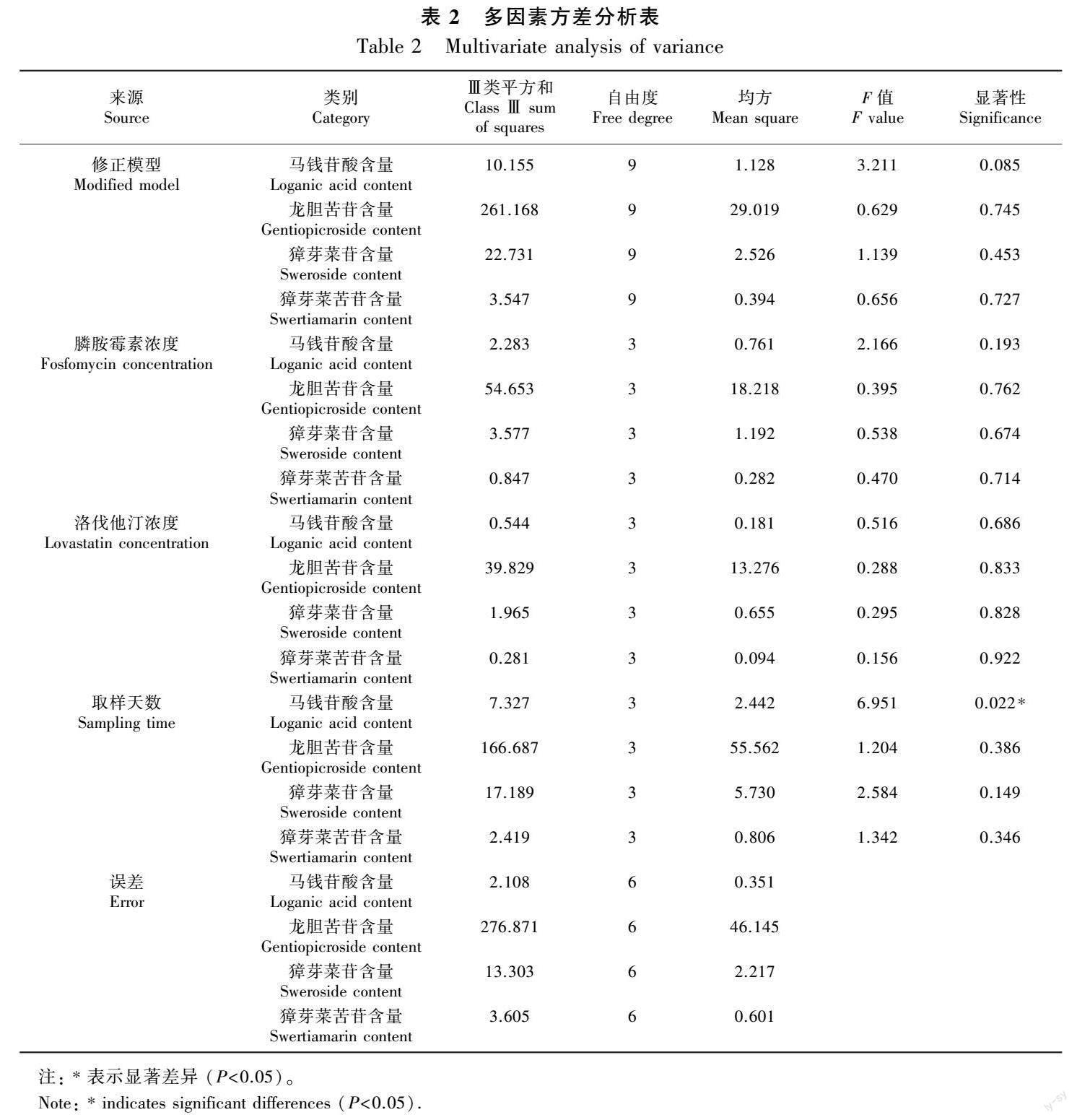
对正交试验结果进行多因素方差分析,发现膦胺霉素和洛伐他汀对秦艽马钱苷酸、龙胆苦苷、獐牙菜苦苷和獐牙菜苷中含量的积累均无显著性影响,仅取样天数对马钱苷酸的含量积累有显著影响,而对龙胆苦苷、獐牙菜苦苷和獐牙菜苷的含量积累均无显著性影响(表2)。
2.3 相关性分析
由表3可知,取样天数与獐牙菜苦苷含量呈显著负相关,与马钱苷酸、龙胆苦苷和獐牙菜苷的含量呈极显著负相关;马钱苷酸与獐牙菜苦苷、龙胆苦苷和獐牙菜苷的含量呈显著正相关;獐牙菜苦苷与龙胆苦苷、獐牙菜苷的含量呈极显著正相关;龙胆苦苷与獐牙菜苷含量呈极显著正相关。当以MEP和MVA途径的抑制剂膦胺霉素和洛伐他汀处理秦艽幼苗时,可显著影响马钱苷酸、獐牙菜苦苷、龙胆苦苷和獐牙菜苷的含量,表明洛伐他汀与膦胺霉素可作为研究环烯醚萜类化合物生源合成途径代谢调控机制的有效抑制剂。取样天数与代谢产物含量呈负相关,表明随着取样天数的延长,代谢产物含量的生成呈降低趋势。此外,4種环烯醚萜化合物之间含量变化呈正相关,推测是抑制剂作用后,代谢流具备连续响应的机制。
2.4 极差分析
由表4可知,影响4种环烯醚萜类化合物含量的因素由强到弱的顺序是取样天数>膦胺霉素浓度>洛伐他汀浓度;取样天数对次生代谢产物含量的影响因素最大,其次是膦胺霉素浓度,洛伐他汀浓度影响最小。
本研究考查抑制剂对秦艽4种环烯醚萜类化合物含量的影响,关注引起化合物含量降低的因素,因此根据极差分析结果(表4),A(4)B(2)C(4)即膦胺霉素400 μmol·L-1,洛伐他汀50 μmol·L-1,处理6 d为马钱苷酸的最佳抑制条件;A(1)B(2)C(4)即膦胺霉素0 μmol·L-1,洛伐他汀50 μmol·L-1,处理6 d为獐芽菜苦苷的最佳抑制条件;A(4)B(2)C(4)即膦胺霉素400 μmol·L-1,洛伐他汀50 μmol·L-1,处理6 d为龙胆苦苷的最佳抑制条件;A(1)B(2)C(4)即膦胺霉素0 μmol·L-1,洛伐他汀50 μmol·L-1,處理6 d为獐芽菜苷的最佳抑制条件。综合考虑,4种代谢产物最佳的抑制条件为A(4)B(2)C(4)即膦胺霉素400 μmol·L-1,洛伐他汀50 μmol·L-1,处理天数为6 d(图2)。
2.5 验证试验
以正交试验设计筛选所得最优抑制条件处理秦艽一月生幼苗,重复3次,检测其次生代谢产物含量,以验证最优抑制条件的可靠性。验证试验结果表明,与空白组相比,处理组中4种环烯醚萜的含量均有不同程度的降低,马钱苷酸、獐芽菜苦苷、龙胆苦苷以及獐芽菜苷含量分别下降了69%、36%、33%和4%(表5)。这表明最优抑制条件为洛伐他汀50 μmol·L-1、膦胺霉素400 μmol·L-1和取样天数6 d。
3 讨论与结论
