连作年限对辣椒根际土壤真菌群落结构的影响
2023-05-13刘兰英何肖云邱胤辉任丽花黄锐敏傅建炜
李 莹, 刘兰英, 何肖云, 邱胤辉, 任丽花, 黄锐敏, 傅建炜
(1.福建省农业科学院农业质量标准与检测技术研究所,福建 福州 350003;2.福建省农产品质量安全重点实验室,福建 福州 350003;3.三明市农业科学研究院,福建 三明 365000)
近年来,果蔬消费的增加及种植者对高经济效益的追求促使温室果蔬生产体系迅速发展[1].当前我国设施栽培模式多呈现出高投入和高产出,且作物类型单一化种植以及复种指数高等特点[2-4],但作物长期单一化连续种植会导致土壤退化、土传病害频发、作物产量和品质下降等连作障碍问题[3-6].研究表明,土壤环境恶化、病原菌繁殖、养分有效性失衡和自身毒性物质积累都是引发连作障碍的重要原因[5-8].目前,种植者多尝试使用化肥和杀虫剂来克服设施农业中的连作障碍,但收效甚微,且易适得其反[9].
土壤微生物是农业生态系统的重要组成部分,对土壤健康至关重要.研究表明,根际微生态系统失衡是导致作物连作障碍的最根本原因[5,10-11].作为土壤微生物区系的重要成员,真菌与其他微生物一起参与土壤生态系统中的物质循环和能量流动,在促进植物生长、维持土壤健康方面起到了重要作用[8,12],其群落结构和多样性对土壤生态系统结构的平衡及功能的发挥至关重要.研究证实,作物长期连作后,土壤微生物由细菌主导型向真菌主导型转变,而许多真菌与植物疾病密切相关[13].如辣椒和黄瓜等作物连作会导致土壤中镰刀菌的相对丰度显著增加[8],而镰刀菌是一种重要的植物病原菌.相反地,一些真菌也可以通过抑制致病菌,在植物促生防病中发挥着积极作用[14].如:在葡萄连作的过程中,许多因素都可能导致病害压力的增强,其中,真菌致病菌的增加及有益真菌群落结构的简化可能是重要原因[6];相似的结论在地黄[15]、番茄[16]和百合[17]等作物连作中也被证实.表明作物连作障碍与土壤真菌群落结构密切相关,目前已引起国内外研究者广泛关注[18].相对于土壤细菌群落,对设施栽培中作物连作导致的土壤真菌群落变化的研究仍较少.因此,研究连作管理下土壤真菌群落多样性及其演变规律,对于探索土壤可持续利用、保持土壤健康至关重要.
随着设施栽培技术的普及,辣椒设施栽培面积越来越大,但连作年限对土壤真菌群落的影响以及有益或致病真菌与土壤理化因子的关系却鲜见报道.据此,本研究以福建省三明市农业科学研究院蔬菜所辣椒育种基地为研究对象,对设施栽培条件下不同连作年限辣椒根际土壤真菌群落结构进行研究,旨在揭示土壤真菌群落结构在不同连作年限下的变化规律,探索真菌群落特征与土壤理化因子之间的关系,为辣椒种植产业可持续发展提供技术支持.
1 材料与方法
1.1 研究区概况
本研究于三明市农业科学研究院蔬菜所辣椒种植基地(117°41′E,26°15′N)进行.该区属亚热带季风性湿润气候,气温日际变化大,昼夜温差悬殊,冷暖不定,一年四季明显,雨水充沛,干湿分明,年均降雨日174 d,年日照时数1 878 h,无霜期303 d,年平均降水量1 754 mm.
1.2 试验设计
本研究选择连续种植1、3、5、10 a的大棚辣椒地为样地,辣椒品种为‘明椒系列’,不同连作年限的辣椒隶属不同的大棚,且大棚间的直线距离不超过500 m.大棚建成之前,原始土壤性质类似,大棚建成后均没有其他作物种植历史.所有大棚土壤类型均为红壤土.
