中国新疆地衣新记录*
2023-05-16热衣木马木提范胜男图尔荪阿依排祖拉
热衣木·马木提,范胜男,图尔荪阿依·排祖拉
(新疆大学 生命科学与技术学院,新疆 乌鲁木齐 830017)
0 引言
形成地衣或地衣化是真菌的主要进化方式之一.地衣作为一种典型的共生生物,由于藻类和真菌长期紧密地结合在一起,无论在形态上、结构上、生理上和遗传上均形成一个单独固定的有机体.分子钟研究表明,地衣最早出现在二叠纪和三叠纪,大约300~200 mya,其中有些大型地衣,如梅衣科地衣Parmeliaceae在白垩纪晚期就出现了[1-4].早期,魏江春明确地衣的概念为地球生物圈内生态系统中地衣型真菌作为建群种(constructive species)与相应的藻类或蓝细菌作为伴生种(companion species)结成稳定的胞外共生生命的支撑系统,即菌藻共生群落[5].它们在地球生物圈内的系统地位则分别是藻类和真菌的组成部分.最近,有学者通过多种地衣基因组和荧光原位杂交技术研究发现:地衣不只是由一种子囊菌和一种藻类组成,存在第三种共生真菌(担子菌)参与的现象[6],这一发现打破了140年来普遍被人们所接受的地衣是一种真菌(通常是子囊菌)与光合作用伙伴之间的共生关系的这一概念.现有地衣物种总数为19 387种,隶属8纲、39目、115科、995属,占已知真菌110 000种的17%、已知子囊菌种的27%[7-8].地衣中共生的担子菌门数量较少,共172种,占总数的0.9%,隶属1纲、5目、5科、15属[9],与其共生的藻类为绿藻和蓝藻,共20余属.我国已报道的地衣物种总数为3 085种,隶属真菌界的2门、10纲、28目、99科、445属[10].地衣作为自然界的先锋生物,在环境指示和检测中发挥着重要作用,在食用、染料、化学、医学、香料、测年、地质探矿等方面也具有较高的应用价值.随着科学技术的不断发展,地衣资源的应用范围将会日益扩大.对地衣型真菌的生物多样性研究,无论是在科学领域还是实践领域均具有重要意义.
被誉为“原始森林”的巩乃斯林场位于中国新疆天山西部巴音郭楞蒙古自治州和静县,具有7.853 3万公顷天然林资源,海拔在1 600~2 400 m之间.2003年,被国家林业局批准为国家级森林公园.巩乃斯林场地处天山腹地,属大陆高寒山地气候,冬季漫长、寒冷多雪,夏季较短、雨量充沛,多东南风.向西开口的喇叭地形拦截了到达新疆上空的大西洋和北冰洋等较湿润的大气环流,形成一定的降水,植被生长发育繁茂,宛如是突出在整个新疆干旱戈壁荒漠大平原上的一座庞大的“绿色岛屿”.2019年10月18日,巩乃斯林场入选“中国森林氧吧”榜单.与生态地理环境多样性相适应,植被类型也表现出了极大的丰富性,基本为多年生中生型和湿生型.地质构造属天山纬向褶皱构造体系,由一系列的东西向复式褶皱和断裂、北东向和北西向的扭性断裂、东西主追踪断裂和南北向张性断裂等和所夹岩块组成.构造形迹无论是褶皱还是断裂,均严格按照一定的方向、部位、顺序和组合关系分布,各个形迹之间都依次有着生成联系.其特有的地理位置、气候、植被和生物资源,形成了特殊的类型,是自然馈赠给人类的宝贵财富,具很高的科研和经济价值.经考察发现,该森林内以死树或朽木作为栖息地的附生植物的地衣种类多,且地衣多样性往往高于活树上生长的地衣种类,因此也成为中国濒危地衣研究的一个重要地区.
2013―2021年,对巩乃斯林场进行地衣资源调查过程中发现了一个新疆新记录属:四胞极衣属Tetramelas Norman.和五个新记录种:多变橙衣Caloplaca variabilis (Pers.) M¨ull.Arg、旱生野粮衣Circinaria arida Owe-Larss、祁连鸡皮衣Pertusaria qilianensis Ren Q & Zhao ZT、肉饼干衣Rinodina sarcogynoides (H.Magn.) Q.Y.Zhong & Li S.Wang和绿色四孢黑瘤衣Tetramelas chloroleucus (Korb.) A.Nordin.研究结果丰富了新疆地衣的物种多样性,为我国地衣物种区系研究提供了基础资料.
