Wnt/β-catenin信号通路在骨质疏松症环境下促进成骨的研究进展
2023-05-21王浩阗邵阳汪国澎叶灵山王秋波王建伟
王浩阗 邵阳 汪国澎 叶灵山 王秋波 王建伟,★
骨质疏松症(osteoporosis,OP)是由多因素引起的,其是以骨量丢失、骨骼结构退化、骨骼强度变低为特征的一种全身性骨代谢疾病[1],位居危害老年人群身体健康的慢性病首位。罹患骨质疏松症将大大增加老年人口骨折的风险,甚至导致死亡的发生[2]。近年来,我国社会老龄化逐年加剧,截至2020 年底我国60 岁以上人口已超过2.6 亿,约占总人口的18.7%[3],未来几年内老龄化趋势仍可能继续,对我国公共卫生系统挑战巨大。
Wnt/β-catenin通路是调节骨重建平衡的关键信号通路之一[4],本文将结合国内外最新研究进展,探究Wnt/β-catenin信号通路在骨质疏松环境下的作用,以期为治疗、预防骨质疏松症提供参考。
1 Wnt/β-catenin 与骨质疏松的关系
1984 年,Nusse 首先在小鼠乳腺癌中发现Int-1基因,并且Int-1与1976 年Sharma 和Chopra 发现的Wingless基因具有高度同源性,合称为Wnt基因,首次提出Wnt概念。时至今日,Wnt通路已是目前研究较为明确的通路,其具有调节多种细胞的功能,在多个环节发挥重要的生理作用,已被证实与心血管疾病、糖尿病、癌症、骨质疏松等多种疾病相关。
Wnt信号通路分为依赖β-catenin的经典信号通路和非依赖β-catenin的信号通路,如Wnt/Ca+通路等。本文主要探讨Wnt/β-catenin途径,在Wnt未被抑制下,Wnt与低密度脂蛋白受体相关蛋白LRP 5/6(LDL receptor related protein5/6,LRP 5/6)和卷曲受体蛋白(frizzled,Fzd)组成的双受体复合物结合,启动Wnt/β-catenin信号传导。轴蛋白(Axin)通过与起正调控作用的蓬乱蛋白(dishevelled,Dsh)相互作用,被Fzd 招募移动到细胞膜上LRP5/6 处,与FRAT1 和糖原合成酶激酶-3β(glycogen synthase kinase-3β,GSK-3β)形成复合物,以阻止β-catenin 的磷酸化及其蛋白质体降解。细胞质内β-catenin 水平上升并积聚,进而易位到细胞核,和核内转录因子T 细胞因子/淋巴增强因子TCF/LEF(T cell factor/Lymphoid enhancing factor,TCF/LEF)结合,从而激活转录因子活性调节靶基因表达;反之,在Wnt被抑制的情况下,Axin 和腺瘤性息肉病蛋白(adenomatous polyosis coli,APC)将β-catenin 带到GSK-3β 和酪蛋白激酶1(casein kinase 1,CK1),导致其在Axin-APC-GSK-3β 特定丝氨酸/苏氨酸残基上磷酸化。β-catenin(p-βcatenin)磷酸化后被靶向进行泛素化(主要通过E3连接酶B-TrCP)和蛋白质体破坏。见图1。
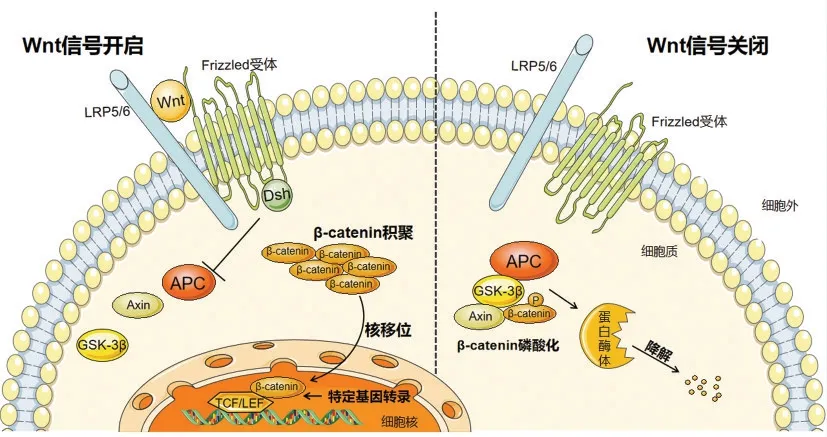
图1 Wnt/β-catenin 信号通路Figure 1 Wnt/β-Catenin signal pathway
