Interplay of Cyclic Electron Transport and Mehler-like Reaction in Synechocystis Under Different Light Regimes
2023-05-17ZHANGYeZHANGHaoZHANGPengpeng
ZHANG Ye, ZHANG Hao, ZHANG Pengpeng
(Biotechnology Research Institute, Chinese Academy of Agricultural Sciences, Beijing 100081, China)
Abstract:Light is an essential substrate for photosynthesis. However, excessive light could cause oxidative stress and severe damages of photosynthetic organisms. To cope with constantly changing light environments, cyanobacteria have developed flexible electron transport network. Cyclic electron transport (CET) around photosystem I (PSI)recycles electrons from ferredoxin to plastoquinone pool and generates ATP without NADPH accumulation. Two distinct CET pathways consisting of NDH-dependent route and PGR5-dependent route have been identified both in cyanobacteria and plants. Cyanobacterial flavodiiron proteins Flv1 and Flv3 functioning in Mehler-like reaction accept electrons after PSI to reduce oxygen to water without formation of reactive oxygen species. Here, we studied the function of CET routes and Mehler-like reaction in Synechocystis by physiological characterization under different light regimes and kinetic analysis of P700 oxidation/reduction using various CET and Flv mutants. It showed that NDH-1 complexes contributed over 90% of CET and sustained cell growth under steady high light condition, while Mehlerlike reaction activated rapidly was responsible for releasing transient high light pressure. We proposed that NDH-1 dependent CET was the major mechanism in Synechocystis robustly supporting high light acclimation, whereas Mehler-like reaction was the spare route operating when the existing main path was largely insufficient. The fast responded FLV route could be a complementation of CET in the WT as well as NDH-1 defected mutant M55.
Key words:cyclic electron transfer; NDH-1 complex; Mehler-like reaction; PGR5; P700 kinetics; Synechocystis
Light is an essential environmental factor for all photosynthetic organisms, including higher plants, algae and cyanobacteria. Light energy absorbed by antenna pigments excites photosystem I (PSI) and photosystem II(PSII), leading to charge separation, which is further stabilized by electron transfer[1-3]. Although the photosynthesis is driven by light, the amount of light absorbed that exceeds the capacity of metabolic reactions including CO2fixation inevitably leads to decrease of photosynthetic efficiency. Over excitation caused accumulation of photosynthetic electrons at acceptor side of PSI and production of reactive oxygen species (ROS), which generated oxidative stress[4]. To protect PSI from photodamage, cyanobacteria had developed a flexible electron transport network[5-6], as well as antioxidant enzyme system[7].
The cyclic electron transport (CET) around PSI was one of the most important auxiliary electron transport pathways in cyanobacteria and chloroplasts[8]. Recycling electrons from ferredoxin to plastoquinone pool (PQ pool) and producing extra ATP without net NADPH formation, CET had shown to be essential for photosynthesis under both stress and non-stress conditions[9]. Two distinct CET pathways had been identified both in cyanobacteria and in higher plants. One pathway depended on NAD(P)H dehydrogenase type 1 (NDH-1) complexes[10-11],and the other pathway relied on PROTON GRADIENT REGULATION 5 (PGR5)[12-13]. Although CET had been discovered by Arnon and his colleagues[14]for more than half century, the physiological significance of CET had been neglected until Munekage and his co-workers[15]addressed the essential role of CET for photosynthesis. Unlike the dominant roles of PGR5-CET in plants, NDH-1 complex dependent CET had shown physiological importance whereas PGR5 route was negligible in cyanobacteria[10,13]. Reverse genetics and functional proteomics had demonstrated the presence of functional distinct NDH-1 complexes in cyanobacteria[16-17]. The NDH-1 complexes containing NdhD1/NdhD2 and NdhF1 subunits (NDH-11,2) were involved in cellular respiration, while NDH-1 complexes containing NdhD3/NdhD4 and NdhF3/NdhF4 (NDH-13,4) were specific in CO2uptake. Both types of NDH-1 contribute to CET[18-20].
When photosynthetic light reaction produced“extra” electrons, e.g. high light condition, oxygen was reduced at acceptor side of PSI to form superoxide radical (O2·-) in plant chloroplasts, which was called Mehler reaction[21]. The ROS produced in Mehler reaction was detoxified by antioxidant enzymes. The superoxide was first converted to hydrogen peroxide(H2O2) by superoxide dismutase (SOD). And the H2O2was further reduced to H2O by peroxidases[4]. The Mehler reaction had long been considered to protect chloroplasts under stress conditions[22]. InSynechocystissp. PCC 6803 (Synechocystis), a Mehler-like reaction,consuming electrons from PSI acceptor side to reduce O2to H2O without ROS formation, was identified[23].Two flavodiiron proteins Flv1 and Flv3 were involved in the reaction[24], which were essential to the growth of cyanobacteria under fluctuating light[25]. In addition to Mehler-like reaction, cyanobacteria also possessed antioxidant enzymes for ROS scavenging, including SODs, catalases and peroxidases[26-28]. Compared to sophisticated ROS scavenging system in plants,cyanobacteria has much simple system with less type and number of antioxidant enzymes. InSynechocystis,slr1516is the only gene encoding SOD and suggested to be essential[29].
The CET and Mehler-like reaction have been well elucidated, however, the underlying interactions of these two electron pathways nearby PSI acceptor side remain unclear. To this end, variousSynechocystisCET and Flv mutants were constructed and investigated. The significance and communication of these different electron routes under various light regimes were discussed. A working hypothesis was summarized.
1 Methods
1.1 Construction of different Strains
TheSynechocystissp. PCC 6803 glucose-tolerant strain[30]was used as the wild type. The primers used in this study were summarized in Table 1.PrimerspUC-PGR5-up-F-HindIII,pUC-PGR5-up-RBamHI,pUC-PGR5-down-F-KpnI andpUC-PGR5-down-R-EcoRI were used to generate a construct containing a spectinomycin-resistant cassette for replacement of the position 54 to 198 ofpgr5. The Δpgr5and M55/Δpgr5were generated by transforming the construct into WT and M55[31],respectively. PrimerspUC-FLV1-up-F-BamHI,pUCFLV1-up-R,pUC-FLV1-down-F-EcoRI andpUCFLV1-down-R-KpnI were used to generate a construct containing a chloramphenicol-resistant cassette for replacement of the position 1 to 1 651 offlv1. The Δflv1, Δflv1/Δpgr5,flv1/M55,flv1/M55/Δpgr5mutants were generated by transforming the construct into WT, Δpgr5, M55 and M55/Δpgr5,respectively.
According to Eisenhut et al.[32]with some modifications, an over-expression vector containing theflv1encoding region was generated (Table 1). An amplicon containing thepetJpromoter and oop terminator was amplified by primerspetJ-PstI and petJ-SalI-oop-BamHI, and was inserted into the pUC18 containing a spectinomycin-resistant cassette.PrimersFlv1-F-SalI andFlv1-R-His-SalI were used to amplify theflv1encoding region and the sequence encoding 6 histidine residues. This fragment was inserted to theSalI site at the 5’-end of the oop terminator. ThespkAgene inSynechocystisWT was naturally disrupted by a frameshift[33], therefore could be used as a neutral integration site for gene expression. The primersspkA-up-F-PstI,spkA-up-RPstI,spkA-down-F-EcoRI andspkA-down-R-EcoRI were used to amplify the 5’ and 3’-flanking regions of the neutral site, which were then inserted into thePstI site andEcoRI site of the overexpression vector respectively. The constructed plasmid was transformed into WT, Δflv1andflv1/M55 respectively, so that the fragment includingpetJpromoter,flv1, oop terminator and a spectinomycin-resistant cassette was integrated into thespkAsite.
Segregations of all mutants were verified by PCR. PrimersPGR5-S-F andPGR5-S-R were used for verifying PGR5-deficient mutants. PrimersFLV1-S-F andFLV1-S-R were used for FLV1-deficient mutants (Table 1).
1.2 Cell growth and treatment conditions
The culture conditions were set according to Zhang et al.[17]. In general, the cells were cultivated in liquid medium or on agar plates under 50 µmol photons m-2·s-1continuous light at 30 ℃ with airlevel CO2(LC) or CO2enriched air (3% (v/v), high CO2, HC).All strains were grown in BG-11 medium buffered with 10 mmol·L-1TES-KOH (pH 8.3) for cultures in the air-level CO2(LC), or 20 mmol·L-1HEPES-NaOH (pH 7.5) for high CO2condition (HC).Proper antibiotics were supplemented for mutant strains.