1 研究方法
1.1 形态解剖学与生物化学研究
标本外部形态特征使用舜宇体视显微镜(SMZ45-ST1)和全自动数码生物显微镜(Ni-E)进行观察、拍照.详细记录特征,包括地衣体生长型、颜色、附属结构和子囊盘大小、形状、盘面及边缘颜色、完整性等;徒手切片法制作子囊盘纵切临时切片,使用研究级倒置荧光显微镜(Ti-E)和尼康E100生物显微镜观察子囊盘纵切显微结构并拍照.化学成分研究采用显色反应(TC)(常用试剂有K:KOH (10%~25%的氢氧化钾);C:Ca(ClO)2(次氯酸钙饱和水溶液);KC:氢氧化钾和次氯酸钙(先使用氢氧化钾,紧接着补加次氯酸钙);P:对苯二胺的乙醇溶液;I-KI:碘-碘化钾等)、薄层色谱(TLC)等方法[11].
1.2 分子系统学研究
使用Solarbio真菌基因组DNA提取试剂盒进行DNA提取,选取rDNA-ITS位点聚合酶链式反应(PCR)扩增,引物选择ITS 1和ITS 4[12-13].反应产物送往山东博迈达生物科技有限公司测序.原始序列通过DNA STARSeqman整理拼接,提交至基因数据库(GenBank).选择NCBI比对结果相似性高以及与自测序列相关的物种序列,详细信息见表1.通过最大似然分析(ML)和贝叶斯分析(BI)构建五个新记录种的系统发育树.使用MEGA 6.0比对(alignment)序列,手动修剪序列矩阵,MrModel Test选择最佳模型,基于最大似然分析和贝叶斯分析评估系统发育树各分支[14-16].