Wnt/β-catenin通路在调节成骨细胞分化、骨形成和内环境稳定中发挥重要作用。2001 年研究报导LRP 5 相关突变将影响成骨过程,进而导致骨质疏松症等,第一次发现Wnt/β-catenin为调节骨骼健康与疾病的关键信号通路,确立了其与OP 发病的密切联系[5]。随后,一系列关于Wnt/β-catenin通路与骨形成的因子相继被发现,如Runt 相关转录因子2(recombinant runt related transcription factor 2,RNUX2)、成骨相关转录因子抗体(osterixl,OSX)和骨保护素(osteoclastogenesis inhibitory factor,OPG)对于成骨细胞分化至关重要,RNUX2 作为早期关键转录因子,促进间充质干细胞(bone marrow mesenchymal stem cells,BMSCs)向前成骨细胞分化;而前成骨细胞的成熟是在OSX 作用下实现的,OPG 可以抑制BMSC 向破骨细胞分化,并且该通路中ALP、Ⅰ型胶原重组蛋白α1(recombinant Collagen TypeⅠAlpha 1,Col1α1)和骨钙素(osteocalcin,OCN)等成骨特异性基因的表达都与通路中RNUX2 和OSX 的表达关联[6-7]。
2 Wnt/β-catenin 信号通路在BMSCs 向成骨细胞分化中的调控作用
非瑟酮和β-catenin 促进了RNUX2 和OSX 的表达,非瑟酮可通过在Ser9 位点磷酸化GSK-3β 而促进前成骨细胞β-catenin 的核移位,进而提升ALP 活性和成骨细胞结节的矿化。特异性阻断β-catenin 和稳定GSK-3β 后,非瑟酮依然可以增强前成骨细胞的成骨活性,表明非瑟酮通过阻断Wnt/β-catenin通路中GSK-3β 介导的β-catenin 降解,进而促进RNUX2 的转录刺激成骨[6]。研究报道,褪黑素对BMSCs 的存活以及分化成骨、成脂谱系存在影响,褪黑素上调肝细胞生长因子的同时下调抑癌基因人第10 号染色体缺失的磷酸酶及张力蛋白同源的基因(phosphatase and tensin homolog deleted on chromosome ten,PTEN)的表达,Wnt和β-catenin 表达激活了Wnt/β-catenin通路,促进了BMSCs 增殖和成骨分化[8]。过度表达隐花色素1(recombinant cryptochrome 1,Cry1),β-catenin 表达上调,GSK-3β 被磷酸化,从而促进BMSCs 成骨分化;此外,发现RNUX2基因的启动子和生物钟基因(Aryl Hydrocarbon Receptor Nuclear Translocator Like Protein,ARNTL)的E-box基因结合位点之间的相似片段,ARNTL可能对RNUX2 具有调节作用,这提醒ARNTL作为RNUX2 的转录抑制剂,Cry1 可能对RNUX2 有直接的调节作用[9]。目前有关Cry1 调节人成骨细胞成骨分化的机制相关报道仍然较少,还需要进一步的体内和体外实验验证Cry1 治疗骨缺损的可行性和临床价值,以及揭示Cry1 调控成骨的确切分子机制。17β-雌二醇(17β-Estradiolum,17β-E2)通过将GSK-3β 磷酸化,显著增加β-catenin 的水平,激活雌激素受体ER/GSK-3β依赖的Wnt/β-catenin通路;免疫荧光和免疫沉淀分析显示,17β-E2 显著诱导β-catenin 向细胞核移位,与TCF/LEF 结合并形成β-catenin/TCF 蛋白复合物,激活靶基因表达,调节增殖因子(cyclin D1)和成骨细胞分化因子(BGP、ALP 和OPN)表达,促进成骨细胞的增殖和分化[10]。