短期增温对若尔盖高原木里薹草沼泽生态系统呼吸的影响
2023-05-13刘宏强张元姝熊银洪董李勤
刘宏强, 张元姝, 言 迫, 熊银洪, 董李勤, 张 昆
(1.西南林业大学国家高原湿地研究中心/湿地学院,云南 昆明 650224;2.四川若尔盖湿地国家级自然保护区管理局,四川 阿坝 624500;3.西南林业大学地理与生态旅游学院,云南 昆明 650224;4.国家林业和草原局西南生态文明研究中心,云南 昆明 650224)
自工业革命以来,全球平均温度不断上升,IPCC第六次评估报告预测,未来20 a全球平均温度的上升幅度将会达到或超过1.5 ℃[1].全球变暖是环境变化的主要驱动因素之一,影响着生物多样性、生态系统结构与功能,最终威胁到生态系统服务[2].近年来,温度升高对陆地生态系统碳循环关键过程的影响已成为全球变化生态学研究的热点[3-5].
生态系统呼吸是生态系统以CO2的形式向大气释放碳的途径,是植物叶片呼吸、根系呼吸、微生物呼吸、土壤有机质分解和动物呼吸的总和[3].作为陆地最大的碳通量之一,生态系统呼吸的微小波动都可能导致大气CO2浓度发生显著变化[6],对气候变暖产生正反馈或负反馈.大量研究从温度[4,7]、养分[5,8]、水分[9-10]、放牧[6,11]等不同环境因子探讨了生态系统呼吸的响应模式,研究发现生态系统呼吸对温度升高的响应尤为敏感[6].青藏高原植物生长最适温度全球最低,当前青藏高原生长季温度仍低于其植物生长最适温度[12],加之作为气候变化的敏感地区[13],其生态系统呼吸对全球变暖的响应比其他地区更为显著[14].研究发现,温度升高延长生长季[15],能满足植物对热量的需求,通常促使植物株高、叶面积、叶片数和群落密度的增长[16-17],提高植物生产力,从而提高生态系统呼吸速率[5,7];也有研究发现,受植被物候加速[4]和年际气候特征[18]的影响,增温可能不会显著改变生态系统呼吸速率,甚至出现降低生态系统呼吸速率的情况[19].普遍认为温度升高对土壤呼吸的影响主要表现在土壤温度[20]和土壤水分[9,19].在较为湿润地区,通过提高土壤温度、改善植物根系和提高土壤微生物活性,促进土壤呼吸对全球变暖的积极响应[14];在较为干旱地区土壤温度升高造成土壤含水率降低,对土壤微生物和根系产生干旱胁迫[20-21],可能不会对土壤呼吸产生积极影响[9],甚至抑制土壤呼吸[19].土壤温度、水分、养分条件,以及植物和微生物群落特征等因素的差异,导致生态系统呼吸对增温的不同响应[11,22-23].
高寒草本沼泽因其较高的净初级生产力而对维持青藏高原陆地生态系统碳平衡发挥着重要作用[24],目前在青藏高原开展的增温试验多集中在高寒草地和沼泽化草甸生态系统,而对常年积水高寒草本沼泽的研究相对较少.鉴于此,本研究在若尔盖高原纳勒乔沼泽湿地进行原位增温控制试验,选择开顶式增温箱(open-top chambers, OTC)对广泛分布的木里薹草(Carexmuliensis)沼泽生态系统进行被动增温,分析增温对木里薹草生长指标(株高、叶长、群落密度和地上生物量)、木里薹草沼泽土壤呼吸和生态系统呼吸的影响,以期为全球气候变化下高寒薹草沼泽碳循环研究提供参考.
1 材料与方法
1.1 研究区概况
若尔盖湿地总面积为1 665.7 km2,是青藏高原高寒湿地生态系统的典型代表,是我国分布最为集中的高寒沼泽湿地分布区[25].木里薹草是若尔盖高寒草本沼泽的主要建群种[25-26],在若尔盖高原分布面积最广.湿地为高原浅丘沼泽地貌,丘陵断续分布,丘顶浑圆,丘间开阔,地势平坦.大陆性气候特征明显,气候寒冷,四季区分不明显,年均温度1.4 ℃,年均降水量657 mm,年均蒸发量1 232 mm.湿地内土壤类型主要为沼泽土、草甸土壤、高原褐土及人工草地土壤.试验样地位于若尔盖湿地国家级自然保护区纳勒乔木里薹草沼泽湿地(33°30′52″N,102°39′37″E),海拔3 450 m,样地常年积水,沼泽建群种为木里薹草,伴生种为海韭菜(Triglochinmaritima)、溪木贼(Equisetumfluviatile)、杉叶藻(Hippurisvulgaris)、矮泽芹(Chamaesiumparadoxum)和睡菜(Menyanthestrifoliata)等.试验样地土壤为厚层泥炭土,厚度为5.40 m,pH值为5.80~6.37.
