粗糙脉孢菌7种子囊型归类教学探究
2019-11-28王春明孙英莉林昌俊王铭裕牛月李晓峰冯虎元
王春明,孙英莉,林昌俊,王铭裕,牛月,李晓峰,冯虎元
粗糙脉孢菌7种子囊型归类教学探究
王春明,孙英莉,林昌俊,王铭裕,牛月,李晓峰,冯虎元
兰州大学生命科学学院, 兰州 730000
遗传分析是本科遗传学教学的重要内容,四分子分析是真菌类遗传分析的独特方式,其中的粗糙脉孢菌顺序四分子分析不仅可以研究基因与着丝粒间的重组、基因间的重组,而且还可以精细研究同源染色体非姊妹染色单体之间的交换形式,是深入理解交换重组分子机制的重要遗传学研究材料。对此,教材中已有详尽的介绍。美中不足的是,中外教科书和相关专业期刊中缺乏对顺序四分子两基因连锁分析中7种基本子囊型归纳方法的具体介绍,令相关内容的学习留下疑惑。本文从顺序四分子两基因连锁遗传分析过程中涉及的3种四分子类型(PD、NPD和T)与两基因4种分离时期组合(Ⅰ Ⅰ, Ⅱ Ⅱ, Ⅰ Ⅱ, Ⅱ Ⅰ)的相互关系入手,设计表格,直观展现了二者的12种可能组合结果(3×4=12),并应用直观的对错符号(√、×),同时结合表后的3项分析,排除了其中的5种组合形式,最终归纳得到7种基本子囊型。本文提供了7种基本子囊型具体的归纳分析方法,解决了相关教学内容的论述不足问题,以期为真菌类顺序四分子遗传分析的教学提供帮助。
真菌;顺序四分子;基本子囊型;四分子分析;遗传分析
经典遗传学研究的一个主要任务是确定基因间的图距,这在当前综合性高校本科遗传学教学中仍然是教学的重点。由于动物、植物、细菌和病毒等不同生物的生活史千差万别,遗传学家针对不同生物的特点研究了独特的遗传分析方法。其中,四分子分析(tetrad analysis)是专门针对真菌生活史设计的遗传分析方法。四分子(tetrad)是指性母细胞减数分裂产生的4个子细胞。真菌有性生殖过程中形成的合子随即进行减数分裂,也会形成4个子细胞,这4个子细胞共同存在于子囊里,紧接着再进行一次有丝分裂,形成了含有8颗子囊孢子的子囊。这8颗子囊孢子因两两源于有丝分裂而在遗传性状上一致(图1),子囊孢子间的差异仅仅体现在最后这次有丝分裂前的四分子的遗传背景上,因此在遗传分析时,可以忽略一半的子囊孢子(忽略最后这次有丝分裂新形成的孢子),而仅仅考察减数分裂形成的四分子的遗传差异,这种针对四分子遗传性状的分析方式就是真菌类遗传分析所特有的四分子分析。
真菌的子囊类型依据内部子囊孢子排列的特点可进一步区分为顺序四分子(ordered tetrad)和非顺序四分子(unordered tetrad)。顺序四分子因其所在子囊狭小,限制了减数分裂后期两个子细胞核的自由移动,造成子细胞在子囊内的随机分布与分裂后期纺锤体的方向直接相关,如果某基因与着丝点之间发生交换,则可以通过子囊孢子在子囊中的排列情况加以研究,进而计算出该基因与着丝点之间的图距,这就是顺序四分子的着丝粒作图(centromere mapping)。顺序四分子分析通常以粗糙脉孢菌()为代表。