另有体外实验表明,使用17β-E2 可以促进骨膜蛋白(Recombinant Periostin,Postn)表达,Postn mRNA 表达上调,免疫荧光分析显示Postn 定位于细胞质中,Wnt3a、β-catenin 表达随之上升,进而促进RNUX2、ALP 表达,这解释了雌激素限制骨膜骨扩张并且刺激骨内膜骨附着方面的作用。与之对比,在甲状旁腺激素治疗后Postn小鼠近端胫骨纵向切片中的Postn 在骨膜处表达上调,但在皮质表面表达不明显,说明Postn 很可能在骨骼形成和形态维持中起重要作用。Postn 的极化表达也可能对甲状旁腺激素和17β-E2 在骨膜与皮质内表面的不同影响和效力产生影响。有两种主要机制可以促进OVX BMSCs 中的Postn 激活,首先,17β-E2 以配体依赖方式与BMSCs 细胞膜中的ER-α 或ER-β 结合,从而调节转录并促进Postn 激活;其次,17β-E2 通过细胞因子促进Postn 的表达。TGF-β 可以诱导BMSCs 分泌Postn 以支持骨形成,Postn 可能通过自分泌/旁分泌机制调节Wnt信号传导,是骨合成代谢的关键介质。研究证实Postn 是雌激素靶向的,通过Wnt/β-catenin途径在雌激素调节的骨形成过程中起重要作用[11]。成骨细胞分化的早期阶段涉及细胞增殖,RNUX2 的表达对成骨分化至关重要。在Wnt 信号转导下,β-catenin 转位通过TCF/LCF 的结合诱导RNUX2 表达。使用2,4,5 三甲氧基黄连醌(TMDQ)处理成骨细胞后,Wnt1、Wnt3a mRNA 和磷酸化GSK3 的表达上调,Bmp2 和Bmp4 mRNA 表达上调,成骨细胞中总β-catenin 和核β-catenin 的水平增加,同时β-catenin下游靶基因RNUX2表达水平显著增强,RNUX2进一步靶基因ALP、OCN、骨唾液蛋白(bone sialoprotein,BSP)和OPN mRNA 的表达同时上调,并且观察到β-catenin 快速核移位,证实了TMDQ 通过Wnt/β-catenin促进成骨分化[12]。研究人员使用药物棉酚处理OVX 小鼠发现Wnt信号在小鼠体内上调,并且CK1、Axin、β-catenin 等中下游分子显著上调,通路内Wnt 抑制因子人Wnt 抑制因子1(Wnt inhibitory factor-1,WIF1)的表达明显减少,是通过促进CpG 岛的甲基化水平,Wnt上调,OPG、血清降钙素的水平随之上调,破骨细胞分化因子(Receptor Activator of Nuclear Kappa-B Ligand,RANKL)的表达显著下调实现的[13]。
3 Wnt/β-catenin 通路通过抑制BMSCs 成脂促进成骨
近年来,骨生成和脂肪生成之间的不平衡也被认为是骨质疏松症最重要的发病机制之一[14]。正常情况下,BMSCs 被招募到特定部位,并分化为成骨细胞以补充骨量,然而,在老年骨质疏松症患者中,BMSCs 倾向于分化为脂肪细胞而不是成骨细胞,从而导致脂肪累积和骨丢失,这种分化倾向是由Wnt信号调控的[15]。在生长骨软骨细胞中条件敲除β-catenin基因的小鼠中发现,由于β-catenin基因的条件性缺失,导致了软骨-骨连接处成骨减少而成脂增加,基因敲除小鼠表现出骨质疏松表型[12]。β-catenin 信号作为主调节因子决定了BMSCs 向成骨细胞而不是脂肪细胞的分化[16-17]。