1.2 样地设置
依据典型性和代表性原则在若尔盖湿地国家级自然保护区纳勒乔木里薹草沼泽湿地建立增温控制试验平台(图1),样地大小为100 m×100 m,周边设置围栏围封以杜绝人为干扰.2020年6月,在样地内随机选取4 m×4 m木里薹草长势一致的小区,每个小区内设置增温(W)和对照(CK)2个配对处理,每个处理设置5个重复.采用OTC进行增温试验,OTC增温与温室效应原理相似,并能保证样地土壤物理结构的完整性[16].OTC为正六边形,由5 mm厚亚克力板制成,其下部由6块边长为120 cm的正方形亚克力板围成,上部由6块上边长86 cm、下边长120 cm、高60 cm的梯形围成,近地面5 cm处开有多个圆孔以保证水体自由流动.OTC增温装置全年放置在野外.每个处理单元内嵌入40 cm×40 cm的静态箱基座和直径20 cm、高30 cm的土壤环,分别用于生态系统呼吸和土壤呼吸的定位观测,基座和土壤环嵌入地表5 cm.

图1 增温试验控制平台
1.3 生态系统呼吸速率和土壤呼吸速率的监测
于2021年5—9月,每月选取一个晴朗无云或少云的上午,于9:00—11:00测量生态系统呼吸速率和土壤呼吸速率.生态系统呼吸速率测量采用静态箱法.静态箱为暗箱,外由隔热棉包裹,长、宽、高分别为40、40、50 cm.利用LI-6400XT便携式光合仪对CO2浓度进行测定,测量间隔为10 s,持续测量2 min.测量时,静态箱内部悬挂2个小风扇,以混合箱内气体.土壤呼吸速率测量方法与生态系统呼吸速率测量方法一致,在测量日期前一天去除每个土壤环内植物,对新生植物进行齐地面剪除,直接封闭土壤环(作为静态箱),记录土壤环顶部到水面的距离以计算气体通量.气体通量计算公式[27]为:
式中,F为CO2通量(mg·m-2·s-1),∂C′/∂t为CO2浓度斜率,M为CO2摩尔质量(g·mol-1),V0为标准摩尔体积(22.4 L·mol-1),P/P0为测量大气压与标准大气压的比值,T0为标准状态下气体绝对温度(℃),t为箱内实测空气温度的平均值(℃),H为静态箱高度(cm).
1.4 环境要素获取
利用土壤温湿度记录仪(TMS5-50)同时测定-10 cm土壤温度和50 cm大气温度,测量间隔为15 min.在测量生态系统呼吸速率时,利用温度记录仪(HOBO MX2201)同步测量静态箱内的温度,记录间隔为10 s.2021年8月在试验样地采集表层土壤样品,用于测量土壤养分状况.土壤采集深度为10 cm,每个样地采集3次,共采集30个土壤样品.将采集的土壤样品分为两部分,一部分于-20 ℃冷冻保存,用于测量土壤速效氮、有效磷等有效养分含量;另一部分自然风干、过100目筛后测定土壤总碳、总氮和总磷含量.
1.5 木里薹草测量和地上生物量估算
在静态箱基座内(40 cm×40 cm)选择长势均一的10株植物,利用卷尺测量其株高和叶长,株高为从植株出土处至主干最长叶叶尖,叶长为从木里薹草主干最长叶基部至叶尖[28].为保证基座内植物的完整性,采用生物量模拟的方法估算地上生物量[29].在样地附近随机剪取30株健康木里薹草(共150株),记录其株高、叶长,并带回实验室于65 ℃烘干24 h后,称取其生物量,通过逐步线性回归分析得出健康木里薹草单株生物量与株高、叶长的关系,地上生物量=(0.014×株高+0.005×叶长-0.238)×群落密度(P<0.001,R2=0.851).