正如戴灼华和王亚馥主编的教材中总结的那样,顺序四分子在遗传分析中具有独特的优越性,可以揭示其他遗传分析方法所不能发现的遗传本质[1]。教材具体总结了顺序四分子分析的4个优点:(1)将着丝粒视作一个基因座,计算某一基因与着丝粒的重组率;(2)子囊孢子的对称性,证明了减数分裂是一个交互的过程;(3)可检验染色单体的交换是否存在干涉现象,并且可以利用它来研究基因转变现象;(4)发现双交换不仅可以包括4线中的2线,而且还可以包括3线或4线。这些都深入揭示了交换重组的精细结构信息[1]。

图1 着丝粒作图所涉6种子囊型的形成过程
A:第一次分裂分离型(MI)子囊中的子囊孢子A/a基因与着丝粒之间没有发生交换重组,可获得2类子囊型,这2类子囊型出现的几率相等; B:第二次分裂分离型(MII)子囊中的子囊孢子A/a基因与着丝粒之间发生了1次交换重组,可以得到4类子囊型,这4类子囊出现的几率相等。
顺序四分子的某一基因与着丝粒之间发生交换可获得6种子囊型(图1),于是在分析某二基因分别与着丝粒之间的交换时,因互为独立事件的属性,理论上可得到6×6=36种子囊型。对36种子囊型进行统计分析无疑是个艰巨的任务,遗传学家通过分析归纳,将这36种子囊型归纳为7种基本子囊型,大大简化了顺序四分子的遗传分析。遗憾的是,查阅几种中英文遗传学教材,并没有发现对如何归纳的具体过程进行介绍[1-3],检索与四分子分析相关的期刊论文也未发现专门针对这7个基本子囊型归纳方法的论文[4~7]。例如,我校选用的遗传学教材对此的介绍是“一对基因杂交,有6种不同的子囊型,两对基因杂交必有6×6=36种不同的子囊类型;但是因为半个子囊内的基因型次序可以忽视,不论是n+孢子对在“上面”,+a孢子对在“下面”,还是+a在“上面”,n+孢子对在“下面”,都不过是反映着丝粒在减数分裂过程中的随机趋向而已,所以可以将36种不同的子囊型归纳为7种基本子囊型”[1]。师生普遍反映这样的描述并不能让他们明白这7种基本子囊型具体是如何归纳的。这种具体归纳方法和过程的缺失,令在教学中透彻理解7种基本子囊型的来源一直存在困难。因此,本文针对这7种基本子囊型具体的归纳过程,力求给出清晰可控的归纳方法,以期为真菌类顺序四分子遗传分析的教学提供帮助。
1 着丝粒作图涉及到6种子囊型
顺序四分子在子囊中的排列直接反映减数分裂时同源染色体以及姊妹染色单体的分离和分布情况。图1A示基因A/a与着丝粒间无交换情况下产生2类子囊型,亦称MI型,代表A和a这对等位基因在减数分裂的第一次分裂时即已分离,亦即得到的二分子是A/A和a/a,四分子是AAaa和aaAA。图1B示A/a基因与着丝粒之间发生一次交换的情况下,产生4类子囊型,亦称MII型,表示A和a基因在减数分裂的第二次分裂后的四分子时期才分离,而在第一次分裂形成的二分子中仍然以A/a的杂合状态存在。因此,在考察基因A/a与着丝粒间的图距时,一共可获得6种子囊型,其中2种MI型是非交换型,4种MII型是交换型,亦即可将这6种子囊型归纳为MI和MII两类。中文教材对着丝粒作图已有详细介绍,并给出了利用这6种子囊型计算基因与着丝粒之间图距的计算公式[1],在此不再详述。