秦艽作为中国传统药材,环烯醚萜类化合物是其主要的药用有效成分。该类次生代谢产物的生源合成途径先经MVA和MEP途径合成焦磷酸香叶酯,再经过裂环等多步酶促反应合成龙胆苦苷等萜类化合物(康恒等,2021)。本研究以膦胺霉素浓度、洛伐他汀浓度以及取样天数为3个主要因素进行了3因素4水平的正交试验,筛选了可显著抑制马钱苷酸、獐牙菜苦苷、龙胆苦苷和獐牙菜苷的最佳条件。本研究极差分析结果表明,膦胺霉素对代谢产物含量的抑制效果高于洛伐他汀,推测MEP途径在秦艽环烯醚萜类次生代谢产物的合成途径中起主导作用,与王彩云等(2014)基于对滇龙胆转录组数据推测龙胆苦苷生物合成主要来源于MEP途径,部分来源于MVA途径的结论相似。此外,抑制剂对马钱苷酸的含量降低程度显著强于獐牙菜苦苷、龙胆苦苷和獐牙菜苷,
推测马钱苷酸可能相较其余3种代谢产物位于代谢通路的上游,因此抑制剂对马钱苷酸含量的影响最大。相关性分析发现,4种代谢产物含量的变化呈显著正相关,推测抑制剂对其代谢流可能具有“级联趋同”的调控作用。但是,取样天数和代谢产物含量呈显著负相关,即抑制剂处理时间越长,4种环烯醚萜含量的呈显著降低趋势,而关于抑制剂对代谢产物影响的最长时效还需进一步研究。
基于本研究正交法优化的最佳抑制条件为膦胺霉素400 μmol·L-1、洛伐他汀50 μmol·L-1、取样天数6 d。在该条件下马钱苷酸、獐牙菜苦苷、龙胆苦苷和獐牙菜苷的含量均有所下降,分别下降了69%、36%、33%和4%。这表明基于正交法筛选的抑制条件可作为进一步研究秦艽中主要次生代谢产物MVA和MEP合成途径调控机制的最优抑制条件。本研究所得最佳抑制条件,为后期研究秦艽环烯醚萜类化合物生物合成过程MVA和MEP代谢通路的调控机制奠定了一定的基础。
参考文献:
CAO XY, GUO XR, YANG XB, et al., 2016. Transcriptional responses and gentiopicroside biosynthesis in methyl jasmonate-treated Gentiana macrophylla seedlings [J]. PLoS ONE, 11(11): e0166493.
HUA WP, ZHENG P, HE YH, et al., 2014. An insight into the genes involved in secoiridoid biosynthesis in Gentiana macrophylla by RNA-seq [J]. Mol Biol Rep, 41(7): 4817-4825.
KANG H, ZHAO ZL, NI LH, et al., 2021. Transcriptome analysis and validation of key genes involved in biosynthesis of iridoids in Gentiana lhassica [J]. Chin J Trad Chin Med, 46(18): 4704-4711.[康恒, 赵志礼, 倪梁红, 等, 2021. 全萼秦艽转录组中环烯醚萜类相关基因挖掘及验证 [J]. 中国中药杂志, 46(18): 4704-