1.6 数据分析
利用SPSS 22.0对试验数据进行整理和分析,增温处理和对照间差异显著性测验采用单因素方差分析(显著性均设定在0.05水平),图表中数据为平均值±标准差.
2 结果与分析
2.1 增温箱内外环境因子的变化
由图2可知,在2021年5—9月,增温样地平均温度为12.0 ℃,变化范围为3.4~18.1 ℃,对照样地平均温度为10.8 ℃,变化范围为2.7~16.6 ℃,OTC增温效果为1.2 ℃.OTC对土壤温度和水位深度没有显著影响,样地土壤温度变化范围为3.1~15.0 ℃,水位深度变化范围为7.2~16.4 cm.
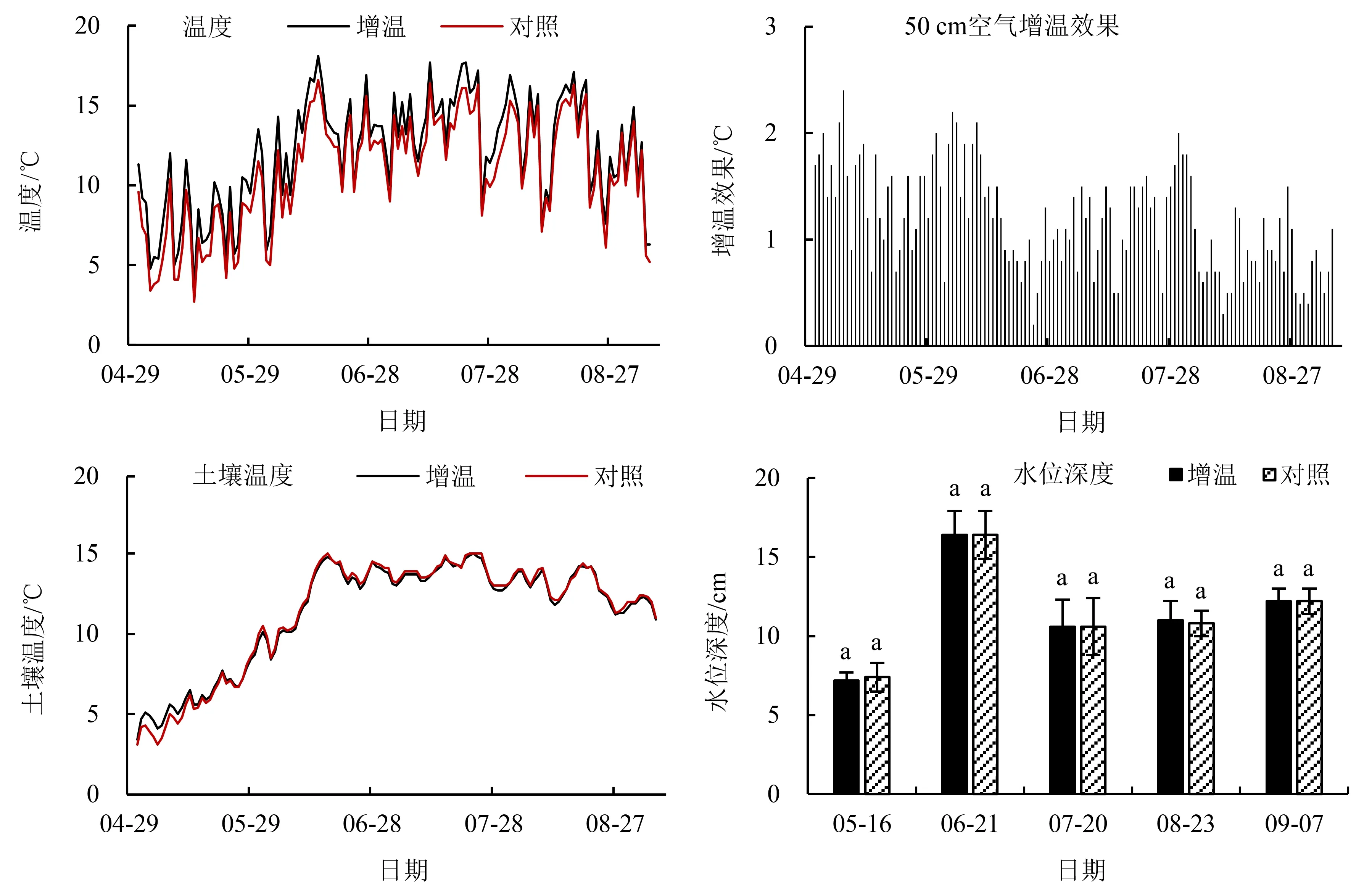
不同小写字母代表不同处理间的差异显著(P<0.05).