研究使用虎杖苷(OG)处理BMSCs,显示RNUX2、骨钙素mRNA 和蛋白质表达增强,结合Western blot 分析,经OG 处理后OVX BMSCs 中活性β-catenin(α-ABC)水平升高至正常水平,并且处理后α-ABC 在BMSCs 的细胞核中积累,促进了α-ABC 的转录活性,说明OG 通过激活β-catenin通路增强了BMSCs 的成骨分化[18];qPCR 分析显示,OG 处理后BMSCs 成脂分化过程中三种成脂标志物——转录因子PPARγ、C/EBPα 和脂联素的mRNA 表达受到抑制,同时三种标记物的蛋白质表达水平相应下降;用油红O 染色法进行监测,经OG 处理BMSCs 细胞中的脂肪生成较少,这些结果表明OG 抑制了BMSCs 的成脂分化;随后使用Wnt/β-catenin抑制剂(ICG-001)处理OG-VOX BMSCs 后,BMSCs 成脂分化和脂滴形成的抑制效应被显著消除,这表明Wnt/β-catenin信号通路还介导了OG 对BMSCs 成脂分化的抑制作用。
4 中药通过Wnt/β-catenin通路治疗骨质疏松
中药因具有多靶点、不良反应小、来源广泛等优势,被越来越多的科学家所关注,2017 年中华医学会骨质疏松分会发布的指南中就明确了中药治疗的重要地位。中医药治骨质疏松讲究从肾论治的独特思维,因此诸如天然植物黄酮芹菜素[19]、人参皂苷[20]、葛根素[21]、肉豆蔻[22]、左归丸[23]、右归丸[24]等中药、方剂被广泛研究,这为骨质疏松症的治疗提供了更多选择和可能[25-26]。据报道,使用骨碎补总黄酮与碳酸钙联合治疗OVX 大鼠,观察到Wnt3a 表达显著增强,p-β-catenin 表达显著降低,这说明治疗后Wnt3a/β-catenin通路的活性明显得到激活;并且还观察到抑制了血清中ALP、抗酒石酸酸性磷酸酶(tartrate resistant acid phosphatase,TRACP)升高,通过ER-α 依赖机制增强成骨细胞OPG 的生成,同时只有骨碎补总黄酮与碳酸钙联合使用时,RANKL基因的表达才会显著抑制,提示这个过程和上调Wnt3a、β-catenin和下调RANKL 有关,促进了成骨分化和抑制破骨细胞生成,从而维持β-catenin 的转录调节活性[26]。OP 是糖皮质激素治疗的常见副作用,继发性骨质疏松症是最常见的形式[27]。OP 中的骨内稳态失衡是由骨吸收增加和成骨减少的影响引起的,使用中高剂量中药古米宝汤干预OP 大鼠BMSCs 后,Dickkopf 相关蛋白1(Dickkopf Related Protein 1,DKK1)和组织蛋白酶K(Cathepsin,CTSK)的mRNA 表达下调,RNUX2 的mRNA 表达上调,这提示中高剂量古米宝汤可以通过Wnt/β-catenin通路抑制BMSCs中的DKK1 和CTSK,从而抑制破骨并且上调RNUX2促进成骨,从而调节骨代谢防治OP[28]。
5 结语与展望
Wnt/β-catenin信号通路在骨质疏松中对于BMSCs 向成骨细胞分化,对成骨细胞增殖、分化和活性,以及抑制脂分化起调控作用,其中一些作用和机制已被证实,在再生医学、干细胞研究和临床诊断治疗中具有极高的应用价值。但仍有不少挑战,包括如何使药物作用于靶细胞发挥作用,如何传递细胞因子、控制细胞的激活,如何提高标志物兼具敏感性和特异性,如何降低干细胞移植的免疫排斥反应等,都需要进一步深入研究。未来可将Wnt/β-catenin信号通路作为切入点,通过调控该信号通路的活性来达到对骨质疏松等骨代谢疾病的预防、诊断与治疗的目的。