4711.]
LI GW, WANG L, 2018. Anti-inflammatory and analgesic effects of QinJiao in the treatment for arthritis [J]. W Trad Chin Med, 31(3): 133-136.[李跟旺, 王磊, 2018. 秦艽在关节炎抗炎镇痛治疗中的作用 [J]. 西部中医药, 31(3): 133-136.]
LIAO P, HEMMERLIN A, BACH TJ, et al., 2016. The potential of the mevalonate pathway for enhanced isoprenoid production [J]. Biotechnol Adv, 34(5): 697-713.
LI T, YU X, REN YM, et al., 2022. The chromosome-level genome assembly of Gentiana dahurica (Gentianaceae) provides insights into gentiopicroside biosynthesis [J]. DNA Res, 29(2): 1-10.
LIU L, 2012. Research on MVA and MEP ginsenoside biosynthesis pathway by utilizing inhibitors [D]. Changchun: Jilin University.[刘丽, 2012. 利用抑制剂对人参皂苷生物合成途径MVA与MEP的研究 [D]. 长春: 吉林大学.]
LUO YY, DU WF, YING ZX, et al., 2019. Optimization of extraction process conditions of coix seed polysaccharide by response surface method and orthogonal of design experiment[J]. Chin J Trad Chin Med, 34(10): 4847-4851.[羅云云, 杜伟锋, 应泽茜, 等, 2019. 响应面法协同正交试验设计优化薏苡仁多糖提取工艺研究 [J]. 中华中医药杂志, 34(10): 4847-4851.]
MIETTINEN K, DONG L, NAVROT N, et al., 2014. The seco-iridoid pathway from Catharanthus roseus [J]. Nat Comm, 5: 3606.