2.2 增温对土壤养分含量的影响
由表1可知,短期增温显著提高若尔盖高寒木里薹草沼泽土壤的速效磷含量(P<0.05),但土壤总碳、总氮、总磷和速效氮含量没有出现显著差异(P>0.05).

表1 不同处理下土壤养分含量的变化1)
2.3 增温对木里薹草生长的影响
由表2可知,温度升高显著增加木里薹草株高(P<0.05),平均增加7.09%;显著降低群落密度(P<0.05),平均降幅为12.29%.木里薹草群落生物量平均降低7.99%(P>0.05),其中,生长季前期(5—6月)无显著差异(P>0.05),生长季中后期(7—9月)增温处理降低趋势显著(P<0.05).增温对木里薹草叶片长度无显著影响(P>0.05).
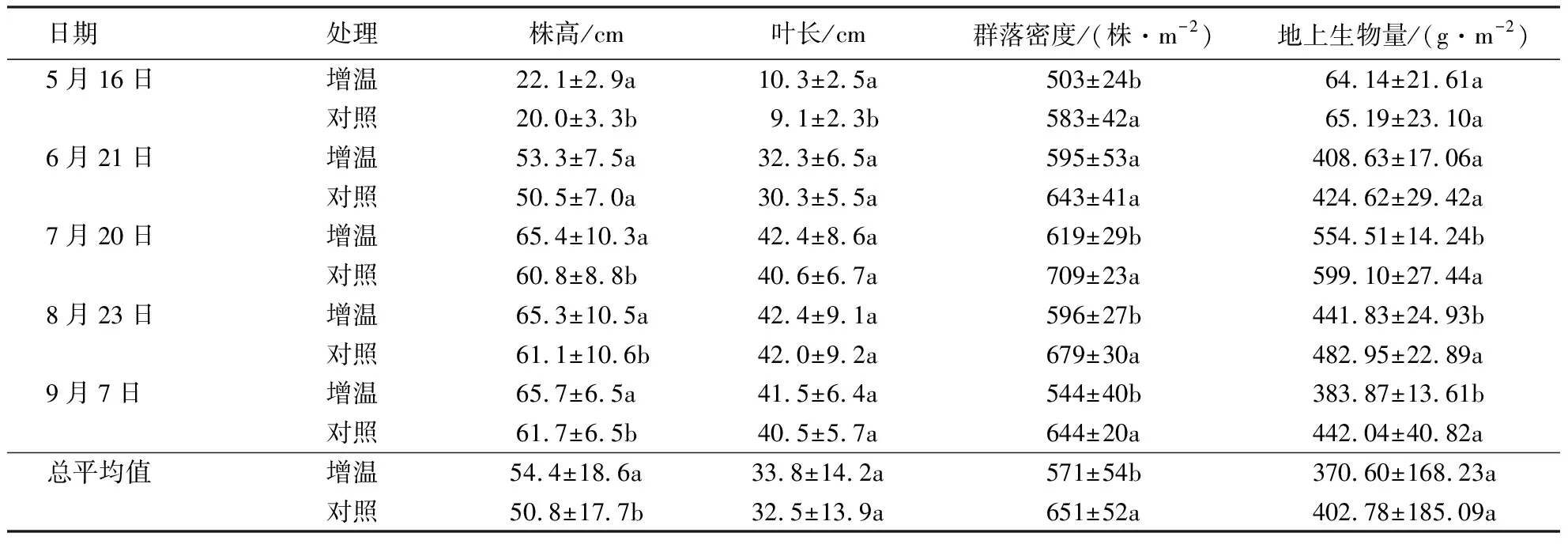
表2 不同处理下木里薹草株高、叶长、群落密度和地上生物量动态变化特征1)
2.4 增温对生态系统呼吸速率的影响
由图3可知,木里薹草沼泽生态系统呼吸速率和土壤呼吸速率在整个观测期间呈现明显季节性变化,生态系统呼吸速率和土壤呼吸速率最大值均出现在7月20日,分别为(6.95±0.69)和(1.84±0.31)μmol·m-2·s-1.增温处理下生态系统呼吸速率显著降低(P<0.05),平均降低26.33%,其中,生长季前期(5—6月)不同处理间生态系统呼吸速率无显著差异(P>0.05),生长季中后期(7—9月)增温显著降低生态系统呼吸速率(P<0.05).土壤呼吸速率略有增加(P>0.05),平均增加13.60%.