辣椒栽培一年两季,种植茬口为春提早和秋延后.种植前先进行翻土,随后施用基肥,基肥为三明市森耕农业科技发展有限公司提供的有机肥.基肥主要以猪粪、鸡粪、蘑菇渣、木耳渣和辅料烟末等为原料,加入生物菌堆沤混合发酵腐熟,其中,ω(有机质)≥45%,水含量≤30%,ω(N+P2O+K2O)≥5.0%[19].通常每茬施用50 t·hm-2有机肥,待辣椒进入旺盛生长期后,每两周施用一次叶面肥(ωN∶ωP∶ωK=15∶15∶15).种植过程中均采用常规管理方式进行病害防治,所有大棚管理方式一致.
1.3 土壤样品采集
每个连作大棚被划分为3个小区(5 m×5 m),于2021年11月在辣椒盛花期,按照5点采样法,在每个小区用直径2.5 cm的不锈钢土钻在耕层(0~20 cm)采集土壤样品,取样点距辣椒根系5~8 cm.将土壤充分混合后装入无菌袋并迅速带回实验室,去除根系等杂物后,一部分样品放入冰箱(-80 ℃)保存用于土壤DNA的提取,测序工作委托北京奥维森生物科技有限公司完成.另一部分带回实验室后立即测定含水量,其余样品自然风干后进行土壤理化因子测定.不同连作年限处理下的土壤理化因子[19]如表1所示.

表1 不同连作年限处理下土壤理化因子的变化1)
1.4 土壤DNA提取、PCR扩增和高通量测序
土壤总DNA按照Mobio PowerSoil DNA Isolation Kit(100)试剂盒说明书的步骤提取,每个样品提取3个平行,分别进行PCR扩增.用1%琼脂糖凝胶电泳检测抽提的DNA的完整性,用Nanodrop 2000分光光度计(美国赛默飞世尔公司)检测DNA 的浓度和纯度,选用特异性引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2F(5′-TGCGTTCTTCATCGATGC-3′)进行PCR扩增.热启动PCR反应体系:2 μL DNA、上下游引物各1 μL、3 μL BSA、12.5 μL 2×TaqPlus Master Mix、5.5 μL ddH2O.反应程序:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸60 s,共34个循环;72 ℃延伸7 min.PCR产物经2%琼脂糖凝胶电泳检测后,采用AxyPrep DNA凝胶回收试剂盒(Axygen公司)对DNA 进行回收.PCR产物利用Illumina Miseq PE300平台进行测序.
1.5 数据处理
Illumina Miseq测序得到的双端序列数据,经过质控、拼接与去除嵌合体、短序列后得到优质序列(clean tags);利用Uparse软件在97%相似度下对优化序列进行聚类,得到操作分类单元(operational taxonomic units, OTU);采用RDP Classifier算法对OTU代表序列进行比对分析,得到每个OTU对应的物种分类信息;采用Mothur软件对全部有效序列进行OTU聚类统计,绘制韦恩图[20];使用Qiime软件计算样品的Chao1指数和香农指数等Alpha多样性指数,其中,以Chao1指数表征真菌群落的丰富度,以香农指数和辛普森指数表征真菌Alpha多样性.
采用SPSS软件进行单因素方差分析和Duncan′s多重比较(P<0.05);使用Unweighted UniFrac距离矩阵对OTU水平的群落组成结构进行非加权组平均聚类(unweighted pair-group method with arithmetic means, UPGMA)分析和非度量多维尺度(non-metric multidimensional scaling, NMDS)分析[21];使用冗余分析探索影响土壤真菌群落物种组成的关键环境因子;采用Canoco 4.5软件进行土壤真菌冗余分析和图表绘制;利用Pearson等级相关系数进行相关性分析.
2 结果与分析
2.1 不同连作年限下辣椒根际土壤真菌群落的多样性
2.1.1 Alpha多样性 本研究中,真菌基因文库覆盖率达99%,测序文库已达到饱和,同时随着测序量的增加,稀释曲线趋于平坦,表明测序数据量合理,能够准确反映各处理土壤样品的真菌群落特征.基于OTU水平的真菌群落Alpha多样性(表2)显示,随着连作年限的延长,土壤中的Chao1指数无显著差异,而连作3、5、10 a的辛普森指数和香农指数均显著低于连作1 a处理(P<0.05),表明连作3 a的真菌群落多样性较连作1 a显著下降,且逐渐趋于稳定.