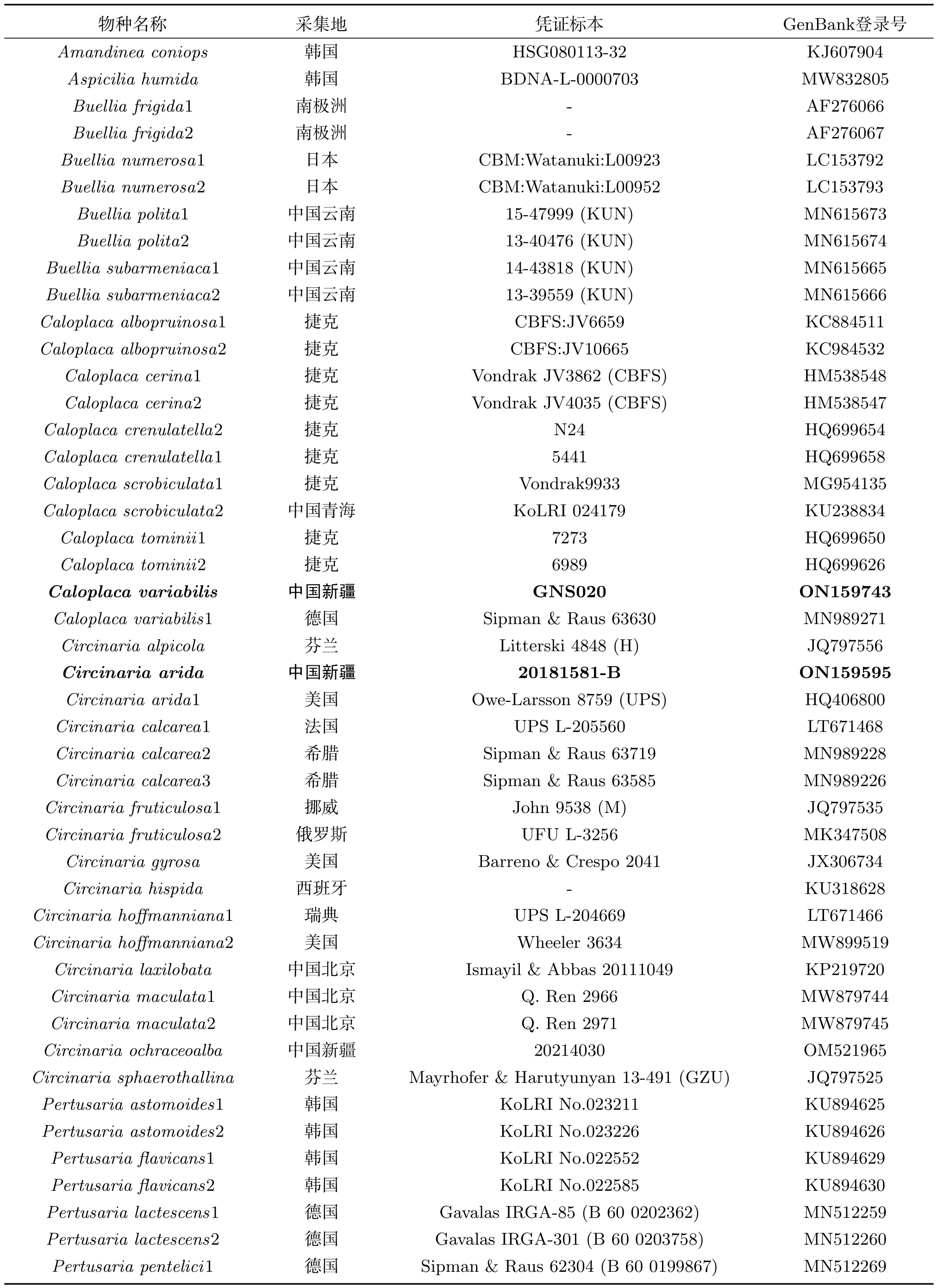
表1 用于系统发育分析的物种及序列信息
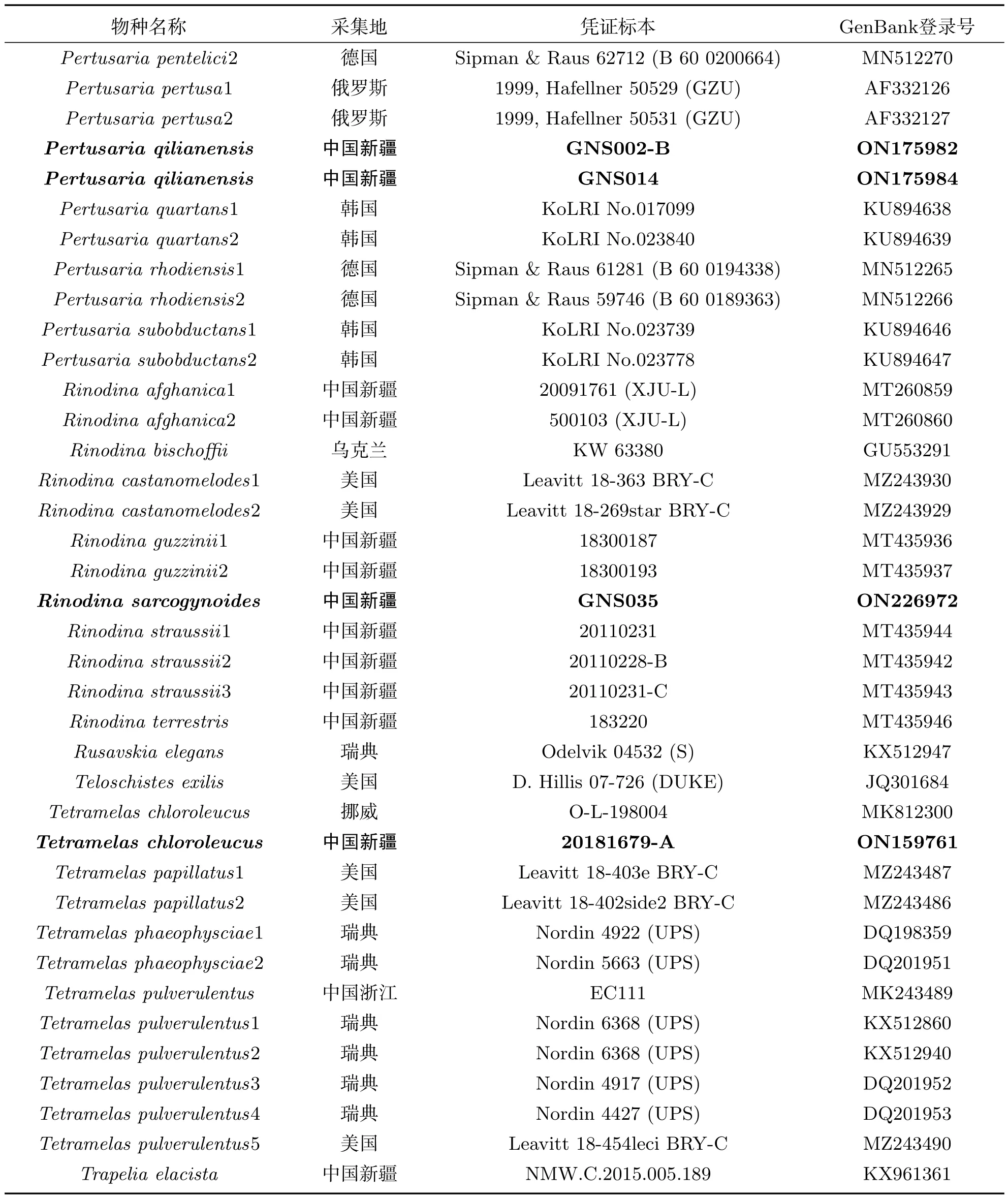
续表1
2 结果与讨论
2.1 系统发育分析
2.1.1 多变橙衣系统发育分析
参与构建多变橙衣ITS系统发育树的序列共14条,以细黄枝衣Teloschistes exilis (Michx.) Vain为外源类群.结果显示系统发育树分为两个主要分支(图1),第一个分支中,自测序列与多变橙衣其它序列聚成一单系群(支持率BI=70),与Caloplaca albopruinosa构成一复系群,亲缘关系较近.
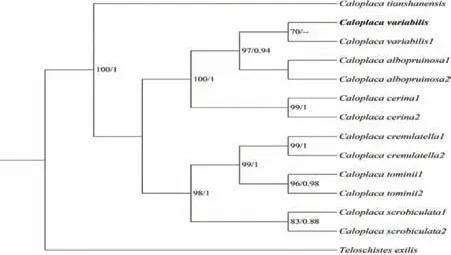
图1 多变橙衣ITS序列系统发育树
2.1.2 旱生野粮衣系统发育分析
旱生野粮衣ITS位点测序产物在NCBI中进行Blast比对分析,结果显示自测序列与GenBank登录号为HQ406800的旱生野粮衣序列相似性极高,达到100%.共使用18条序列构建旱生野粮衣的系统发育树,以Aspicilia humida为外源类群.由图2可知,进化树分为两大分支,旱生野粮衣在第二分支中种间进化关系明确,两个旱生野粮衣物种聚成一单系群(BI=100,后验概率PP=1),且与果野粮衣(Circinaria fruticulosa)亲缘关系最近.