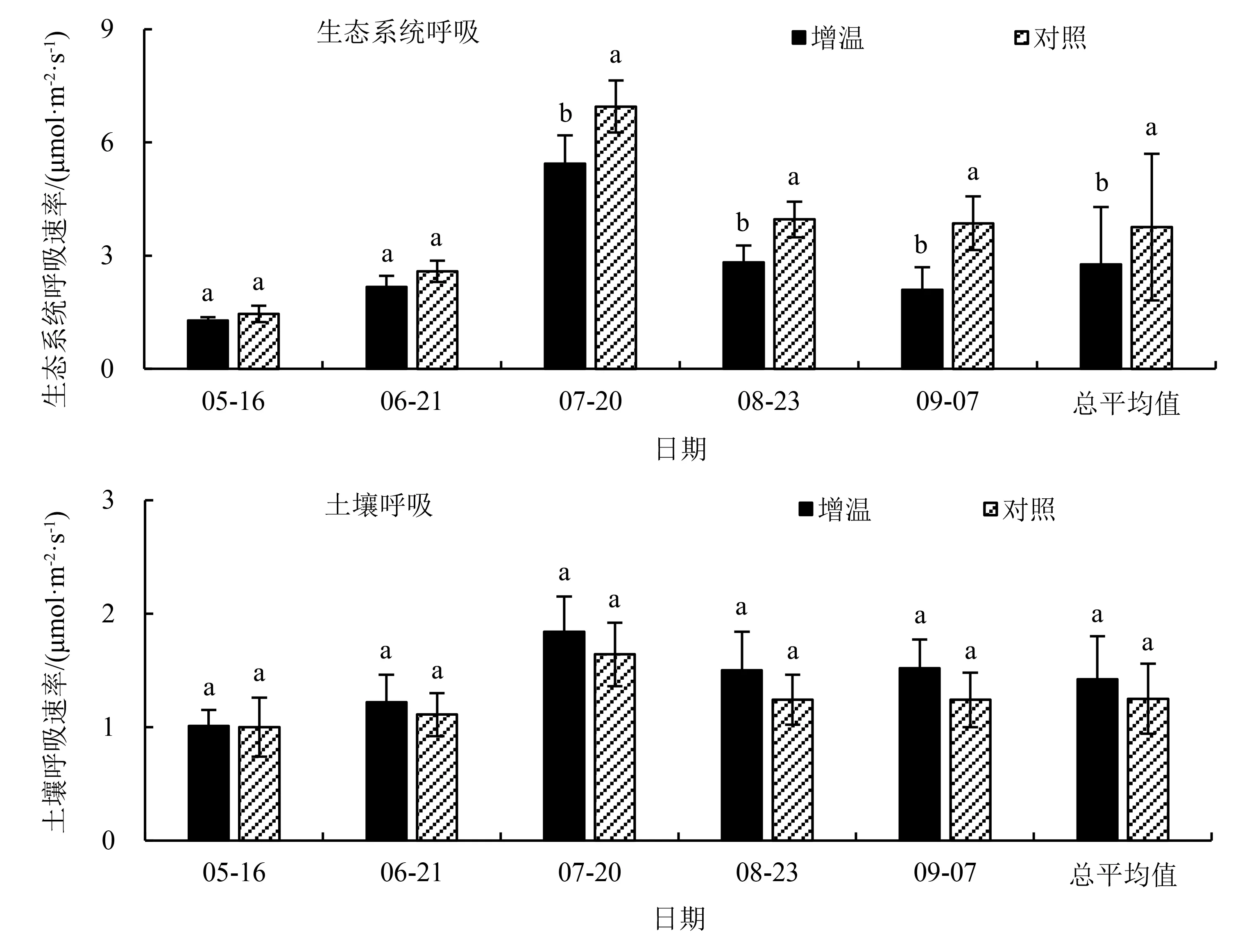
不同小写字母代表不同处理间的差异显著(P<0.05).