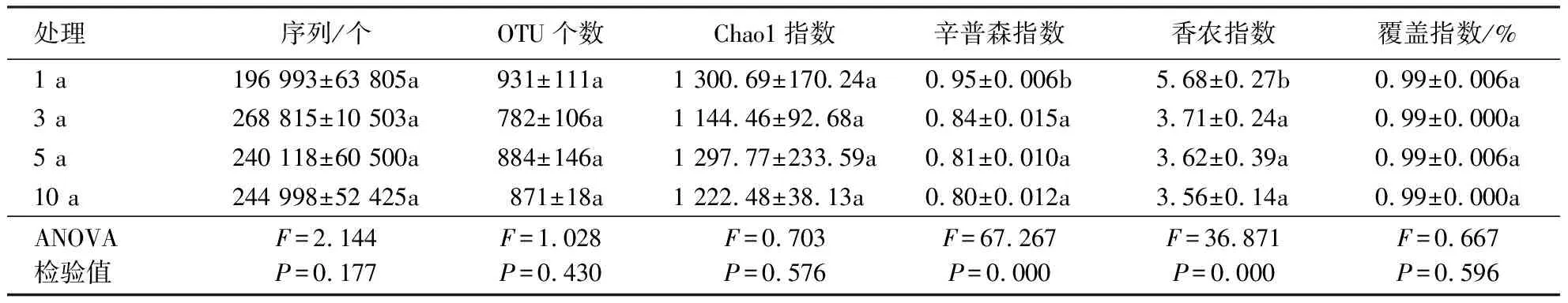
表2 不同连作年限处理下土壤真菌群落 ITS测序数据及Alpha多样性1)
图1显示:4个连作处理所共有的OTU有340个,分别占1、3、5、10 a处理OTU总数的22.68%、22.02%、22.74%和23.58%;此外,1、3、5、10 a处理中特有的OTU分别为497、296、348和307个,各占其OTU总数的33.15%、19.17%、23.28%和21.29%.表明不同连作年限土壤的OTU组成存在差异.
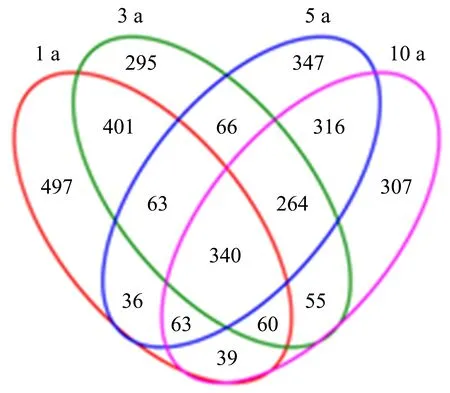
每个圈表示一个处理,重叠部分的数字表示样本之间共有的OTU个数,非重叠部分的数字表示样本特有的OTU个数.
2.1.2Beta多样性 分层聚类分析表明,5、10a连作处理土壤样品中的真菌群落更为相似且聚类,与1、3a连作处理的真菌群落明显分离(图2A).NMDS分析(胁强系数=0.003 6)显示,所有样品分为3类,来自5、10a连作处理的样品高度相似并聚集,与1、3a连作处理的样品分离(图2B),此结果与层次聚类分析结果相似.表明连作改变了土壤真菌群落结构,且不同连作年限下真菌群落结构的差异显著.
2.2 不同连作年限下辣椒根际土壤真菌群落的结构和组成
4个连作处理土壤共鉴定出17个真菌门类.其中,子囊菌门(Ascomycota)是4个处理的绝对优势菌群(图2A),占比达到了79.10%~95.12%;担子菌门(Basidiomycota)为亚优势菌门,其相对丰度为1.50%~13.54%;被孢霉门(Mortierellomycota)和壶菌门(Chytridiomycota)的占比分别为0.34%~3.64%和0.60%~11.34%;梳霉门(Kickxellomycota)在5a连作处理中的相对丰度达1.0%,但在其他3个处理中几乎没有检测到.