For high light treatment, cells were plated on agar plates under the illumination of 180 µmol photons·m-2·s-1. For fluctuating light treatment, the light regime was set as following: 20 µmol photons·m-2·s-1for 6 min and 400 µmol photons·m-2·s-1for 1 min.For fluctuating light and dark treatment, the light regime was set as following: 20 µmol photons·m-2·s-1for 6 min, 400 µmol photons·m-2·s-1for 1 min, dark for 5 min and this cycle repeated for 16 h followed by complete darkness for 8 h in a day. For hydrogen peroxide treatment, the cells were inoculated in BG-11 medium supplemented with different amount of hydrogen peroxide with final concentrations of 0.0, 1.0,1.5, 2.0, 3.0 mmol·L-1, respectively.
1.3 RNA isolation and quantification
Total RNA was isolated by Trizol method according to Zhang et al.[34]. After removal of the genomic DNA, the first-strand cDNA was synthesized from 2 µg total RNA using Revert aid First Strand cDNA Synthesis Kit (Thermo Scientific). The quantitative Real-Time (qRT-PCR) was performed on an ABI7500 system. ThernpBwas used as a reference gene. PrimersqrnpB-F andqrnpB-R,qsll1521-F andqsll1521-R were designed to generate 150~200 bp amplicons ofrnpBandflv1gene,respectively (Table 1). The change inflv1gene expression relative to that of the WT normalized tornpBwas calculated using 2-ΔΔCTmethod[35].
1.4 Isolation of membrane and soluble fractions
The membrane and soluble fractions were isolated according to Zhang et al.[36]. Cells grown at high and airlevel CO2were collected and washed by centrifugation at 5 000 r·min-1, 4 ℃ for 5 min, and then resuspended in Resuspension Buffer (50 mmol·L-1HEPES-NaOH pH 7.5, 30 mmol·L-1CaCl2, 800 mmol·L-1sorbitol and 1 mmol·L-16-aminohexanoic acid). Cells were broken in the presence of glass beads (150~212 µm,Sigma) by manually vortexing eight times at maximal speed for 1 min at 4 ℃ with 1 min cooling on ice between cycles. Glass beads and cell debris were removed by centrifugation at 2 000 g for 5 min at 4 ℃. The supernatant containing both the membrane and the soluble fractions was centrifuged at 30 000 g for 30 min at 4 ℃. The supernatant was collected as the soluble fraction, and the membrane was obtained from the pellet resuspended in Storage Buffer(50 mmol·L-1HEPES-NaOH pH 7.5, 30 mmol·L-1CaCl2, 600 mmol·L-1sucrose and 1 mol·L-1betaine).
1.5 Detection of SOD activity
The SOD activity was detected by inhibition of the pyrogallol (1,2,3-benzenetriol) autoxidation method[37]. This method was based on the fact that the pyrogallol autoxidation rate monitored by spectrophotometry was inhibited by SOD. One unit of SOD activity was defined as, in certain conditions,the number of enzymes that inhibited the rate of pyrogallol autoxidation by 50% per ml per min in absorbance at 325 nm. Cells were collected,resuspended in a buffer containing 50 mmol·L-1Tris-HCl pH 8.2, 1 mmol·L-1EDTA and 1% (w/v)polyvinylpyrrolidone, and then broken with glass beads as described above. The crude enzyme extracts were collected from the supernatant after centrifugation at 30 000 g for 30 min at 4 ℃. A 3-ml assay was built by adding 50 µL of 4.5 mmol·L-1pyrogallol or 10 mmol·L-1HCl (blank) into 2.95 mL 100 mmol·L-1Tris-HCl (pH 8.2). The rate of pyrogallol autoxidation was determined by monitoring the absorbance at 325 nm every 15 s over 3 min(ΔA325). For measuring SOD activity of samples,30 µL crude enzyme extract was supplemented in the assay and the absorbance at 325 nm was monitored. The rate of pyrogallol oxidation in the presence of the sample was calculated as ΔB325. SOD activity was calculated according to the following formula.
Where y was the SOD activity expressed by U·mg-1protein, n was the dilution ratio of the sample, and C was the protein concentration of the sample (mg·mL-1).
1.6 Electrophoresis and immunoblotting
Proteins of membrane and soluble fractions were separated by SDS-PAGE according to Zhang et al.[17].Samples were denatured and solubilized in Laemmli sample buffer[38]containing 5% β-mercaptoethanol and 6 mol·L-1urea, and were separated in 12%polyacrylamide gel with 6 mol·L-1urea. After electrophoresis, proteins were electrotransferred to a polyvinylidene fluoride membrane by a semi-dry apparatus and immunodetected with specific antibodies against D1, PsaB, NdhK, AtpB, CpcA,FNR and RbcL, respectively. The D1, PsaB, AtpB and RbcL antibodies were purchased from Agrisera.The NdhK, CpcA and FNR antibodies were kind gifts mentioned in the acknowledgement. The relative expression was semi-quantified by comparison to the WT grown at LC. Signal intensities were analyzed by ImageJ.
1.7 P700 measurements
According to Bernat et al.[19]with some modifications, P700oxidation-reduction kinetics was measured quantitatively to determine electron transport rates. When excited by far-red light, the subsequent rereduction rate of P700+(KNO) reflected the difference between electron inflows of metabolites (KM) with cyclic electron transfer (KC) and outflow of terminal oxidases (KR).
In the presence of methyl viologen (MV) which accepted electrons directly from acceptor side of PSI, CET was blocked and the P700+rereduction rate(KMV) reflected the difference between electron inflow of metabolites (KM) and outflow of terminal oxidases (KR).
When the electron efflux to O2(respiration) was blocked by KCN or NaN3, the P700+rereduction rate(KNaN3) presented the sum of the CET and the electron inflow of metabolites.
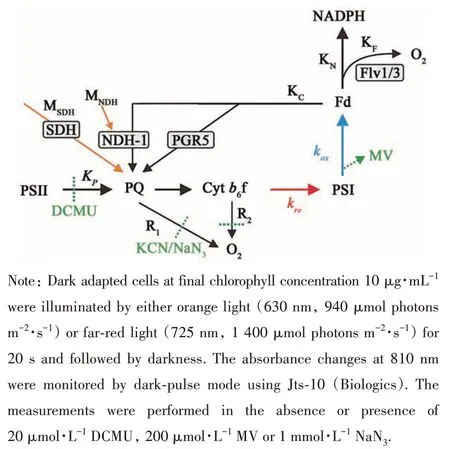
Fig. 1 Major electron transport pathways around PSI
2 Results
2.1 Construction of flv1 mutants in cyclic electron transfer deficient background
To explore the interplay of Mehler-like reaction and CET, various mutants were generated. Twopgr5inactivation mutants (Δpgr5and M55/Δpgr5) were generated. In Δpgr5and M55/Δpgr5, only a single band (~5 kb) different to that of the WT was obtained, indicating that thepgr5gene was completely knocked out (Figure 2A). Severalflv1mutants were attempted in WT, Δpgr5, M55 and M55/Δpgr5. PCR results showed that the Δflv1and Δflv1/Δpgr5obtained a single band (~1.5 kb)indicating disruption offlv1. With repeated attempts,an additional band (~2 kb), representing the WT copy offlv1gene, was still detected inflv1/M55 andflv1/M55/Δpgr5(Figure 2B), indicating the existence of the WT copy offlv1in these two mutants.