由图4可知,指数方程能较好地拟合生态系统呼吸速率与环境温度的关系,增温处理和对照的生态系统呼吸速率均随温度的升高而增加.增温处理和对照生态系统呼吸的温度敏感性(Q10)分别为2.4和2.6.
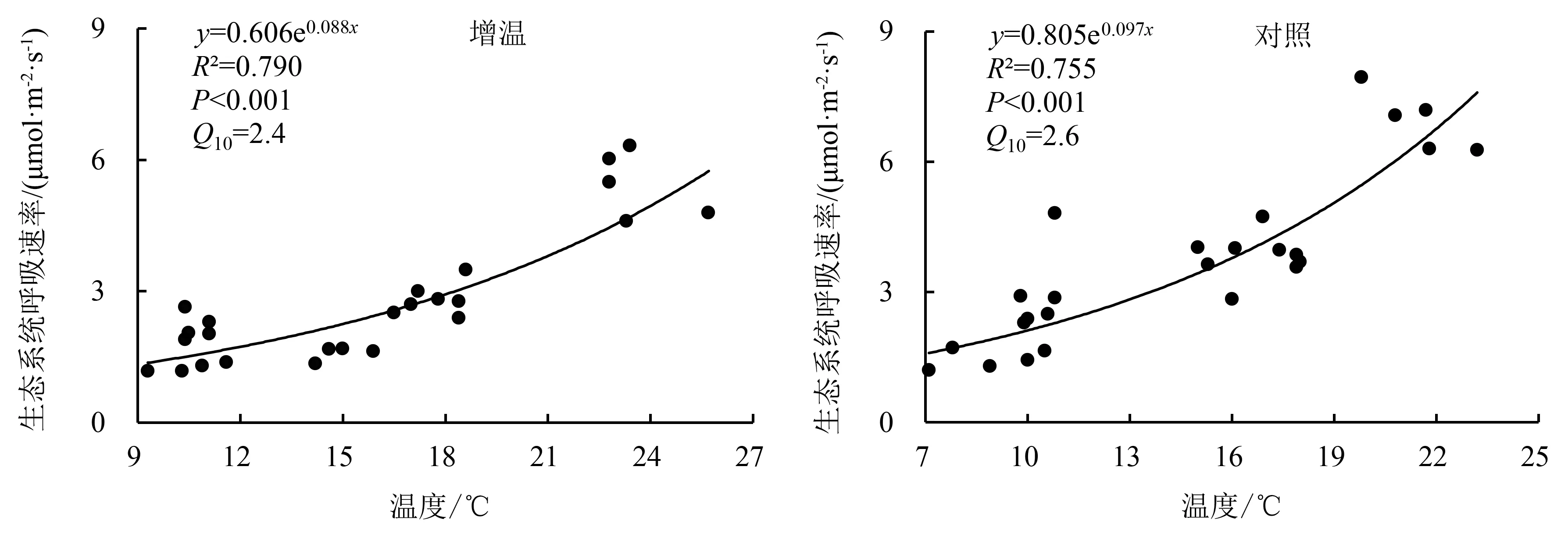
图4 生态系统呼吸对温度的敏感性
由图5可知,增温处理和对照的土壤呼吸速率均随温度升高而增加.增温处理和对照土壤呼吸的温度敏感性分别为1.5和1.3,但拟合度较低,分别为0.257和0.167.