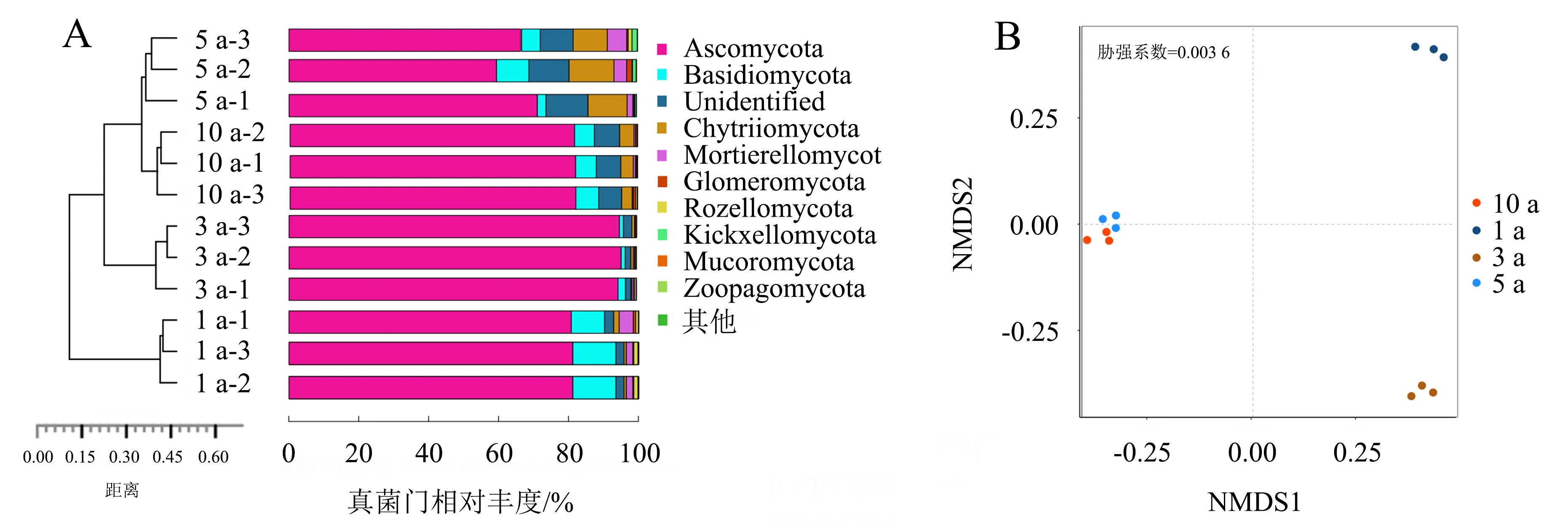
图2 基于Unweighted UniFrac距离的 UPGMA聚类树(A)和NMDS分析结果(B)
在属水平上对真菌进行物种注释分析,共获得18个优势属群(平均相对丰度>1%)(表3).其中,子囊菌门中的Ovatospora、镰刀菌属和毛壳菌属是4个连作处理共有的优势真菌属.对比发现,枝葡萄孢属、木霉属、伽穆孢属和白鬼伞属随着连作年限的延长而逐渐下降,而矮菇属、枝顶孢霉属、帚霉属和Alphamyces则呈相反趋势;被孢霉属在连作1~5 a的相对丰度稳定,但在连作10 a后显著下降;粪壳菌属、麻孢壳菌属、Ovatospora和毛壳菌属随着连作年限的延长呈先显著增加后明显下降的趋势;而镰刀菌属和曲霉属随着连作年限的延长呈现出波浪形的变化趋势.值得注意的是,在真菌类群中有一大部分仍未被分类,在4个处理中的占比达到了20.08%~31.67%,这表明可能还有很多对连作响应敏感的类群有待探明.