2 两个连锁基因作图所涉7类基本子囊型的归纳方法
上述着丝粒作图研究的是一个基因与着丝粒之间的交换,包含MI和MII两类、共6种子囊型。如果分别考察两个基因与着丝粒间的交换重组,进而考察两基因间的重组,则可预期有6×6=36种不同的子囊类型。例如,考察A基因与着丝粒间的交换,可得6种子囊型;考察B基因与着丝粒间的交换,同样可得B基因的6种子囊型。那么,同时考虑A和B两个基因间的关系,就可能得到6×6=36种子囊型。
以基因型AB × ab个体杂交为例,如果仅仅考察A基因与B基因间是否发生了交换重组,即不考虑孢子排列,只考虑A和B两个基因的组合,可以依据后代子囊所含子囊孢子的基因型将后代子囊归纳为3类(表1):(1)后代子囊中的子囊孢子基因型如果只有AB和ab两种,与两个亲本相同,这样的子囊称为亲二型(parental ditype, PD);(2)后代子囊中的子囊孢子如果只有Ab和aB两种基因型,即二者基因型与双亲(AB, ab)均不同,则将此类子囊称为非亲二型(nonparental ditype, NPD);(3)还有一种情况是后代子囊中的子囊孢子基因型包括AB、ab、Ab和aB 4种组合,既有双亲的基因型(AB, ab),又有重组型(Ab, aB),这样的子囊含有4种基因型的孢子,故称为四型(tetratype, T)。由于交换重组的交互性,一般不会出现这3类以外的子囊类型。可见,PD子囊中的孢子在A和B基因间没有发生重组,保持亲本的基因型(AB, ab);NPD子囊中的孢子在A和B基因间全部发生了重组,都是重组型(Ab, aB);而T子囊中的孢子有一半是亲本型(AB, ab)一半是重组型(Ab, aB)。因此,考察A和B基因是否重组,可将后代子囊分为PD、NPD和T 3类。中文教材中详细介绍了这3类子囊的形成机制,以及如何利用它们计算二基因间的图距[1],此处不再赘述。本文要解决的问题是表2中列出的7种基本子囊型是如何从36种可能子囊型中归纳出来的。
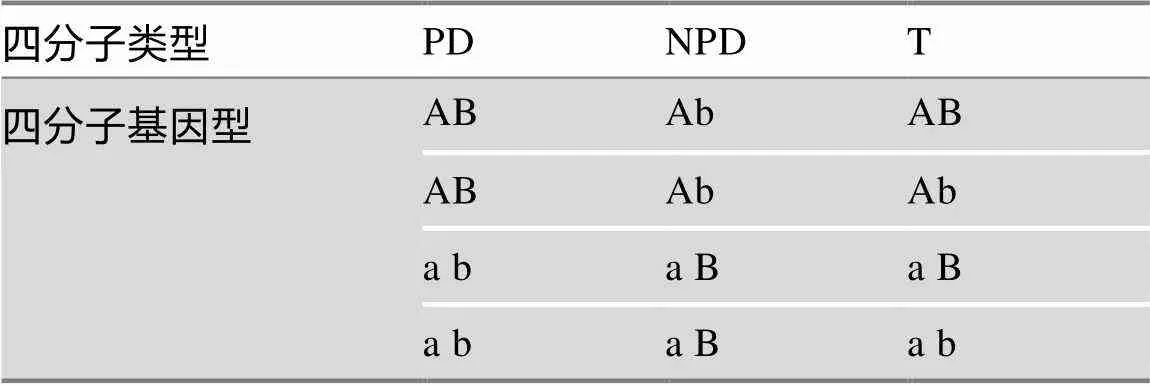
表1 AB×ab杂交子代可归纳为3种四分子类型
如前述,如果分别考察A/a基因及B/b基因与着丝粒间的交换重组,可分别将子囊类型区分为MI和MII型(图1)。综合考虑这些特点,即可将36种子囊型归纳为7类基本子囊型(表2)。
由表2可见,依据A基因和B基因在子囊形成过程中的分离时期,可将子囊分为4类:即Ⅰ Ⅰ、Ⅱ Ⅱ、Ⅰ Ⅱ、Ⅱ Ⅰ分别代表A/a基因和B/b基因所属MI或MII分离型的4种组合方式。即这两对等位基因同时属于MI分离(Ⅰ Ⅰ);同时属于MII分离(Ⅱ Ⅱ);A/a属于MI分离,B/b属于MII分离(Ⅰ Ⅱ);A/a属于MII分离,B/b属于MI分离(Ⅱ Ⅰ)。此外,依据子囊孢子的基因型,可将子囊分为PD、NPD、T 3类。应用这两个特征(4种分离型组合、3类子囊型)即可将36种子囊型归纳为7种基本子囊型。也就是说,表2所示7类基本子囊型可以看作是4种分离型组合与3类四分子类型共同决定的(表3)。