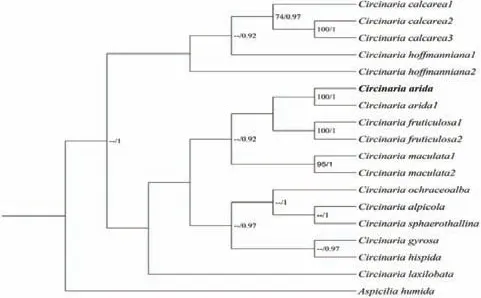
图2 旱生野粮衣ITS序列系统发育树
2.1.3 祁连鸡皮衣系统发育分析
截至目前,祁连鸡皮衣还未有相关分子数据,因此将自测序列在NCBI中Blast比较后未找到关系密切序列.以Trapelia elacista为外源类群,构建祁连鸡皮衣的ITS系统发育树.由图3可知,本文自测的两条祁连鸡皮衣序列聚成一个独立单系群(BI=86,PP=1),很好地汇聚在鸡皮衣属的系统发育分支中,且与南方鸡皮衣(Pertusaria rhodiensis)形成具有良好支持的姊妹群(BI=100,PP=1).
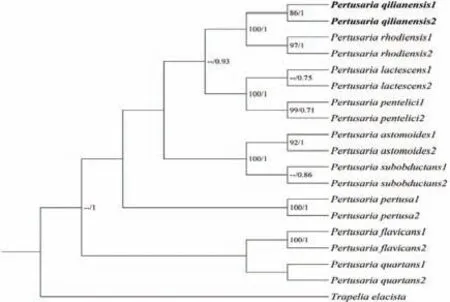
图3 祁连鸡皮衣ITS序列系统发育树
2.1.4 肉饼干衣系统发育分析
将自测序列与NCBI中下载的12条序列构建ITS位点的系统发育树,以丽黄磷衣(Rusavskia elegans)为外源类群.由图4可知,肉饼干衣的单条序列形成一个独立分支,与类栗饼干衣(Rinodina castanomelodes)亲缘关系最近,两物种聚成一并系分支(BI=79,PP=0.91),但两种外部形态差异明显,因此作为两个物种处理.