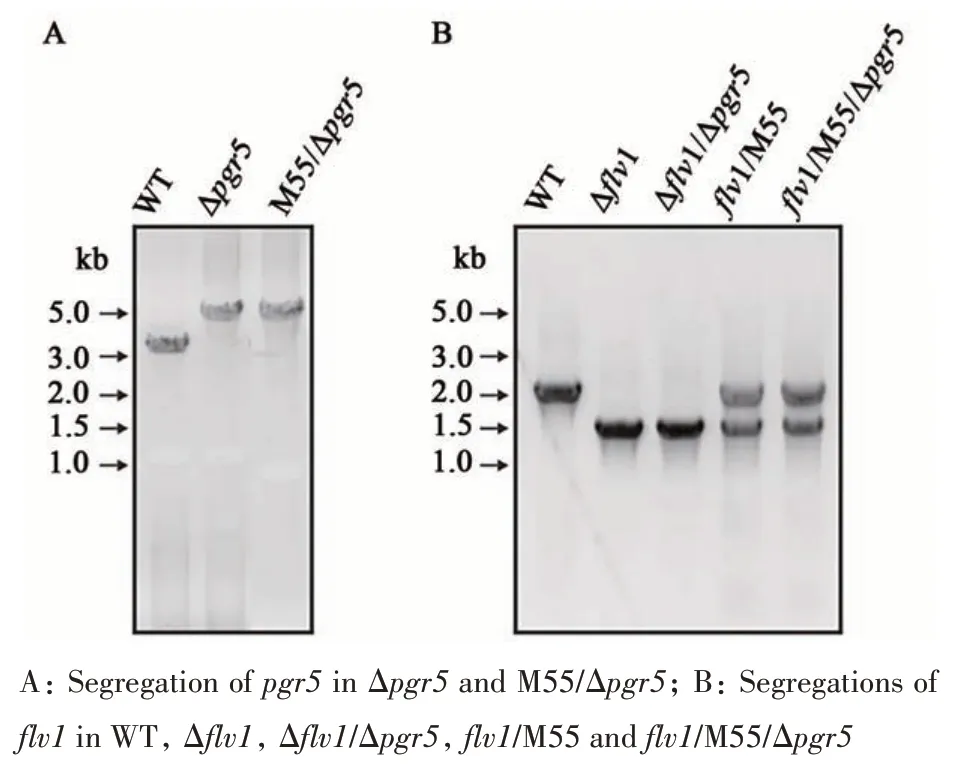
Fig. 2 Verification of pgr5 and flv1 mutants by PCR
The expression offlv1gene inflv1/M55 andflv1/M55/Δpgr5were further analyzed at transcription level. It showed that theflv1transcripts inflv1/M55 andflv1/M55/Δpgr5were decreased to 38% and 49%of that in WT, respectively (Figure 3A). The P700oxidation kinetics showed that the oxidation of Δflv1was extremely slow compared to that of the WT,which was in agreement with previous reports[24]. The relatively slow P700oxidation offlv1/M55 andflv1/M55/Δpgr5indicated that down-regulation offlv1could affect the P700oxidation kinetics, in spite thatflv1was not completely knocked out (Figure 3B).Thus, fourflv1mutants in this study were subjected to further analyses. Severalflv1overexpression strains were also constructed and included in this work, named as WT/PpetJ::flv1, Δflv1/PpetJ::flv1,flv1/M55/PpetJ::flv1, respectively.
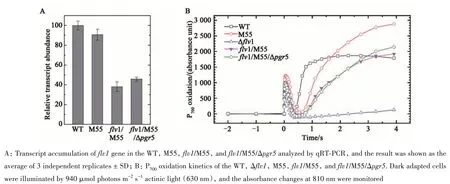
Fig. 3 Characterization of various flv1 mutants
2.2 Growth of WT and mutants under different CO2 and light conditions
To investigate physiological significance of NDH-CET, PGR5-CET and Flv1-mediated Mehlerlike reaction, WT and mutants were grown under different light intensities (growth light, GL; high light, HL), CO2conditions (3% CO2, HC; air level CO2, LC), fluctuating light (FL) and combination light (FLD). Under GL either HC or LC,Δpgr5,Δflv1andΔflv1/Δpgr5grew moderately, similar to the WT,while M55 and M55/Δpgr5grew slower than WT,flv1/M55,flv1/M55/Δpgr5grew very poorly (Figure 4A).Under HL and LC condition,Δpgr5andΔflv1grew similar to the WT, whereas all NDH deficient mutants, including M55, M55/Δpgr5,flv1/M55 andflv1/M55/Δpgr5, could not grow. Supplementation of Na2S2O3promoted growth of all the strains, but it was unable to rescue the growth offlv1/M55 andflv1/M55/Δpgr5under HL condition (Figure 4A). Under FL, Δflv1and Δflv1/Δpgr5were unable to grow,whileflv1/M55 andflv1/M55/Δpgr5were survived,but grew extremely poor, which probably due to the remnant Flv1.Δpgr5was similar to the WT, but growth of M55 and M55/Δpgr5was severely retarded. Overexpression offlv1(Δflv1/PpetJ::flv1)rescuedΔflv1but not to the WT level, whereasflv1/M55/PpetJ::flv1grew similarly toflv1/M55 and M55(Figure 4B). Under FLD,Δpgr5grew well as WT. On the contrary, Δflv1, Δflv1/Δpgr5and M55 exhibited significant retarded growth andflv1/M55 could not even grow (Figure 4C).
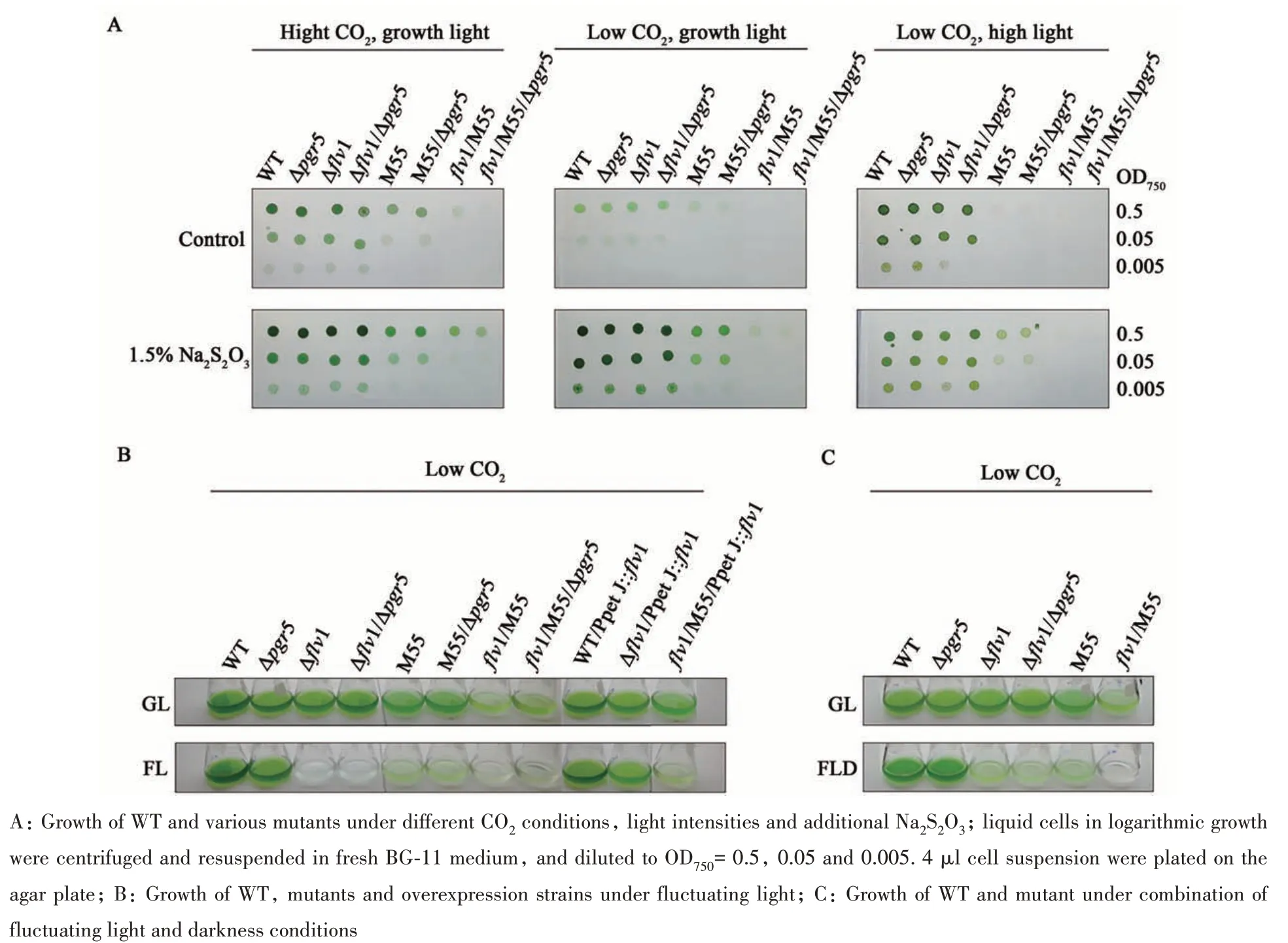
Fig. 4 Growth of WT and mutants under different conditions
2.3 Effects of hydrogen peroxide on the growth of WT and mutants
Since the presence of reducing reagent promoted the growth, the physiology upon oxidative stress was studied. WT,Δpgr5,Δflv1and M55 were grown in the presence of different concentrations of hydrogen peroxide. Figure 5A showed that higher concentration of hydrogen peroxide had more severe negative effects on the growth of all strains. At a low concentration (1 mmol·L-1), the effect of hydrogen peroxide was shown at 16 h but unobservable at 48 h. In the presence of 1.5 mmol·L-1hydrogen peroxide,Δflv1andΔpgr5showed more sensitive to the treatment at 16 h, andthe phenotype at 48 h was not obvious. At a concentration of 2 mmol·L-1,Δflv1andΔpgr5exhibited hypersensitivity, but M55 grew even better than the WT. At a high concentration(3 mmol·L-1), none of tested strains survived(Figure 5A).