由表3可见,A/a和B/b基因的4类分离型与3类四分子类型两个条件理论上可以归纳得到3×4= 12种基本子囊型。其中有5类子囊型是不存在的,可作如下推理分析:

表2 AB × ab杂交子代归纳出7类基本子囊型
Ⅰ:第一次分裂分离型MI;Ⅱ:第二次分裂分离型MII。
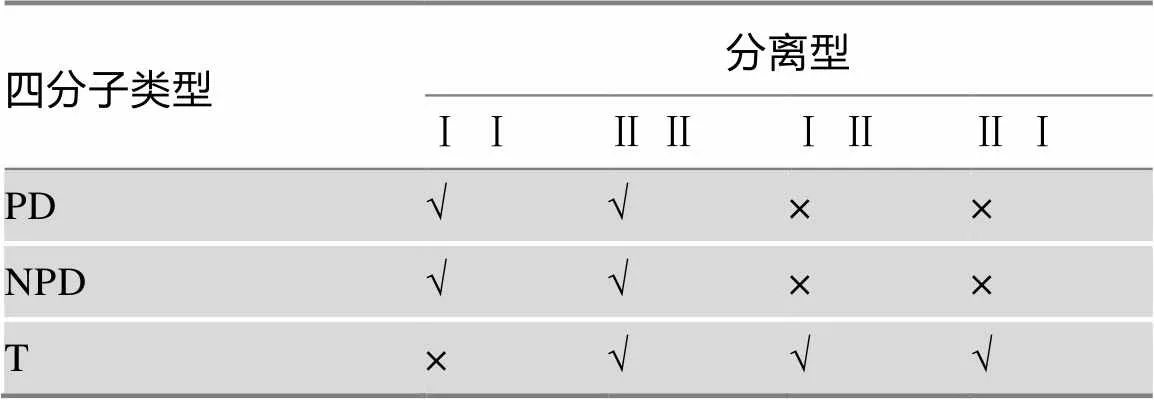
表3 7类基本子囊型的归纳推导方法
Ⅰ:第一次分裂分离型MI;Ⅱ:第二次分裂分离型MII。√:有此类子囊型;×:无此类子囊型。
(1) PD类型的子囊中,A/a基因和B/b基因的分离时期要么同为MI,要么同为MII,不可能一个是MI而另一个是MII,反之亦然。例如:A/a基因为MI,AA/aa,对应的B/b基因如果是MII,那么四分子的基因型将是AB Ab/aB ab,此类子囊属于T型而非PD。因此表2中的PD与Ⅰ Ⅱ和Ⅱ Ⅰ两类分离型组合的子囊是不存在的。故排除2类子囊型。
(2) NPD类型的子囊中,A/a基因和B/b基因的分离型组合与PD类型同理。因为二基因不同分离型的组合,得到的将是T型,亦非NPD。故排除2类子囊型。
(3) T类子囊中,A/a基因和B/b基因的分离时期同为MI是不可能的。举例如下,A/a基因的MI分离(AA/aa),如果B/b基因也是MI分离,则其可能的子囊型为AB AB/ab ab(PD)或Ab Ab/aB aB (NPD),而非T型。故排除1类子囊型。
由此可见,表3理论上的12类子囊型中有5类是不成立的,12−5=7,于是就可以根据4类分离型和3类四分子类型通过分析归纳得到7类基本子囊型。表3内容结合其后的3点分析很好地给出了7类基本子囊型的归纳推导方法。
3 讨论
近期的遗传学教学论文逐渐回归和重视教学内容的研究[8~10],使相关论文的含金量得到较大的提升。这其中,既有关于新的教学内容的建议[8,10],也有对具体实验设计的探讨[9],这无疑会对遗传学教学起到重要的提升和促进作用。本文作者在长期的遗传学教学实践过程中,也比较注重教学内容的研究,曾就遗传漂变定义中“抽样”概念的使用专门撰文提出建议[11],理清了“抽样”概念的具体涵义,助力遗传漂变概念的讲解。本文则针对教材对丝状真菌粗糙脉胞菌顺序四分子两个连锁基因遗传分析,涉及到的7种基本子囊型的归纳方法介绍得不够详尽这一不足,结合自己多年教学中对这一问题的思考,以及查阅相关文献,总结出一套较为简便易懂的归纳方法(表3及相关推理),以期提高顺序四分子分析的教学效果。