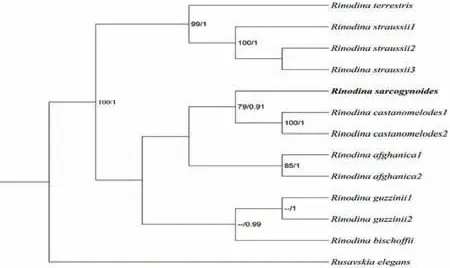
图4 肉饼干衣ITS序列系统发育树
2.1.5 绿色四胞极衣系统发育分析
将绿色四胞极衣自测序列在NCBI中Blast比对,结果显示与Genbank登录号为MK812300的绿色四胞极衣密切相关,达到99%,以Amandinea coniops为外源类群,使用最大似然分析和贝叶斯分析构建系统发育树.由图5可知,该种与四胞极衣属关系更为密切,与已报道的绿色四胞极衣种间的进化关系明确,聚成一个单系群(BI=100,PP=1).
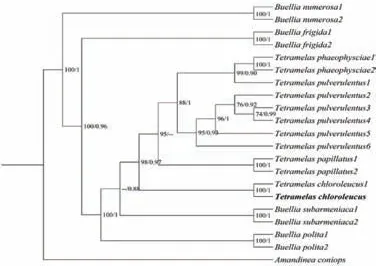
图5 绿色四胞极衣ITS序列系统发育树
2.2 分种描述
2.2.1 多变橙衣Caloplaca variabilis (Pers.) M¨ull.Arg.M´em.Soc.Phys.Hist.nat.Gen`eve 16(2): 387(1862)(图6)
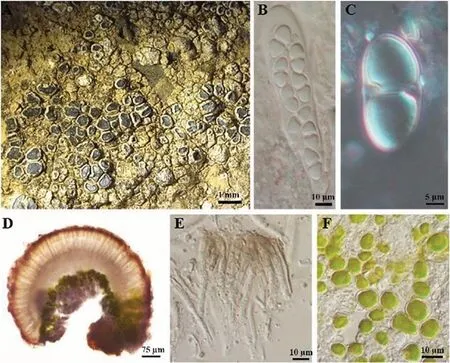
图6 多变橙衣地衣体形态和子囊盘解剖结构图
GenBank登录号:ON159743.
地衣体壳状,石内生,灰色至深灰色;子囊盘无柄,圆形或略带棱角状,直径常为1~2 mm,子囊盘盘面深灰色至黑色,有时覆粉霜,盘缘明显,与地衣体同色或白色.子囊盘茶渍型,子实上层浅褐色至深褐色,12~18.05 μm,未见晶体;子实层和子实下层无色透明;子囊细长棍棒状,8孢,55~63 μm;子囊孢子无色,对极型双胞,15~20 μm×7.5~10 μm,中间缩狭部2~5 μm;侧丝具隔,不分支或部分少见二叉分支,顶端略膨大.
化学反应:地衣体K-,C-,KC-,I-,P-;子实上层K+淡紫色.
化学成分:TLC未测出次级代谢产物.
基物:岩石生.
引证标本:巩乃斯林场,2003年8月16日,2 200 m,N 43°32′10′′、E 84°22′25′′,热衣木·马木提,GNS 020.
分布:欧洲[17]、北美洲[18]分布;国内分布于陕西[19-20].
讨论:该种与双生橙衣(Caloplaca diphyodes)形态相似,但后者地衣体具柄而多变橙衣地衣体石内生;多变橙衣子囊盘盘面附着粉霜,而双生橙衣子囊盘不具粉霜.