Figure 5A showed thatΔflv1showed obvious hypersensitivity when treated with appropriate concentrations of hydrogen peroxide (1.5 and 2.0 mmol·L-1), which might be related to the Mehlerlike reaction. SODs were crucial enzymes involved in Mehler reaction in plants, reducing O2-to H2O2and O2. Likewise, the SOD activities ofSynechocystisWT and mutants were analyzed. Under HC, SOD activities of all strains remained at a relatively low level with insignificant difference. The SOD activities of the WT,Δpgr5,Δflv1and Δflv1/Δpgr5grown under LC increased significantly compared to those in HC condition. Moreover, the SOD activities ofΔpgr5,Δflv1and Δflv1/Δpgr5mutants increased much more than that of the WT in LC. Precisely, the SOD activities ofΔpgr5,Δflv1and Δflv1/Δpgr5under LC were 1.7, 1.7 and 2 times to that of the WT, respectively. Noticeably, the difference of SOD activity was no significant among four NDH deficient mutants (M55, M55/Δpgr5,flv1/M55,flv1/M55/Δpgr5) between the cells grown under LC and HC(Figure 5B).
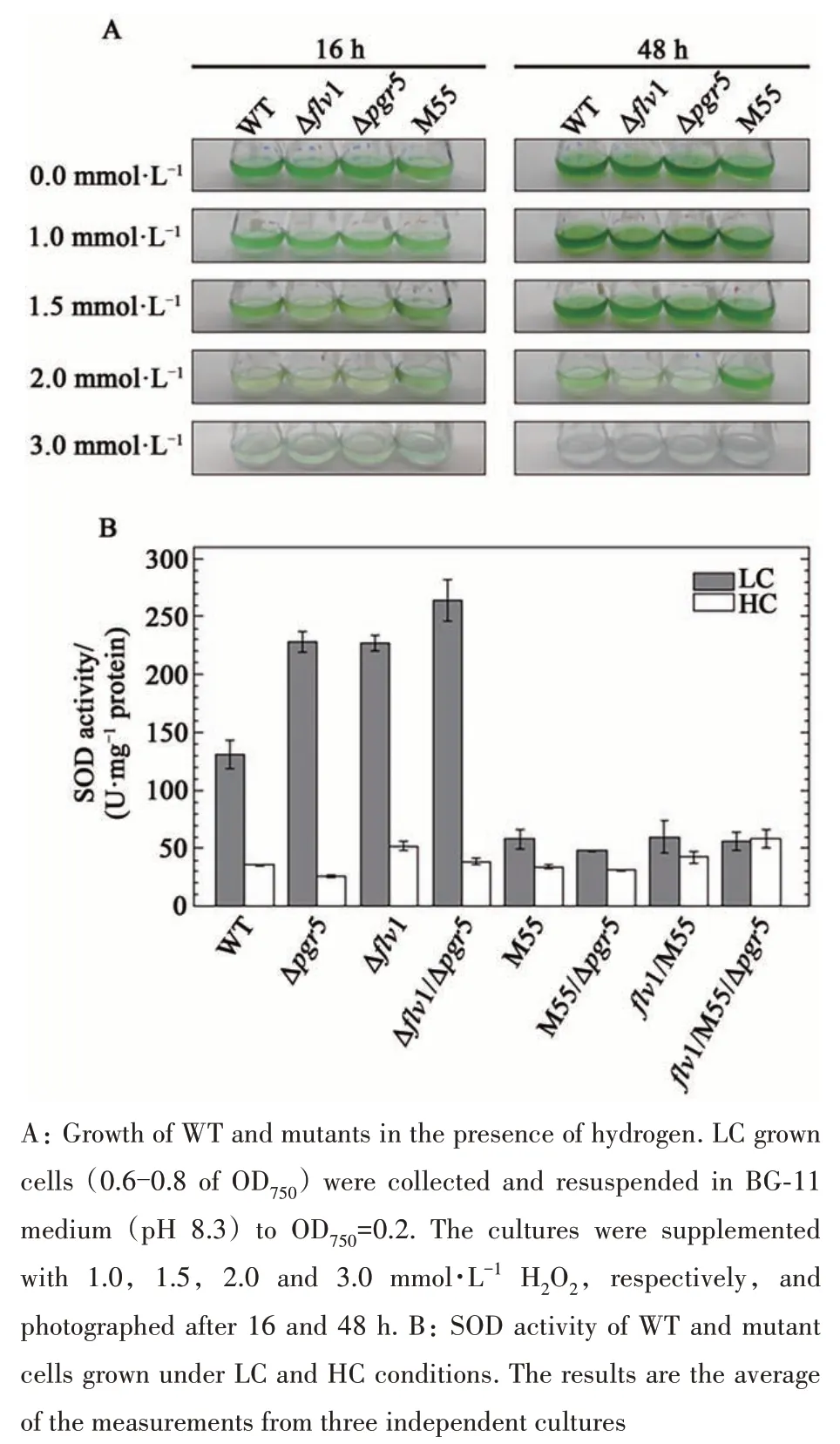
Fig. 5 Hydrogen peroxide treatment and SOD activities of WT and mutants
2.4 Accumulation of photosynthetic proteins in WT and mutants
Since some mutants showed obvious retarded growth (Figure 3 and Figure 4A), it was reasonable to analyze the accumulation of major photosynthetic proteins. In LC, the PSII core subunit, D1, varied little among Δpgr5, Δflv1and Δflv1/Δpgr5, but increased to 135%~175% of WT in four NDH deficient mutants. The PSI subunit, PsaB, varied less than 10% of WT in all mutants. In HC, no significant differences of D1 were detected and PsaB in four NDH deficient mutants was only 15%~25%compared of WT. As expected, an NDH complex subunit, NdhK, was hardly detected in all NDH deficient mutants. The internal reference of a membrane protein, AtpB, varied barely in all strains.Phycobilisome proteins and rubisco are two most abundant proteins in cyanobacteria[39]. In LC, slightly decreases of rubisco large subunit, RbcL, were detected in Δpgr5, M55 and M55/Δpgr5. In HC, the contents of RbcL inflv1/M55 andflv1/M55/Δpgr5were only 30% of that in WT. Ferredoxin: NADPH reductase (FNR), which catalyzes the last step of LET, is an important photosynthetic protein. The large isoform FNRLwas thought to be involved in photosynthetic electron transfer from Fd to NADPH[40]. In LC, no significant differences of FNRLwere detected in all strains. While in HC, the amounts of FNRLinflv1/M55 andflv1/M55/Δpgr5strains were decreased to 45% and 65% of that in WT. More CpcA was expressed in HC than in LC,but there were no significant differences between strains under same growth conditions (Figure 6 and Table 2).