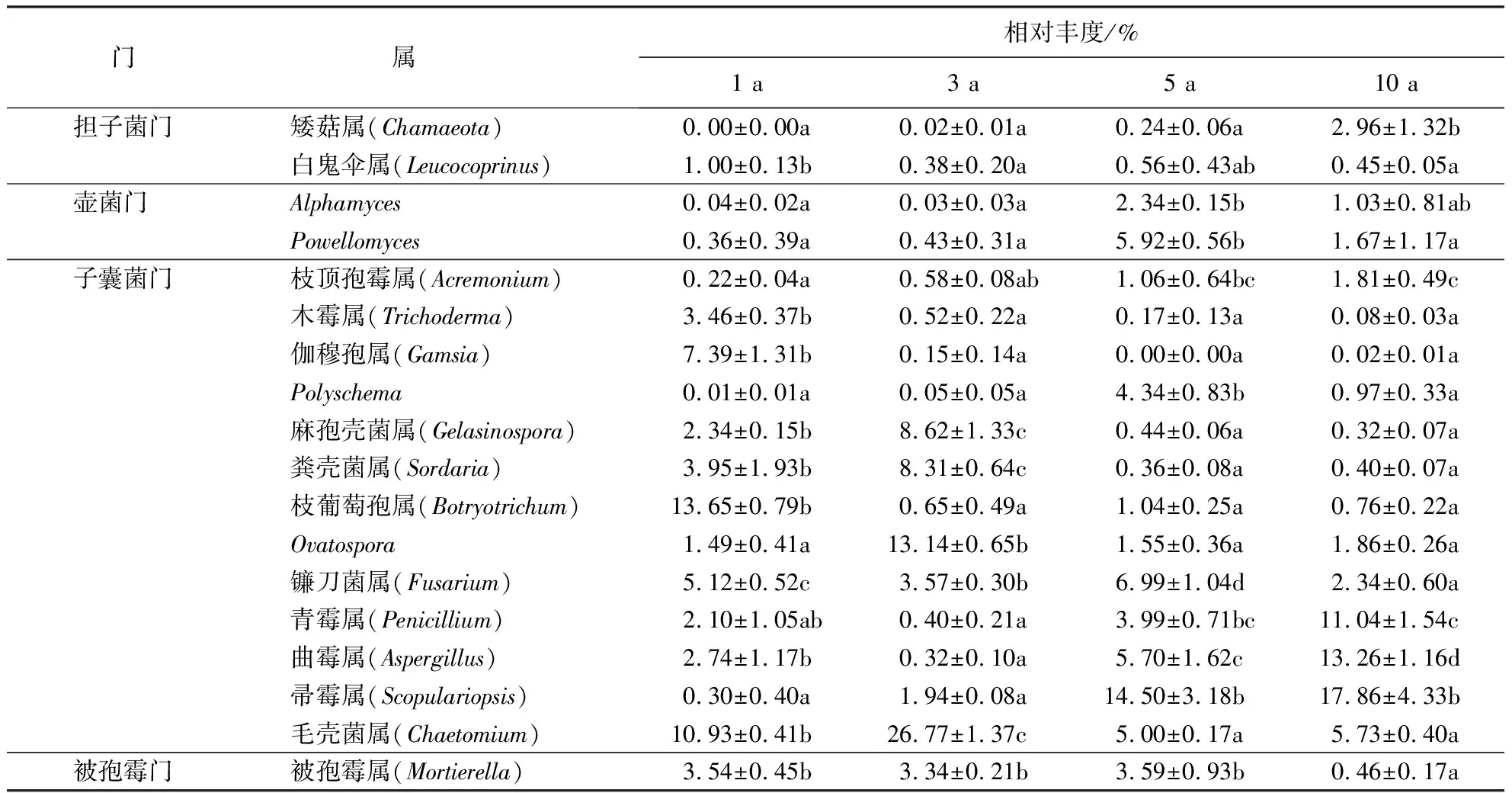
表3 不同连作年限处理下土壤真菌属水平的组成和分布1)
2.3 真菌群落结构与土壤理化因子间的相互关系
Pearson相关性分析(表4)表明,香农指数、辛普森指数与土壤pH均呈极显著负相关(r值分别为-0.846、-0.816),与土壤可溶性有机碳的质量分数呈显著负相关(r值分别为-0.597、-0.625).