2.2.2 旱生野粮衣Circinaria arida Owe-Larss.A.Nordin & Tibell (2011)(图7)
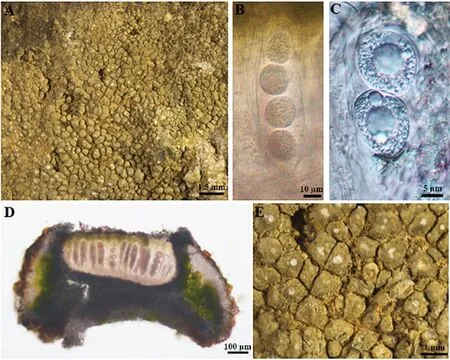
图7 旱生野粮衣地衣体形态和子囊盘解剖结构图
GenBank登录号:ON159595.
地衣体壳状,棕色或深,有时可见较浅裂缝,龟裂片不规则形状;每裂片生1~3子囊盘,常为圆形或略微棱角状,0.25~0.5 mm;盘面黑色,覆粉霜;盘缘明显,白色,部分与地衣体同色或浅.子囊盘茶渍型,子实上层橄榄绿色至棕色,15.6~33.4 μm;子实层无色透明,125~195 μm;子囊棒状,平茶渍型顶器,常2~4孢;子囊孢子无色,单细胞,亚球形至球形,17.5~35 μm×15~27.5 μm.
化学反应:地衣体K-,C-,KC-,I-,P-.
化学成分:TLC检测含Aspicilin.
基物:岩石生.
引证标本:巩乃斯林场,2018年8月16日,2 150 m,N 43°14′47′′、E 84°40′55′′,热衣木·马木提,201815 81-B.
分布:北半球分布[21];国内分布于甘肃、青海、内蒙古[22-23].
讨论:该种主要特征为地衣体棕色,子囊盘盘面具大量粉霜,子囊2~4孢,含有Aspicilin.该种外部形态与平茶渍属(Aspicilia)相近,但其子囊中所含子囊孢子数目以及子囊孢子大小与平茶渍属不相符.该种与内卷野粮衣(Circinaria contorta)近似,但内卷野粮衣地衣体较薄,白色至灰绿色.
2.2.3 祁连鸡皮衣Pertusaria qilianensis Ren Q&Zhao ZT.in Ren Q et al.Mycotaxon 108: 441-442(2008)(图8)
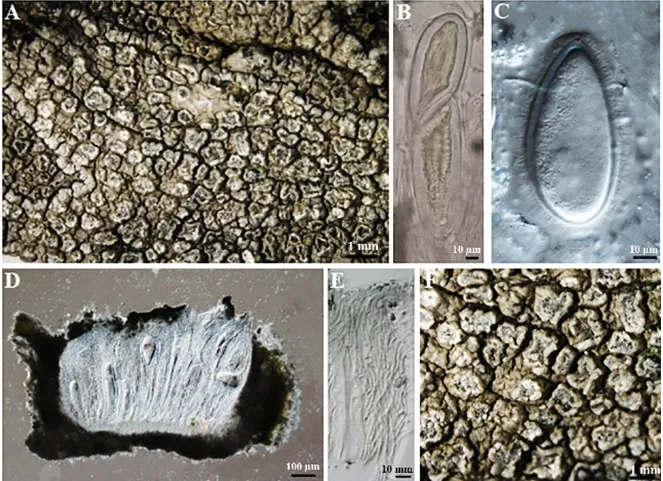
图8 祁连鸡皮衣地衣体形态和子囊盘解剖结构图
GenBank登录号:ON175982,ON175984.
地衣体壳状,上表面粗糙,浅灰色至灰色;具盘状果疣;子囊盘埋生于果疣,每果疣内生1子囊盘,盘面黑色,不规则状,覆白色粉霜,粉霜薄厚与子囊盘发育程度有关.子囊盘茶渍型,子实上层无色,22~38 μm,子实层无色,231~309 μm;子囊2孢,78~84.5 μm×297~305 μm;子囊孢子浅黄褐色至无色,单胞,115~124 μm×206~219 μm.
化学反应:地衣体K-,C-,KC-,I-,P-.
化学成分:TLC检测含Planaic酸.
基物:树生和朽木生.
引证标本:巩乃斯林场,2013年7月30日,2 660 m,N 43°11′78′′、E 84°20′114′′,热衣木·马木提,GNS002-B,GNS014;巩乃斯林场,2018年8月18日,2 440 m,N 43°36′36′′、E 84°19′37′′,热衣木·马木提,20181531,201815 25-C;巩乃斯林场,2018年8月19日,2 460 m,N 43°10′55′′、E 84°19′57′′,热衣木·马木提,20181661-B,20181673-A,20181673-B.