MUHAMAD FADZIL NS, MAHENDRAN S, SIEW HG, et al., 2021. Chemistry, pharmacology and therapeutic potential of Swertiamarin — A promising natural lead for new drug discovery and development [J]. Drug Des Dev Ther, 15: 2721-2746.
National Pharmacopoeia Committee. Pharmacopoeia of the Peoples Republic of China [S]. Part I. Beijing: The Medical Science and Technology Press of China, 2020: 282.[国家药典委员会. 中华人民共和国药典 [S]. 一部. 北京:中国医药科技出版社, 2020: 282.]
RATHER GA, SHARMAA, JEELANI SM, et al., 2019. Metabolic and transcriptional analyses in response to potent inhibitors establish MEP pathway as major route for camptothecin biosynthesis in Nothapodytes nimmoniana (Graham) Mabb [J]. BMC Plant Biol, 19(1): 301.
PENG MC, AI XH, 2021. Research progress in chemical constituents, pharmacological effects and clinical application of the flowers of Gentianae macrophylla [J] Centr S Pharm, 19(6): 1243-1249.[彭美晨, 艾晓辉, 2021. 秦艽花化学成分、药理作用及其临床应用的研究进展 [J]. 中南药学, 19(6): 1243-1249.]
SINHA RK, SHARMA SN, VERMA SS, et al., 2018. Effects of lovastin, fosmidomycin and methyl jasmonate on andrographolide biosynthesis in the Andrographis paniculata [J]. Acta Physiol Plant, 40(9): 165.
TUNDIS R, LOIZZO MR, MENICHINI F, et al., 2008. Biological and pharmacological activities of iridoids: recent developments [J]. Mini Rev Med Chem, 8(4): 399-420.
WANG CY, ZHANG XD, SHEN T, et al.,2014. Research progress of biosynthesis pathway of gentiopicroside[J]. Jiangsu Agric Sci, 42(3): 4-9.[王彩云, 张晓东, 沈涛, 等, 2014. 龙胆苦苷生物合成途径研究进展 [J]. 江苏农业科学, 42(3): 4-9.]
WEI JH,2013. Research on biosynthesis manipulation of isoprenoids based on HMGR and DXR genes fromAmomum villosum Lour [D]. Guangzhou: Guangzhou University of Chinese Medicine.[魏洁书, 2013. 基于阳春砂HMGR和DXR基因的萜类化合物生物合成调控研究 [D]. 广州: 广州中医药大学.]
WU XY, LIU XL, 2017. Progress of biosynthetic pathway and the key enzyme genes of iridoids [J]. Chin J Ethnomed Ethnopharm, 26(8): 44-48.[吴昕怡, 刘小莉, 2017. 环烯醚萜类成分生物合成途径及关键酶基因研究进展 [J]. 中国民族民间医药, 26(8): 44-48.]
YANG DF, DU XH, LIANG X, et al., 2012. Different roles of the mevalonate and methylerythritol phosphate pathways in cell growth and tanshinone production of Salvia miltiorrhiza hairy roots [J]. PLoS ONE, 7(11): e46797.
YANG FX, WANG Y, XIA PF, et al., 2020. Research progress on chemical constituents and pharmacological effects of Gentianae macrophyllae Radix and quality markes(Q-marker) prediction and analysis[J]. Chin Trad Herb Drug, 51(10): 2718-2731.[杨飞霞, 王玉, 夏鹏飞, 等, 2020. 秦艽化学成分和药理作用研究进展及质量标志物(Q-marker)的预测分析 [J]. 中草药, 51(10): 2718-2731.]
YANG YF, HOU S, FAN W, et al., 2019. Expression patterns of some genes involved in tanshinone biosynthesis in Salvia miltiorrhiza roots [J]. Ind Crops Products, 130: 606-614.
YU SC, DENG HY, JIANG Y, et al.,2013. A research on optimum conditions for algal inhibition by reed extract [J]. Acta Hydrobiol Sin, 37(6): 1051-1058.[于淑池, 鄧红英, 姜燕, 等, 2013. 芦苇秆浸出液抑藻的最佳环境条件研究 [J]. 水生生物学报, 37(6): 1051-1058.]
(责任编辑 李 莉)