表4 真菌群落多样性指数与土壤理化因子间的相关系数1)
以不同连作年限处理的OTU水平作为响应变量,研究土壤真菌群落结构与土壤理化因子的关系.结果(图3)显示,冗余的前2个轴解释了真菌群落69.1%的变异,表明第1、2排序轴能较好地反映出真菌群落结构与土壤理化因子之间的相互关系.其中,3、1 a连作处理组分别分布在第1排序轴的两侧,5、10 a连作处理组聚为一簇,与1 a连作处理组分别分布在第2排序轴的两侧,与Beta多样性的结果一致.被选择的土壤理化因子中,pH及有机质、有效磷、速效钾的质量分数对真菌群落结构的影响较大,是影响真菌群落组成的重要因素.其中,pH主要影响3 a连作土壤的真菌群落结构,有机质、有效磷和速效钾的质量分数则主要影响5、10 a连作土壤的真菌群落结构.速效钾、有效磷和有机质质量分数矢量箭头之间的夹角为锐角,表明这3种理化因子之间可能具有协同效应.

RDA1、RDA2分别表示第1、2排序轴.EC:电导率;C/N:碳氮比;铵态氮(硝态氮);DOC:ω(可溶性有机碳);AP:ω(有效磷);AK:ω(速效钾);SOM:ω(有机质).
3 讨论
3.1 连作年限对辣椒根际土壤真菌群落多样性的影响
土壤微生物的多样性被认为是维持土壤健康的关键[22].本研究中,尽管连作10 a下辣椒根际土壤的真菌丰富度与连作1 a的差异不显著,但连作导致的真菌群落多样性显著降低;与之相似,黑胡椒、丹参、咖啡和黄瓜在长期连作后,其根际土壤真菌的香农指数显著下降[3,22-23];与之相反,草莓根际土壤真菌群落多样性随着连作年限的延长而增加[24].连作对真菌群落多样性的影响可能与作物种类、管理措施、种植土壤和连作年限等因素有关[24].本研究中,化感物质的累积以及辣椒根系分泌物长期释放的生态效应,可能会促进某些特定微生物富集[25];同时,多年相同的田间管理模式为特定真菌提供了优良的繁殖环境,但也导致部分微生物在生物竞争中减少甚至消失,这些都可能是长期连作后真菌群落多样性下降的重要原因[26].本研究的Beta多样性显示,连作年限显著影响辣椒根际土壤真菌群落的多样性,此结果与之前对细菌群落多样性的分析结果[19]有所差异.3、5 a连作处理组的土壤真菌群落分散分布;相反,5、10 a连作处理组的真菌群落聚集,表明连作5 a后,真菌群落结构趋于稳定.
3.2 连作年限对辣椒根际土壤真菌群落结构的影响
本研究中,子囊菌门是4个连作处理的绝对优势菌群.子囊菌门多为腐生营养型真菌,是土壤最重要的有机质分解者[27].研究表明,子囊菌门更容易在高肥力的土壤中富集[28].但在本研究中,子囊菌门的相对丰度随着连作年限的延长呈现出剧烈的波浪形变化,与土壤无机氮和有效磷等养分含量的变化未呈现出明显的一致性,这可能与子囊菌门有较高的物种多样性和进化速度有关.