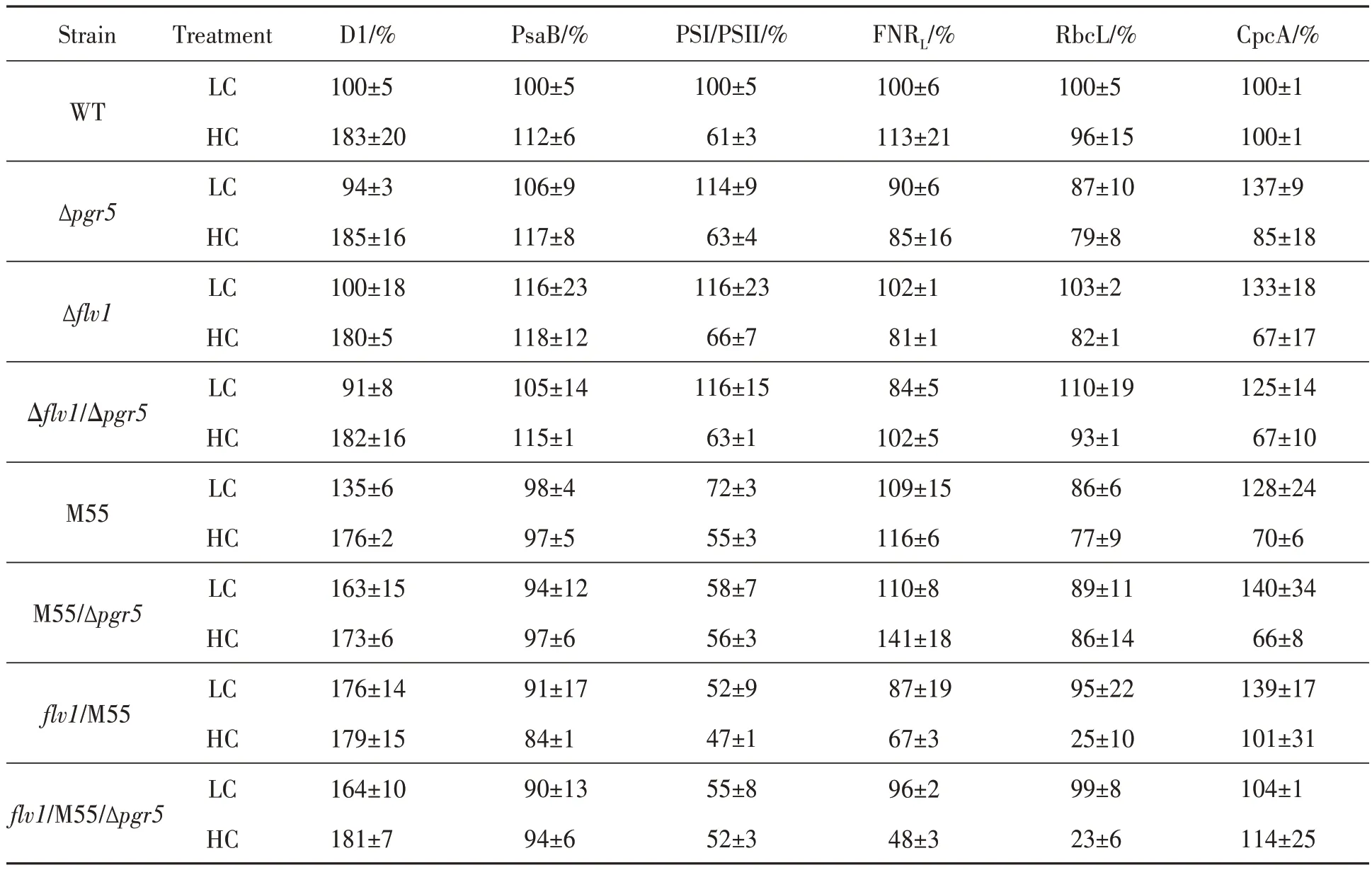
Table 2 Semi-quantification of D1, PsaB, FNR, RbcL and CpcA
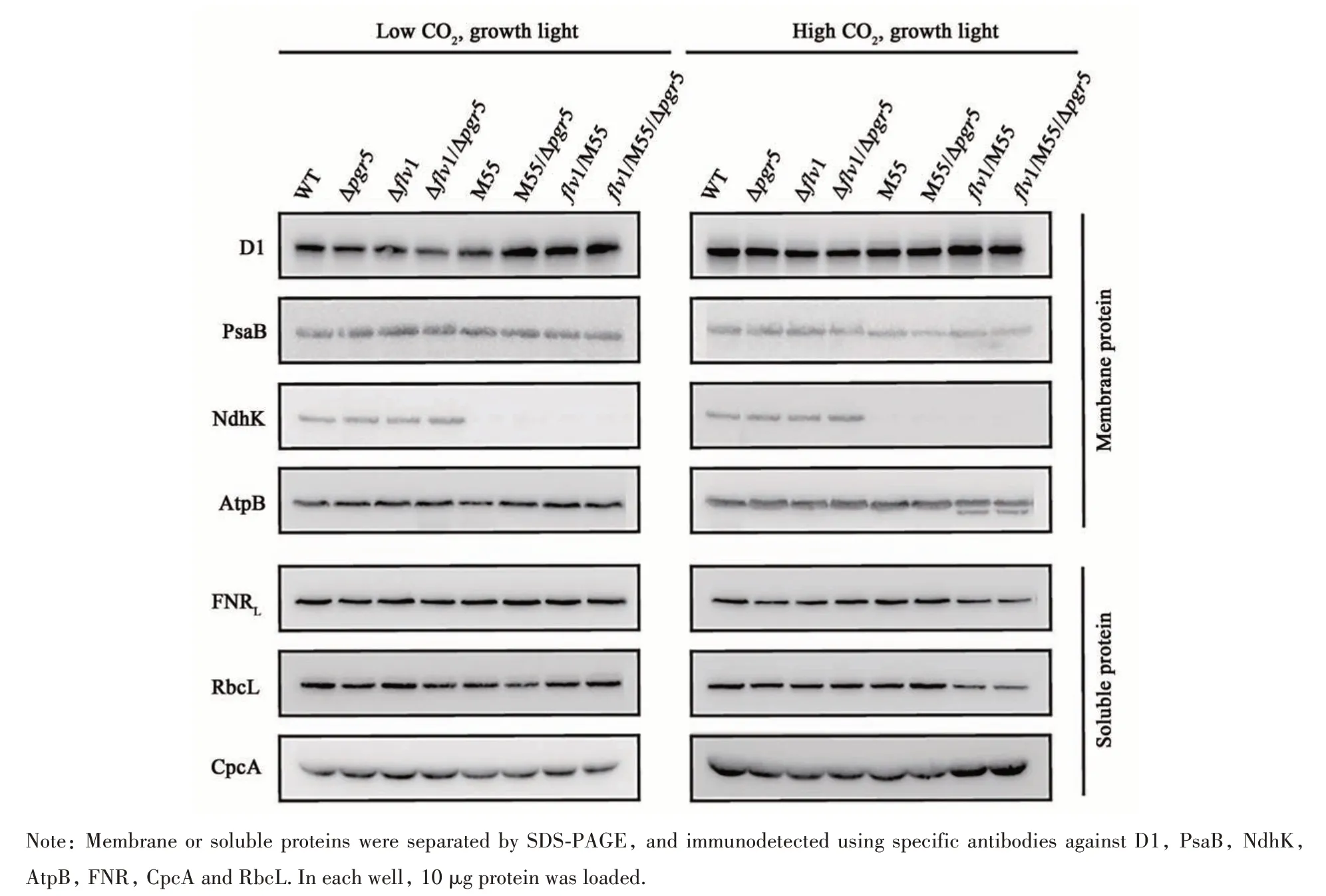
Fig. 6 Expression of major photosynthetic proteins on the thylakoid membranes and in the soluble fraction
2.5 Kinetic analysis of P700+ reduction in WT and mutants
Since mutation sites in this study were involved in several photosynthetic electron transport pathways close to PSI, it was necessary to quantitatively analyze the electron transport rates in these mutants by the measurements of P700oxidation-reduction kinetics. The absorbance changes at 810 nm standardized to 0-1 were shown in Figure 7.
KCin WT represented the electron transfer rate of two distinct CET pathways which consisted of NDH-1 and PGR5 dependent routes. While KCinΔpgr5represented only the NDH-1 dependent route,and that in M55 represented only the PGR5 dependent route. In LC condition, the rate of NDHCET was 99 times higher than PGR5-CET,contributing the majority of the CET. As expected,KCwas near 0 in the CET deficient mutant, M55/Δpgr5(Figure 8A). In HC condition, the NDH-CET was only 9 times higher than PGR5-CET (Figure 8A),probably due to the repression of NDH-13type complex[17]and high expression of PGR5 under HC[41]. In cyanobacteria, four different types of NDH-1 complexes were involved in CET[20]. To further clarify effects of various types of NDH-1 complexes on CET, two mutants, lacking NDH-11,2(ΔD1/D2)and NDH-13,4(ΔD3/D4), were included. Data showed that the sum of KCin ΔD1/D2 and ΔD3/D4 nearly equaled to the KCin Δpgr5.And NDH-11,2dependent sub-route contributed 40% while NDH-13,4dependent sub-route contributed 60% (Figure 8B).
Besides the CET, other electron transfer pathways were also analyzed quantitatively. KFrepresented the rate of Flv1/3 mediated Mehler-like reaction and KNrepresented the rate of electron flow to FNR, thus the sum of these two (KF+KN) represented the efflux rate. The KMrepresented the electron flows from metabolism and showed the electron flow to oxygen via terminal oxidase. In LC, the KF+KNin all NDH deficient mutants were two times more than that in WT. Both in HC and LC conditions, the KMand KRof all NDH deficient mutants decreased significantly(Figure 9B and C).
3 Discussions
3.1 Cyanobacterial NDH-1 complexes contribute major CET and regulate respiratory and metabolic electrons
High light is one of the adverse environmental conditions occurring to most photosynthetic organisms.High-dose irradiation will cause excessive photoelectrons at the acceptor side of PSI and subsequent photo-oxidative damage. The physiological roles of CET have long been underestimated, but recent reports have supported its dominant roles[9]. The CET could recycle those excessive photo-electrons from ferredoxin back to the PQ pool generating more ATP for CO2assimilation[12,42]. In this work, the CET deficient mutant, M55/Δpgr5, died under HL,further demonstrating that the CET is essential under high light stress (Figure 4A). Two independent pathways so far were identified to be involved in the CET around PSI in photosynthetic organisms. One is mediated by NDH-1 complexes and the other one is involved with PGR5 and PGRL1[11-12,42-43]. With a recent finding of functional analogue of PGRL1 inSynechocystisand limited contributions of PGR5[13],the CET in cyanobacteria was thought to mainly depend on NDH-1 complexes[19,44]. Our data not only supported this assumption, but also presented accurate quantification of these two routes. All NDH defect mutants (M55, M55/Δpgr5,flv1/M55,flv1/M55/Δpgr5) were unable to acclimate under HL.And the growth rate of the NDH-1 deficient mutants,M55 and M55/Δpgr5was similar under our experimental conditions (Figure 4A). Further quantitative analysis of P700kinetics of M55, Δpgr5and M55/Δpgr5showed that the NDH-CET took 99% and 90% of the total CET activity under LC and HC, respectively(Figure 7 and Figure 8).