如前所述,我校选用的高教版遗传学教材对这一问题并没有给出具体的归纳方法[1],国外的英文版遗传学教材对这一问题也没有给予足够的重视。Hartwell等主编的2011版教材提及并列表介绍了这7类子囊型(These tetrads were classified in seven different groups—A, B, C, D, E, F, and G—as shown in Fig. 5.22a.),但并未介绍具体的归纳方法[3]。而另外两部英文教材则根本没有提及这7类基本子囊型的情况[2,12]。其中Anthony等主编的2015版教材虽然没有在正文中涉及,但是在课后习题中提出了为什么只有7种子囊型这一问题(Why are there only seven classes?)[2],但遗憾的是,作者给出的习题答案也没有介绍具体的归纳方法。这些教材存在的不足,促使本文作者继续查阅相关文献,以期寻找一些帮助。
外文数据库中检索到相关度较高的英文文献很少,比较有参考价值的是一篇题为“顺序四分子统计分析”的文献[7],作者竟然写到要“检查全部36种子囊……”(After examining all 36 distinguishable patterns, the number of distinct probabilities is seven……),这显然难以在课堂上推广。从CNKI数据库检索到涉及四分子分析的3篇中文文献,有一篇没有提及7类基本子囊型的问题[6],其余两篇则不同程度地涉及到了这一问题[4,5]。2008年杨先泉等[5]发文详细列出了36种子囊型,并给出了对应的7类基本子囊型,但并未从理论上给出归纳总结的依据和推导方法,没有重点介绍7类基本子囊型的分类问题。2009年刘卫东[4]对此问题给出了较好的介绍,“根据上述分类原则,按直线排列孢子对的组合方式,M1M1型子囊只有PD和NPD,M1M2和M2M1只有T,M2M2可有PD、NPD和T型,这样可把所有孢子囊类型简化为7种类型”。这一描述与我们在本文中运用的是同一逻辑,只是在归纳方法上还稍欠直观。
总之,从方便教学和易于学生掌握出发,本文结合二基因“分离型组合”以及“四分子类型”两个条件,通过表3直观展现了二者的12类可能组合(3×4=12),并引入对错符号(√、×),附以表后的3项分析,排除了其中的5类,最终得到7类基本子囊型。本文明确提出了7类基本子囊型具体的归纳分析方法,解决了相关教学内容的遗留问题,为教学实践提供了具体的素材。希望表3的分析方法能够对真菌类顺序四分子遗传分析的教学提供帮助。
[1] Dai ZH, Wang YF. Genetics. Higher Education Press, 2016.戴灼华, 王亚馥. 遗传学. 高等教育出版社, 2016.
[2] Griffiths AJF, Wessler SR, Carroll SB, Doebley J. Introduction to Genetic Analysis. W. H. Freeman and Company, 2015.
[3] Hartwell L, Hood L, Goldberg ML, Reynolds A, Silver LM. Genetics: from genes to genomes. McGraw-Hill Companies, Inc. 2011.