分布:仅发现于中国青海、宁夏两地[24].
讨论:该种为中国特有种,此报道前仅发现于我国青海和宁夏两地.该种的主要特征为子囊盘粉芽化,子囊2孢,含Planaic酸.在鸡皮衣属(Pertusaria)中含Planaic酸的地衣极为少见,含有Planaic酸的鸡皮衣属地衣又可以根据子囊所含孢子数量进行鉴定分辨.
2.2.4 肉饼干衣Rinodina sarcogynoides (H.Magn.) Q.Y.Zhong & Li S.Wang(图9)
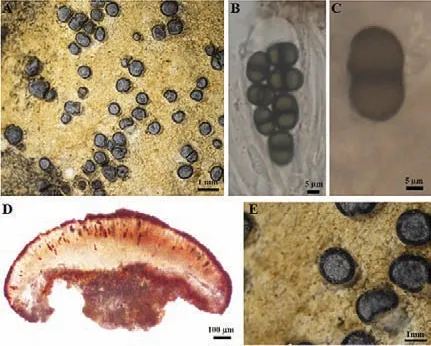
图9 肉饼干衣地衣体形态和子囊盘解剖结构图
≡Buellia sarcogynoides H.Magn., Lich.Centr.Asia 1: 145 (1940)
GenBank登录号:ON226972.
地衣体小颗粒状,浅黄色至土黄色,石内生;子囊盘圆形,0.3~1.0 mm,贴生于岩石表面,盘面黑色,可见微白色粉霜;盘缘黑色,较厚.子囊盘网衣型,子实上层棕色,19.3~30 μm;子实层无色,囊层基无色透明至浅棕色;子囊棍棒状,茶渍型顶器,8孢,42.5~48.8 μm×16.8~18.4 μm;子囊孢子棕色,双胞,15~23 μm×8~12 μm;侧丝分枝或不分枝,顶端略膨大.
化学反应:地衣体K-,C-,KC-,I-,P-.
化学成分:TLC未测出次级代谢产物.
基物:岩石生.
引证标本:巩乃斯林场,2013年7月30日,2 660 m,N 43°11′78′′、E 84°20′114′′,热衣木·马木提,GNS035.
分布:亚洲分布[25];国内分布于内蒙古、甘肃、青海和西藏[26].
讨论:该种外部形态与黑瘤衣属(Buellia)相似,但观察子囊盘解剖结构发现与饼干衣属(Rinodina)特征相吻合,因此判定为饼干衣属物种.该种的内部解剖特征与古氏饼干衣(Rinodina guzzinii)和类栗饼干衣(Rinodina castanomelodes)一致,但三物种之间外部形态差异明显.
2.2.5 绿色四胞极衣Tetramelas chloroleucus (Korb.) A.Nordin.Parerg.lich.: 191 (1860)(图10)

图10 绿色四胞极衣地衣体形态和子囊盘解剖结构图
GenBank登录号:ON159761.
地衣体壳状,黄绿色至绿色,紧贴于基物生长;子囊盘壳状或圆盘状,盘面平至凸出,黑色,直径为0.2~0.5 mm,罕更大.网衣型子囊盘,子实层101~153 μm;子囊棍棒状,8孢,51~54 μm×16~19 μm;孢子双细胞,黑褐色,11.4~13.8 μm×3.4~7.8 μm;侧丝具或不具横隔,顶端膨胀明显.
化学反应:地衣体KC+橙黄色,UV+浅橘黄色.
化学成分:TLC未测出次级代谢产物.
基物:朽木生.
引证标本:巩乃斯林场,2018年8月19日,2 460 m,N 43°10′55′′、E 84°19′57′′,热衣木·马木提,20181554-B,20181679-A.
分布:世界广泛分布;国内分布于四川、云南[27-28].
讨论:该种被发现于海拔2 400 m的岩石上.野外鉴定时,因子囊盘特征被误认为小网衣属(Lecidella)或黑瘤衣属(Buellia),解剖后发现其子囊孢子为黑色双胞,结合外部形态特征发现该种类似玫瑰黑瘤衣(Buellia erubescens),但后者具有Buellia-Type子囊孢子,而绿色四胞极衣具有Callispora-Type子囊孢子.该种还有一个明显的特征是含有6-O甲基arthothelin,地衣体呈黄绿色,UV+淡橙色至浅橘黄色反应,因此作为该种主要判定依据.