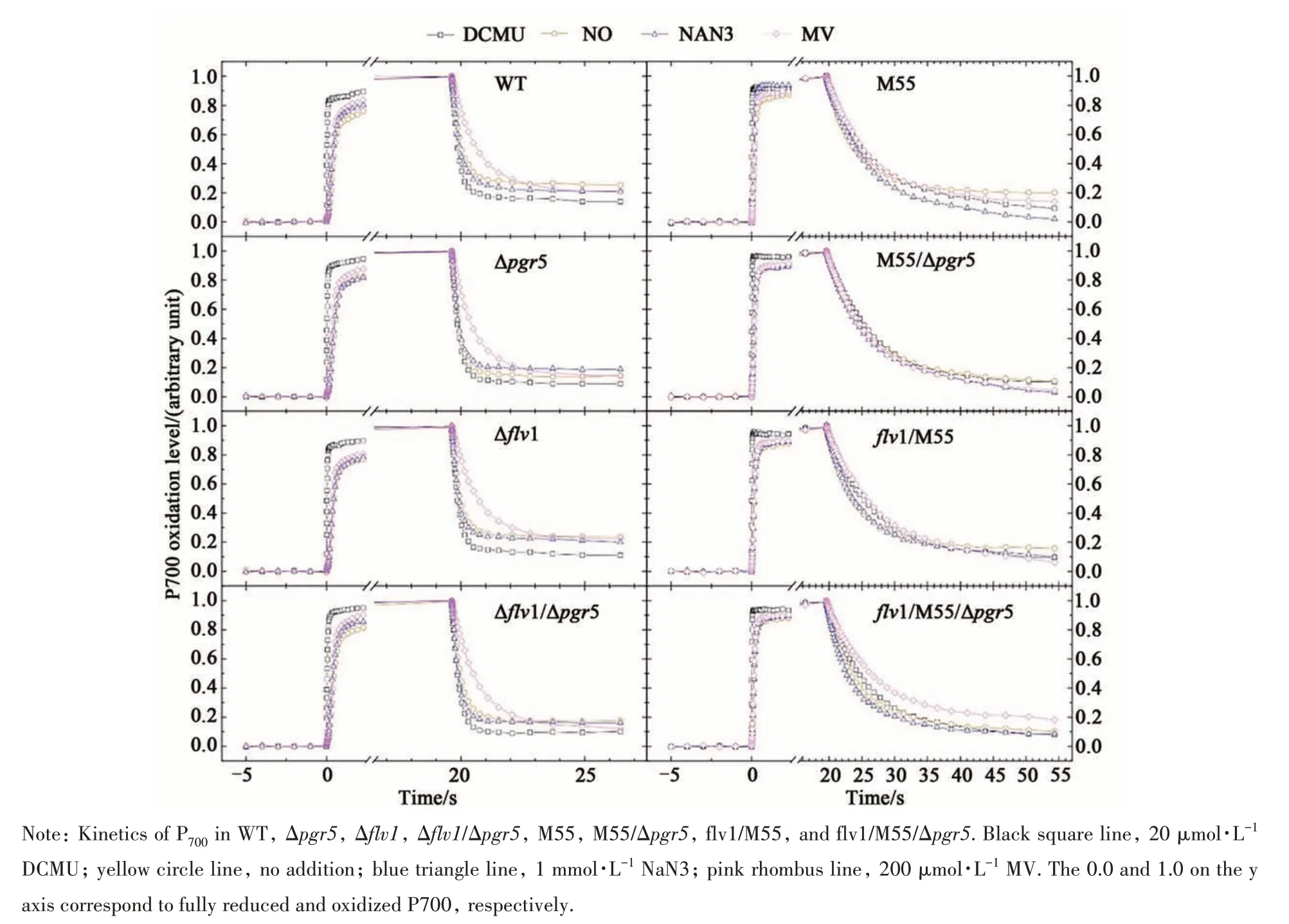
Fig. 7 Kinetic analysis of P700+ reduction in WT and mutants
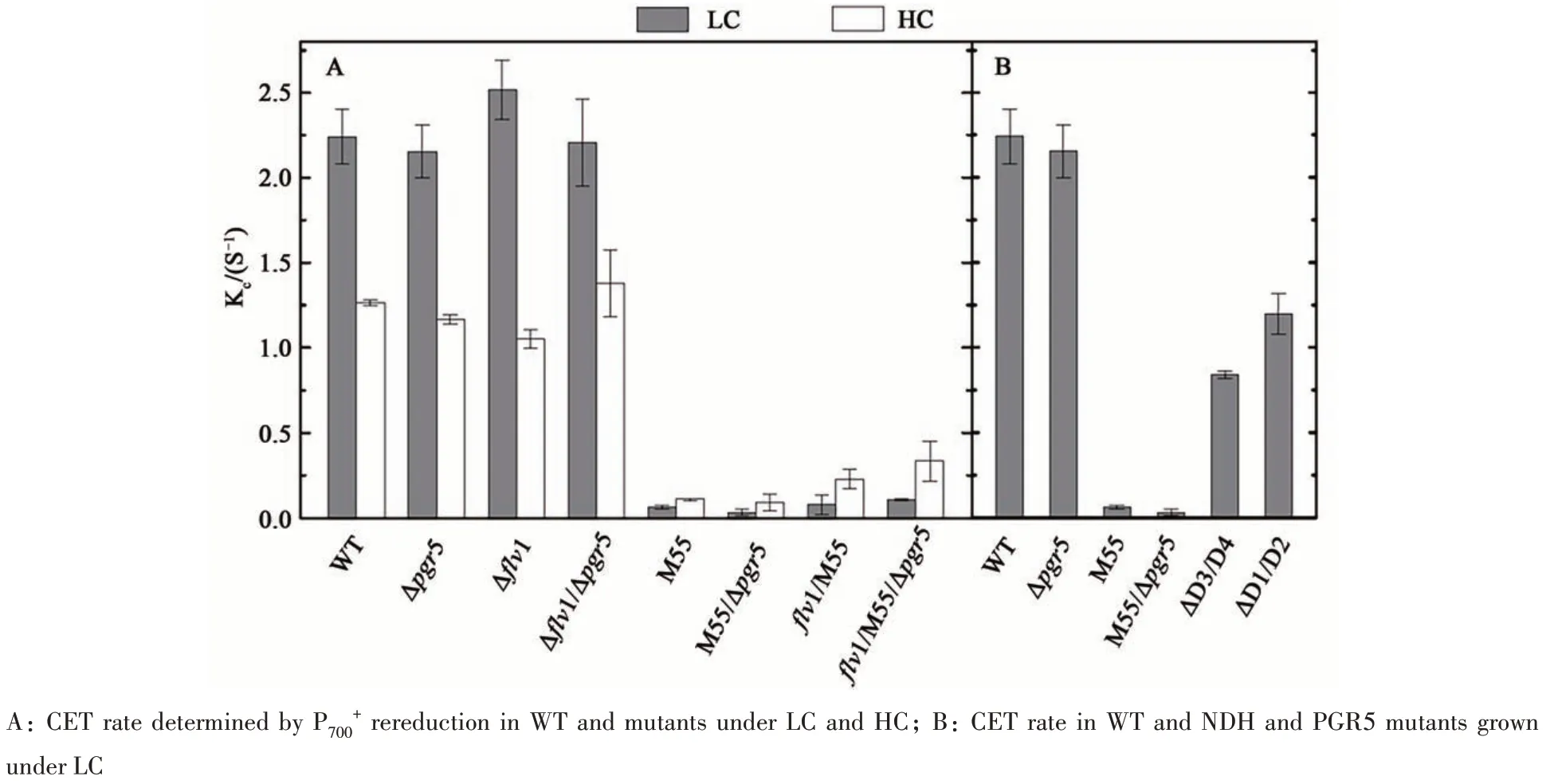
Fig. 8 Cyclic electron flow in WT and mutants
Besides CET, NDH-1 complexes have long been thought as a component shared with distinct electron transfer networks in cyanobacteria, including electron transfer of photosynthetic, metabolic and respiratory pathways[16,19]. Previous work by Bernat and colleagues[19]suggested that the NDH-1 complexes possessed a high capacity for electron donation from metabolites to PQ. Our data of P700analysis further supported this assumption. In allndhB(M55)inactivation mutants, the electron flow rates from metabolites (KM) were only 1/6 and 1/3 of that of the WT in LC and HC, respectively (Figure 7 and Figure 9B). The low KMvalues of all NDH-deficient mutants demonstrated that NDH-1 complexes are important for most electron donation from metabolites to the PQ pool in cells grown under our investigated conditions. Several recent studies have resolved thestructure of NDH-1 complex, which further revealed the basis of enzyme mediated the major CET route in cyanobacteria. It has clarified that NDH-1 complexes are also the member of FQR (ferredoxin quinone oxidoreductase), the same as PGR5/PGRL1. In cyanobacteria, photosynthesis and respiration share some key components, such as the PQ pool and cytochromeb6f. It has been suggested that NDH-1 complexes participated in respiratory electron transfer[17]. However, Cooley et al.[45]have also discovered electron flows via succinate dehydrogenase(SDH) to PQ, indicating SDH may have a bigger impact on respiratory electron transfer than NDH-1.Nevertheless, the KRvalues, in our tested conditions,supported NDH-1 as the major player for respiration.The KRvalues of all NDH-deficient mutants were near to 0 under LC and HC, indicating that NDH-1 complexes may have dominant roles in respiratory electron transfer (Figure 7 and Figure 9C).