真菌在土壤环境中起着重要作用,其与农作物病害密切相关[29].其中,归属于子囊菌门的优势属——镰刀菌属是土传病害中主要的病原菌属[6],会导致辣椒患枯萎病、根腐病、叶枯病和果腐病等[30].一般来说,连作极易造成土壤同质性,导致潜在病原菌的累积.本研究中,镰刀菌属的相对丰度在连作5 a时达到最高,但在连作10 a时的相对丰度较其他处理显著降低,此结果与葡萄长期连作后的结果[6]一致.土壤中致病真菌含量的增加会导致土传病害发病率升高.据了解,10 a样地在连作5~10年期间曾暴发大规模重的土传病害,管理者进行了大规模的药物消杀,可能导致后期(连作10 a)病原体减少[19].这与本课题组前期在对细菌的研究中所发现的生防菌链霉菌属(Streptomyces)和鞘氨醇单胞菌属(Sphingomonas)相对丰度在连作5 a后急剧增加的趋势相吻合.本研究中,土壤枝顶孢霉属、青霉菌属和曲霉菌属等生防菌的相对丰度在辣椒连作5 a后显著增加,连作10 a时的相对丰度分别是连作1 a时的8.23、5.26和4.84倍.枝顶孢霉属和青霉菌属均属于腐生营养型真菌,可以通过分泌某些酶来抑制病原细菌和真菌;青霉菌属对纤维素和半纤维素有很强的降解能力,对植物有促生作用,对多种植物致病真菌也具有抑制或防治作用[31-34];曲霉菌属的代谢产物能有效抑制27种农作物病原菌的生长,是抑制青枯病的拮抗菌[35].研究表明,在纤维素降解和腐殖质形成中发挥重要作用的帚霉属的相对丰度在连作5 a时显著增加[36].植物可以通过招募保护性微生物抑制植物病原体来保护其免受侵害[6,37].以上结果表明,有益真菌在连作5 a时的相对丰度显著增加可能是植物在暴发严重病害后的自我调节防御机制,镰刀菌属的相对丰度在连作5 a时也下降可能与有益微生物丰度的变化有关.相反地,毛壳菌属的相对丰度在连作5、10 a时显著下降;连作1 a时的木霉菌属占比为3.46%,但随着连作年限的延长,该菌群几乎监测不到.相似地,被孢霉属的相对丰度在连作1~5 a时稳定在3%,但在连作10 a时不到1%.毛壳菌属产生的毛壳菌素能够有效抑制多种病原真菌的繁殖[6,38];而木霉菌属被认为是具有重寄生功能的生防菌[29];被孢霉属具有潜在分泌抗菌素和抑制部分病原菌生长的能力,被确定为对土壤有益的微生物[39-40].这些潜在的植物有益真菌,其丰度的下降可能与辣椒连作障碍密切相关.研究表明,植物生长状况和连作障碍可能取决于致病微生物与有益微生物之间的竞争[6].辣椒长期连作后,部分生防菌的减少和病原菌的积累可能是造成辣椒连作障碍的原因之一.
3.3 土壤理化因子与真菌群落结构之间的关系
真菌群落多样性与pH呈显著负相关,这与“土壤pH过高会限制真菌存活”[41]的结果一致.土壤pH下降会刺激嗜酸性真菌繁殖[6];同时,土壤中的真菌大多为异养微生物,需要外界碳分解为其提供养分和能源.但微生物对碳有效性的需求和适应能力存在差异,连作后可溶性有机碳含量的提高可能会导致适宜在低浓度碳环境下生存的真菌的减少,从而促进适宜在高浓度碳环境下生存的真菌的繁殖,导致真菌群落多样性下降.而微生物多样性的减少是土传病害发生的重要原因.
真菌的生长和繁殖依靠土壤的养分.本研究冗余分析表明,土壤pH及有效磷、速效钾、有机质含量对土壤真菌群落组成的影响较大.其中,连作3 a时的真菌群落结构主要受pH影响,而连作5、10 a时的真菌群落结构则主要受有效磷、速效钾和有机质含量的影响.表明在短期连作过程中,pH是影响真菌群落结构的主要因素,这可能是由于本研究施用的有机肥呈碱性,在短期连作下pH呈升高的趋势,对真菌群落结构的影响更明显.但随着连作年限的延长,土壤养分的累积,有效磷、速效钾和有机质逐渐成为影响真菌群落结构的重要因子.此结果与Zhao et al[3]的研究结果相似.Zhao et al[3]在温室条件下对黄瓜进行连作处理,发现土壤养分对土壤微生物群落结构的影响可能比pH更大.
4 结论
辣椒根际土壤在长期连作后,其真菌群落多样性随着连作年限的延长而显著降低;连作显著改变了真菌群落结构,不同连作年限处理间的群落结构存在显著差异;长期连作导致部分有益真菌减少、病原真菌增多,而这主要与pH及有效磷、速效钾、有机质含量密切相关.表明辣椒在长期连作下,合理的养分类型和数量对平衡土壤真菌群落结构、提高土壤健康、降低植株发病率至关重要.