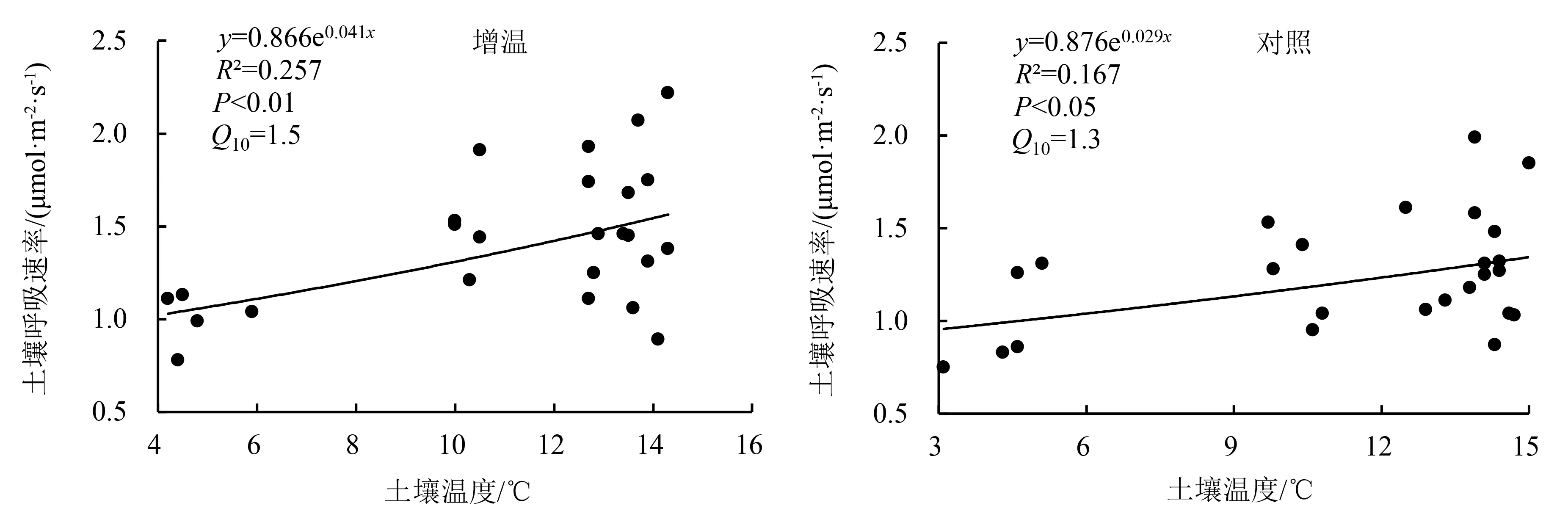
图5 土壤呼吸对温度的敏感性
2.5 影响生态系统呼吸速率的因素
由表3可以看出,生态系统呼吸速率与土壤总碳含量(P>0.05)、总氮含量(P>0.05)、总磷含量(P>0.05)和速效磷含量(P>0.05)的变化总体呈正相关,与土壤速效氮含量呈显著正相关关系(P<0.05).
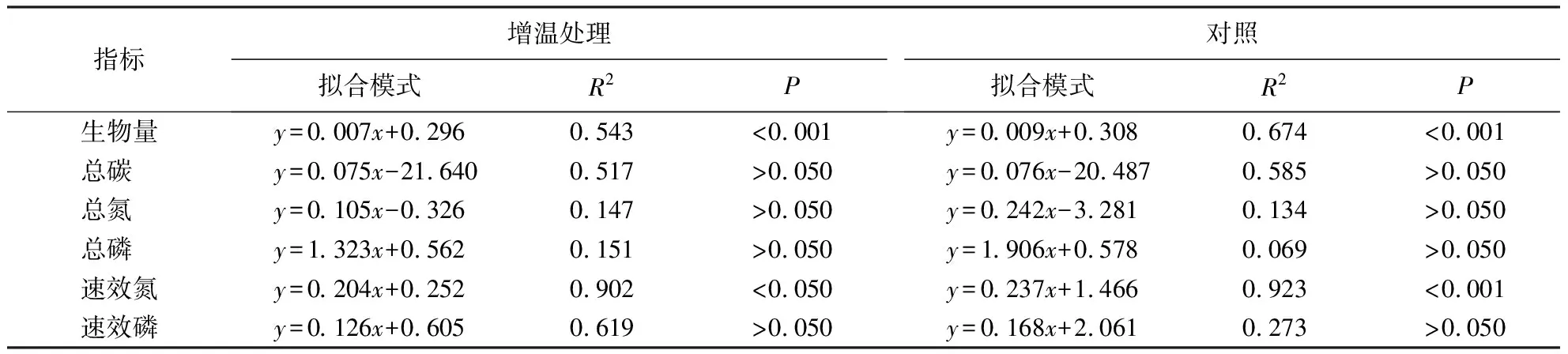
表3 土壤养分和生态系统呼吸的拟合模式
生态系统呼吸速率增量与地上生物量增量呈显著正相关(图6),地上生物量的差值越大,生态系统呼吸速率的差值就越大.