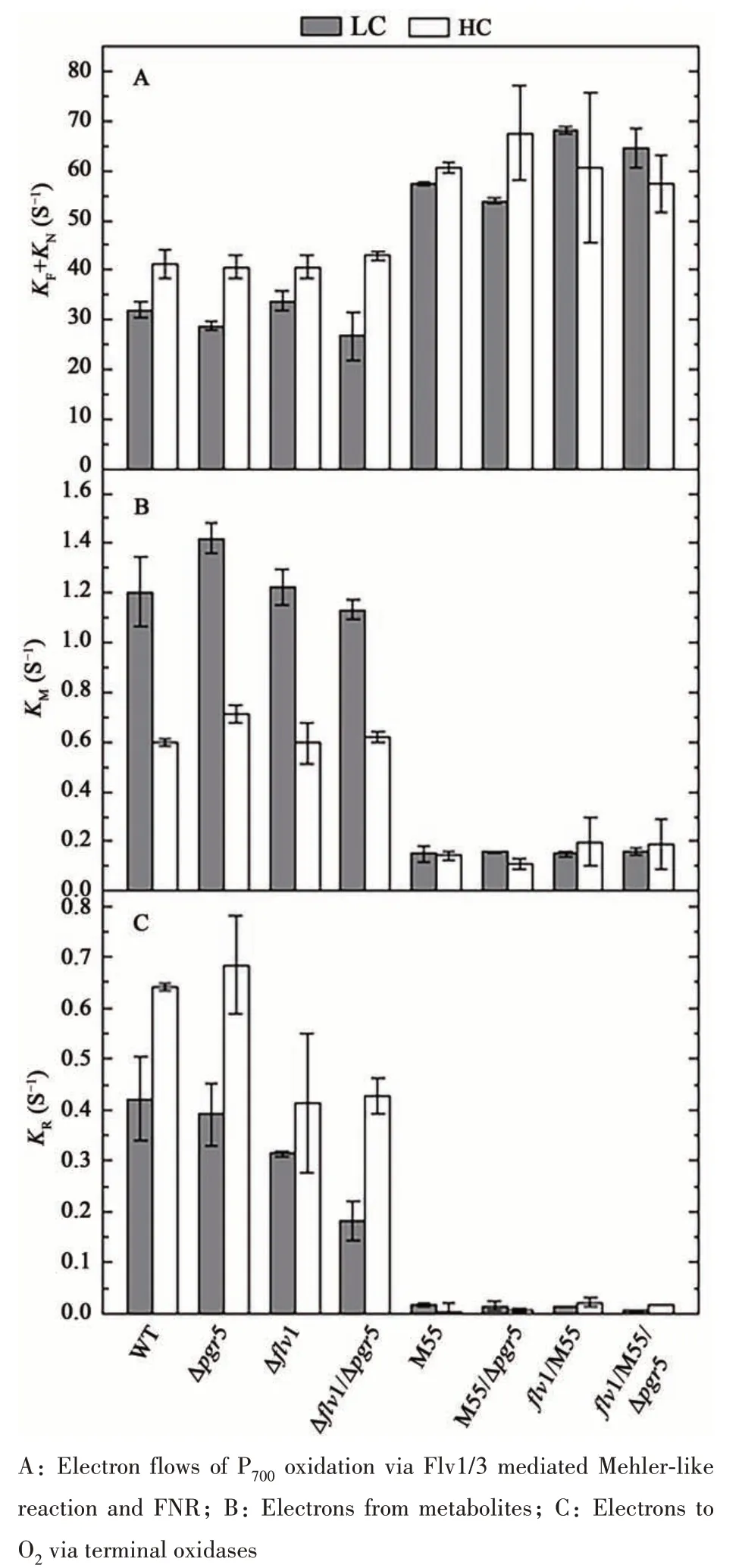
Fig. 9 Electron flows around P700 and PQ pool in WT and mutants under LC and HC
3.2 PGR5 route has limited contribution to CET in cyanobacteria
In higher plants, the dominant CET was mediated by PGR5/PGRL1[12,43]. An ortholog with low homology of PGR5 involved in an antimycin A-sensitive electron transfer was identified in cyanobacteria[13]. However, the physiological role of PGR5 route in cyanobacteria was unclear until now.Our data provide some hints to its physiological significance and further supported the presumption that PGR5-CET was far less important in cyanobacteria. Addition of hydrogen peroxide would result in oxidative stress in cyanobacteria. Negative effects of hydrogen peroxide (2 mmol·L-1) on growth were much more evident on Δpgr5(Figure 5A). This observation indicated the probability of participation of PGR5 in the responses to oxidative stress in cyanobacteria. SOD plays protective roles catalyzing a disproportionation of O-2to offset the damage effects of oxidant[7]. From this perspective, the endogenous SOD activity could be considered as an index to reflect the oxidative stress level. The relatively higher SOD activity of Δpgr5under LC indicated the higher oxidative stress in this mutant and the involvement of PGR5 against oxidative stress (Figure 5B). The KCvalues of Δpgr5were only slightly smaller than that of the WT under LC and HC, and KCvalue of CET deficient double mutant(M55/Δpgr5) was almost same as that of NDH-CET deficient mutant (M55) under LC and HC, indicating that the contribution of PGR5 route to CET was limited (Figure 7 and Figure 8A). All the data in our experiments demonstrated that, in cyanobacteria,PGR5-CET might play a physiological role against oxidative stress, although its contribution compared to NDH-CET was rather small.
3.3 Mehler-like reaction functions in a rapid and protein amount independent mode
Previous work by Helman et al.[24]firstly provided evidence for the involvement ofSynechocystisFlv1 and Flv3 in Mehler-like reaction, directly reducing oxygen to water without production of any ROS. However, no observable phenotypes of Δflv1and Δflv3mutants were found. Later, Zhang et al.[36]found that Δflv1and Δflv3mutants did not show negative effects on growth under steady-state light conditions (either low, medium or high light). Similar observations were also showed in this study (Figure 4A). The physiological role of Flv1/Flv3 involved Mehler-like reaction in cyanobacteria was established until the indispensability of Flv1 and Flv3 inSynechocystisunder fluctuating light[25]. Likewise, we observed the Δflv1incapable of acclimation under FL and severely retarded growth under FLD (Figure 4B and C), while under steady-state light (either GL,LL or HL),flv-deficiency did not cause any negative effects on growth (Figure 4A)[36]. These suggested that Flv-dependent Mehler-like reaction is crucial during rapid change light regime. Considering that HL phase was less than a minute during the fluctuating light cycle, the activation of Flv mediated Mehlerlike reaction ought to be an extremely rapid response.Such quick reaction could not depend on any protein expression or accumulation. Firstly, we have thought the transcript levels offlv1andflv3might be the key. Since the transcript level offlv3was much higher thanflv1inSynechocystis[36], we constructed anflv1overexpression strain driven by a promoter ofpetJ,which was suggested to be strong in cyanobacteria[32].There were no observable phenotypic differences between the WT and WT/PpetJ::flv1under FL(Figure 4B), indicating that the amount of Flv1 in WT was sufficient. Hence, we speculated that the real efficiency of Mehler-like reaction in cyanobacteria might be independent of the amount of expressed protein, but dependent on the amount of Flv1/Flv3 actively involved in the reaction. The Δflv1and Δflv3mutants showed severe defect in quick oxidation of P700when actinic light excited both PSI and PSII (Figure 3B)[24]. However, such phenotype disappeared when DCMU was supplemented or excitation by far-red light (Figure 7). We assumed that Flv1/Flv3 mediated Mehler-like reaction could be activated only at the condition of high electron flux from PSII to PSI, resulting a strong reducing pressure at acceptor side of PSI. This assumption would explain the KF+KNvalues in Δflv1and Δflv1/Δpgr5were similar to that in WT (Figure 7;Figure 9A).