[4] Liu WD. New method of Fungi ordered tetrad analysis., 2009(06): 15, 34.刘卫东. 真菌顺序四分子分析方法新解. 咸宁学院学报, 2009, 29(06): 15, 34.
[5] Yang XQ, Zhao Q, Fu TH. Ordered tetrad analysis of two genes in., 2008, 30(6): 801–806.杨先泉, 赵勤, 傅体华. 脉孢霉两对基因顺序四分子分析. 遗传, 2008, 30(06): 801–806.
[6] Hou WR, He YK. The meaning of gene recombination studied by tettada analysis of fung., 1991(01): 1–10.候万儒, 何奕騉. 试论真菌四分子分析在基因重组研究上的意义. 成都大学学报(自然科学版), 1991(01): 1–10.
[7] Zhao H, Speed TP. Statistical analysis of ordered tetrads., 1998, 150(1): 459–472.
[8] Ma L, Zhang TT. A teaching design to introduce chromosomal aberration in genetics using case studies of chimeric genes., 2018, 40(12): 1129–1135.马磊, 张婷婷. 应用嵌合基因实例拓展遗传学染色体畸变的教学. 遗传, 2018, 40(12): 1129–1135.
[9] Liu ZQ, Zhao YX, Fu XL, Li N. Design and exploration of genetic experiments for non-Mendelian segregation., 2019, 41(3): 262–270.刘自强, 赵苑秀, 傅雪琳, 李楠. 偏孟德尔分离的遗传学实验设计与探讨. 遗传, 2019, 41(3): 262–270.
[10] Wu K, Luo ZH. Application of entomological cases in genetics teaching., 2019, 41(4): 349– 358.吴凯, 罗朝晖. 昆虫学案例在遗传学教学中的应用. 遗传, 2019, 41(4): 349–358.
[11] Wang CM. Exploration of the concept of genetic drift in genetics teaching of undergraduates., 2016, 38(1): 82–89.王春明. 本科遗传学教学中的遗传漂变概念探讨. 遗传, 2016, 38(1): 82–89.
[12] Snustad DP, Simmons MJ. Principles of Genetics. John Wiley & Sons, Inc., 2012.
How to deduce the seven basic class asci of ordered tetrads in
Chunming Wang, Yingli Sun, Changjun Lin, Mingyu Wang, Yue Niu, Xiaofeng Li, Huyuan Feng
Genetic analysis is an important part of undergraduate genetics teaching and tetrad analysis is unique and integral for genetic analysis of fungi. The ordered tetrad inis an important material for genetic analysis, which can not only be used to study recombination between genes and centromeres, but also between genes themselves, as well as study the fine cross patterns between non-sister chromatids of homologous chromosomes. However, in textbooks and related professional journals, there is a lack of specific introduction to the induction methods of the seven basic class asci used in two genes analysis. In the present paper, we designed a table presenting the correlation between the three tetrad types (PD, NPD, T) and the four segregation pattern groups (Ⅰ Ⅰ, Ⅱ Ⅱ, Ⅰ Ⅱ, Ⅱ Ⅰ) to visually show the 12 possible combinations (3×4=12). Then five of them were excluded through the "×" symbol and in addition with three comments attached with the table, thus finally we obtained seven basic ascus types. We hope that this analytical method can assist the teaching of ordered tetrad analysis in.
fungus; ordered tetrad; basic classes ascus;tetrad analysis;genetic analysis
2019-06-24;
2019-09-26
国家基础学科人才培养基金项目(编号J1210033,J1210077)和兰州大学教学研究项目资助[Supported by the Undergraduate Training Project of National Science Foundation of China (Nos. J1210033, J1210077), and Teaching research project of Lanzhou University]
王春明,博士,副教授,硕士生导师;研究方向:细胞遗传学。E-mail: cmwang@lzu.edu.cn
10.16288/j.yczz.19-181
2019/9/26 17:19:06
URI: http://kns.cnki.net/kcms/detail/11.1913.r.20190925.1111.002.html
(责任编委: 谢建平)