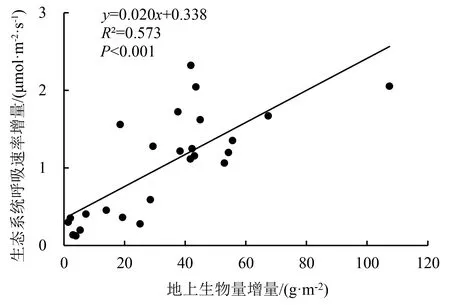
图6 增温引起的生态系统呼吸速率增量与地上生物量增量的关系
3 讨论
3.1 增温对植物生长的影响
本研究中OTC对空气的增温效果为1.2 ℃,效果显著.短期增温显著增加木里薹草植株高度(7.09%),但显著降低木里薹草群落密度(12.29%),这与徐满厚等[30]以及赵建中等[31]的研究结论相一致.温度升高对木里薹草群落密度的影响最大,这可能是由于增温后植株对地上部分生长的投入增加而降低营养繁殖的投入比例[32],地下根茎分蘖能力减弱导致木里薹草群落密度降低.OTC增温装置降低内部风速,增加立枯物量,从而削减光照[33],降低出芽率.因此,短期增温引起的木里薹草株高增加不足以弥补群落密度降低导致地上生产力变化的负效应,两者综合效应表现为生长季中后期地上生物量减少.
3.2 增温对土壤呼吸速率的影响
本研究发现:土壤呼吸速率为0.75~2.22 μmol·m-2·s-1,增温处理和对照的土壤呼吸温度敏感性分别为1.5和1.3;增温处理和对照的土壤呼吸速率分别占生态系统呼吸速率的33.24%和51.26%;短期增温没有对土壤呼吸的温度敏感性产生显著影响.Tiwari et al[7]对喜马拉雅西部高寒草甸的研究也发现,增温对土壤呼吸速率无影响.当沼泽水位降低时,土壤直接暴露在阳光和空气中,土壤温度增加使得微生物活性增强,导致土壤呼吸增强[34-35].本研究样地位于常年积水的高寒草本沼泽,受沼泽水体流动对热量交换作用以及水和泥炭高热容量属性对温度变化缓冲作用的综合影响,OTC短期增温作用对土壤温度、土壤呼吸温度敏感性和水位变化无显著影响,这可能是土壤呼吸速率强度没有发生显著变化的原因.
3.3 增温对生态系统呼吸速率的影响
本研究发现,生态系统的呼吸速率为1.18~7.94 μmol·m-2·s-1,与青藏高原高寒沼泽的研究结果基本一致[5,35-36].短期增温显著降低沼泽生态系统的呼吸速率强度(26.33%),生态系统呼吸速率与土壤速效氮含量和地上生物量呈显著正相关(P<0.05);短期增温对土壤速效氮含量无显著影响(P>0.05).地上生物量在生长季中后期显著降低(P<0.05),表明短期增温对生态系统呼吸速率的影响是由生长季中后期地上生物量显著减少引起的,这与符义稳[37]对高寒草甸碳通量的研究结果一致.这揭示了植物生物量是生态系统呼吸过程中的基质[5,38],当植被生物量呼吸占生态系统呼吸的比例较大时,植物地上部分呼吸对温度上升的响应更敏感[11].由此可知,常年积水高寒沼泽的生态系统呼吸变化主要归因于短期增温对植物地上部分呼吸的影响.
4 结论
短期增温导致木里薹草株高增加7.09%(P<0.05),群落密度降低12.29%(P<0.05).在生长季中后期(7—9月),木里薹草株高增加不足以弥补群落密度降低导致地上生产力变化的负效应.两者综合效应体现为地上生物量减少,地上生物量平均降低7.99%(P>0.05).生态系统呼吸主要受植物地上部分生物量控制.短期增温后土壤呼吸速率略有增强(P>0.05),但不足以抵消生长季中后期地上生物量显著降低(P<0.05)对地上部分呼吸速率的影响,从而导致生态系统呼吸速率降低26.33%(P<0.05).
致谢:感谢若尔盖湿地国家级自然保护区管理局为试验顺利完成提供的帮助.