3.4 Cooperation of CET, Mehler-like reaction and SOD during different HL stresses
Besides linear electron transport, electrons at acceptor side of PSI have three alternative routes to flow away in cyanobacteria. The CET recycles electrons back to the PQ pool, keeping electrons flowing in the photosynthetic electron pathway. The Mehler-like reaction and SOD are two “parallel” routes,leading electrons “escaping” from photosynthetic electron transport. The expression of NDH-1 complexes has been reported to be dynamically in response to environmental changes[46]. Previous studies have shown that the activity of NDH-1 largely relied on the accumulation of NDH-1 complexes. Recently, a report showed that NdhO subunit negatively regulated the stability of NDH-1M and CET activity[47]. Such regulation might be able to quickly alter the NDH-1 activity to a certain extent. Nevertheless, extensively increasing activity of NDH-1 requires protein synthesis and assembly,which were in hours’ time scale. Consequently, it should take some time to fully activate the NDHCET in cyanobacteria. Compared to the observation mentioned above that Flv1/3 rapidly and directly reduce molecular oxygen to water with no concomitant ROS production in less a minute, reduction of oxygen to water by antioxidant enzymes is multi-step and hence relatively time-consuming[7]. In cyanobacteria,SOD is the first antioxidant enzyme to alleviate oxidative stress by disproportionation of O-2to H2O2.Subsequently, the hydrogen peroxide could be reduced to water by catalases and peroxidases or be served as photosensitizers of signaling other responses in cyanobacteria[48-49]. According to our data, we postulated a hypothesis on how CET,Mehler-like reaction and antioxidant enzyme system cooperated under constant low and high light as well as FL conditions (Figure 10).
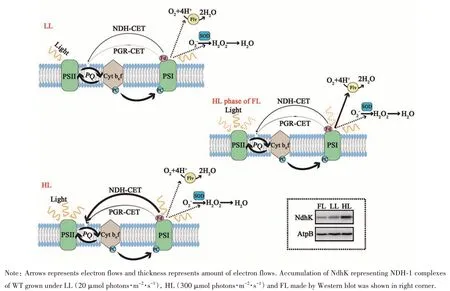
Fig. 10 Working scheme of CET, Mehler-like reaction and SOD in cyanobacteria
Under constant light conditions, Flv activity was limited, most excessive electrons pass through CET (major NDH) and the SOD was a minor route.That was why M55 showed severely retarded growth under HL, whereas Δflv1mutant grew similarly to WT (Figure 4A). When NDH-CET, the dominant electron pathway, was missing, high excitation pressure shown as high value of KF+KNwas observed in all NDH-deficient mutants (Figure 7 and Figure 9A),thus part of electrons was forced to the Flv route.This assumption might explain overexpression offlv1inflv1/M55 promoted the growth under growth light(Figure 4B). Unsuccessfully knockingflv1out in M55 by our attempts also suggested that M55 was highly dependent on the electron route of Mehlerlike reaction. Activation of Mehler-like reaction in M55 under constant light condition may also explain the observation that the M55 grew better than WT in the presence of hydrogen peroxide (Figure 5A).Under constant light condition, the NDH-CET is the dominant pathway inSynechocystis, while the Mehler-like reaction is not activated (Figure 10). For M55, the NDH-CET is blocked and the SOD activity is very low (Figure 5B), but the FLV-route is partially activated. Supplementation of proper concentration of hydrogen peroxide provided a moderate oxidative stress. We assumed that the WT did not activate FLV-route under this condition, and its detoxification probably was via activation of antioxidant enzymes. Since activated Mehler-like reaction in M55 was much efficient than the antioxidant enzyme system in WT, it would be possible that M55 was better able to cope with hydrogen peroxide (Figure 5A).
Deletion of PGR5 had no visible phenotype in all tested conditions (Figure 4) except higher oxidative stress and SOD activity upon hydrogen peroxide treatment (Figure 5). We assumed that minor defect of CET (PGR5 route) would not switch on the Flv route, but rather activate antioxidant enzyme system. Under FL condition, accumulation of NDH-1 complexes was largely dependent on the low light phase (Figure 10). During rapid high light period, the amount of existing NDH-1 complexes was not enough and substantial excessive electrons were accumulated at PSI acceptor side. Since there was barely time for accumulating more NDH-1 complexes, quickly activated Mehler-like reaction was crucial to releasing the extra electrons.Therefore,flv1deletion mutants (Δflv1and Δflv1/Δpgr5) andflv1knock-down mutants (flv1/M55 andflv1/M55/Δpgr5) were extremely sensitive to FL(Figure 4B). Since activation of Flv-mediated Mehler-like reaction was due to inadequate of electrons redirected by NDH-1 complexes, we hypothesized that NDH-1 mediated CET was the major route robustly supporting HL acclimation,while Flv1/3 mediated Mehler-like reaction was a redundancy route operating in case of insufficiency of the main electron path. This presumption could also explain the observation thatSynechocystisflv1andflv3mutants were more miserable under FL20/500 than FL50/500[25].
The Flv-mediated Mehler-like reaction reduced molecular oxygen back to water[23], which could be thought as low efficiency and high energy-consuming in cyanobacteria. Despite all that, we still believed that the Mehler-like reaction, as a redundancy route for NDH-CET, was indispensable and irreplaceable in cyanobacteria. The Mehler-like reaction was a big electron sink functioning as a safety valve for optimal photosynthesis inSynechocystis, which was partially similar to findings that Flv-route was a powerful electron sink inSynechocystisbut not inSynechococcus[50]. Minor deficient in CET (e.g.Δpgr5) could not direct electrons to Mehler-like reaction, suggesting the threshold of Flv-route activation was probably high. Such tightly controlling the activation conditions of Flv-route was important to not waste photosynthetic electrons during moderate stresses. The property of fast response of Flv route might provide cyanobacteria ability to quickly acclimate in rapid environmental changes and adapt to constantly changing environments.
In our study, the effects of SOD functioning as an alternative photosynthetic electron sink were limited. Recent report suggested an essential role of SOD inSynechocystisbased on the observation that the uniquesodBgene could not be knocked out and severe phenotypes were found when the expression ofsodBwas shut down using a Cu2+induced promoter[29]. We thinked thatSynechocystisSOD might physiological significant on redox regulation and signaling, however its contribution to dissipation of excessive electrons was small.
3.5 Implications of CET and Mehler reaction coevolution from cyanobacteria to higher plants
In cyanobacteria, NDH-1 complexes were involved in various important physiological responses, including respiration, carbon concentrating mechanism (CCM)and the CET, and NDH-CET is the dominant CET route[16-17]. This study observed that PGR5 deficient mutants showed no obvious phenotypes, which further demonstrated its minor roles in cyanobacteria(Figure 4). In plants, PGR5 played dominant roles[12].PGR5 regulated the proton gradient, which was highly associated to inductions of the nonphotochemical quenching and the xanthophyll cycle[15,51]. However, the NDH-CET was no longer the main CET route and the chloroplast NDH complex did not participate in respiration and CCM either.The insignificant role of NDH complex was probably due to the separation of plant respiration into mitochondria and evolution of more complicated C4 photosynthesis to replace the CCM function in plant.In mossPhyscomitrella patens, only PGR5/PGRL1-CET and Flv-mediated Mehler-like reaction were reported to function in photosynthetic electron transport and PGR5/PGRL1-CET was the dominant CET[52]. From evolutionary perspective, moss was in the intermediate species between cyanobacteria and higher plants. Development of PGR5-route could be another reason to make NDH-dependent CET unimportant gradually. InArabidopsis, the PGR5 deficient mutants were sensitive to FL indicating its partial replacement the function of Flv[53]. Meanwhile,plants had evolved more complicated antioxidant enzyme systems, which were able to both balance signaling and scavenge excessive ROS efficiently.The scavenging of ROS by antioxidant enzyme system in plants was far more complicated and energy economically than Mehler-like reaction in cyanobacteria. Hence, we speculated that, as the evolution of PGR5-route and antioxidant enzyme system, major functions of NDH-1 complexes were replaced by PGR5 and Flv-mediated Mehler-like reaction finally disappeared in angiosperm plants.
Acknowledgements
We thank Prof. Weimin Ma from Shanghai Normal University for NdhK antibody, Prof. Wendy M. Schluchter from the University of New Orleans for CpcA antibody, and Prof. Ghada Ajlani from Centre National de la Recherche for FNR antibody.
