经颅交流电刺激对阿尔兹海默病小鼠双侧海马有效连接的干预研究
2023-05-05曹甜甜王亚飞张文龙吴林彦黄亮李四楠李龙刘天王珏
曹甜甜,王亚飞,张文龙,吴林彦,黄亮,李四楠,李龙,刘天,王珏
(1. 西安交通大学生命科学与技术学院,710049,西安;2. 榆林市第一医院神经外科,719000,陕西榆林)
阿尔茨海默症(AD)是一种神经退行性疾病,患病率逐年上升。阿尔茨海默症主要表现为认知下降、工作记忆损伤和行为改变等问题,影响患者的正常生活[1]。
在网络层面研究脑网络有效连接记忆机制,可以更好地反馈脑区之间的信息流向和因果关系[2],对脑功能障碍的研究具有重要意义[3]。格兰杰因果分析可以研究大脑不同区域之间的有效连接。有研究表明,与正常对照组相比,AD患者表现出较低的网络连接特性,AD患者脑区之间的连接存在异常[4-5]。AD发病的潜在机制尚不清楚,有研究推测,AD患者认知功能障碍可能与脑区之间连接受损有关[6-8]。
海马在研究工作记忆与学习过程中起着重要的作用。有研究表明,AD患者的海马萎缩程度发生左、右不对称变化,左海马比右海马发生更大程度的萎缩[9-10],左侧海马代谢物的变化与AD患者的记忆障碍有关[11]。推测AD患者存在的记忆缺陷可能与左、右侧海马有关,目前针对AD患者左、右海马之间连接的研究工作相对有限,左、右海马之间有效连接的研究对于了解AD发病机制和改善AD症状具有重要的意义。
经颅交流电刺激(tACS)是一种无创脑刺激技术,通过在头皮表面施加低振幅交流电刺激可调节脑部振荡节律,改善认知功能[12]。多项研究表明,在AD小鼠模型[13]和AD患者[14]中均表现出γ振荡(30~80 Hz)受损。研究发现,40 Hz的视觉刺激诱导γ振荡可以促进小胶质细胞的形成,减少AD小鼠模型中Aβ蛋白和p-tau蛋白的沉积[15]。40 Hz的tACS可以改善AD患者的记忆和认知功能[16-17]。上述研究表明,40 Hz的tACS可能是一种有前景的干预方法,可以改善AD的认知功能。有文献基于因果分析方法,探究了体感电刺激对正常人体脑网络活动的影响[18]。目前,tACS干预AD的研究大多局限于刺激靶区处的局部脑区,对相关脑区的联合分析较少。
在本研究中,通过tACS刺激小鼠右侧海马,基于格兰杰因果分析方法,研究tACS对AD小鼠双侧海马之间有效连接的影响。研究结果显示,40 Hz tACS可以增强AD小鼠双侧海马神经元电活动的功率谱密度,改善AD左、右海马之间异常的有效连接。本文为tACS干预AD的研究提供了参考和依据。
1 材料与方法
1.1 实验对象
实验选用16只3月龄小鼠,其中12只AD小鼠模型为APP/PS1转基因小鼠,4只普通小鼠为C57/BL6-Tg小鼠,均购自中国医学科学院医学实验动物研究所。所有小鼠被饲养在温度适宜的房间内。 将12只AD小鼠分为3组:AD组、tACS组和假刺激组,每组4只小鼠,选用4只普通小鼠作为实验的正常对照组。
1.2 经颅交流电刺激实验
tACS组进行刺激时,将小鼠用2%异氟烷麻醉后固定在立体定位仪上,通过外科手术方法切开头皮,刮除筋膜裸露头骨,以小鼠颅骨的前卤作为参照点,将电极贴附在海马位点(前囟向后2 mm,矢状缝左右1.35 mm)的正上方区域,刺激右侧海马。小鼠连续21 d每天接受20 min的tACS刺激:刺激电流为130 μА,频率为40 Hz,通过函数波形信号发生器(RIGOL DG4202)输出交流电信号。通过示波器实时记录小鼠接受的刺激信号,保证输出刺激信号的准确性。假刺激组是对APP/PS1转基因小鼠仅在刺激实验开始后30 s和结束前30 s进行刺激。实验流程如图1所示。 tACS组和假刺激组对小鼠进行21 d的连续刺激,正常对照组和AD组未进行刺激操作。刺激完成后,所有小鼠均植入记录电极,手术后小鼠恢复3 d,然后进行Y迷宫的任务训练和数据记录。

图1 小鼠tACS刺激实验流程图
Fig.1 Time course of experimental procedures of tACS on rat
1.3 记录电极放置
将小鼠用2%异氟烷麻醉,通过外科手术方法切开头皮,刮除筋膜裸露头骨,以小鼠颅骨的前卤作为参照点,确定记录电极位点(前囟向后2 mm,矢状缝向左1.35 mm和向右1.35 mm),用颅骨钻开颅并用立体定位仪植入记录电极(植入深度为1.29 mm)。其中记录电极采用镍铬直丝,每个脑区3根直丝,每个电极由6根直丝构成。小脑处植入颅骨钉作为参考电极,用铜网包围整个采集电极作为屏蔽。手术后恢复3 d时间,根据手术的损伤情况,在进行Y迷宫训练之前,剔除掉行为异常的老鼠。
1.4 行为训练
小鼠进行Y迷宫行为训练,同时记录小鼠的行为学表现和电生理数据。Y迷宫由3个臂(Ⅰ区、Ⅱ区和Ⅲ区)组成,如图2所示,小鼠被允许自由探索3个臂,每次实验包括两次训练,每只老鼠重复85次实验。小鼠放置在起始端(Ⅰ区),第一次是自由选择阶段,可以任意选择Ⅱ或者Ⅲ区,然后小鼠自行返回起始端(Ⅰ区)。第一次训练结束10 s后,开始第二次训练,要求小鼠进入与自由选择阶段相反的臂。若第二次选择与第一次相同,则认为是一次错误的训练,在小鼠选择错误的Ⅱ区或者Ⅲ区的臂底施加弱脉冲刺激小鼠1 s,弱电流刺激小鼠产生的刺激痛感作为选择错误的惩罚。若第二次与第一次选择不同则认为是正确的训练,允许小鼠正常返回。在Y迷宫的交叉口,记录一次实验的起始时间,当小鼠进入Y型迷宫Ⅱ区或者Ⅲ区辨别的决策点时,再次记录实验的时间,两次时间点之差表示小鼠一次工作记忆训练的时间。

图2 Y型迷宫训练任务示意图Fig.2 Schematic of the training task on Y-maze
1.5 数据记录与分析
1.5.1 数据记录
局部场电位记录:在进行Y迷宫行为训练的过程中,通过多通道生理采集系统cerebus记录左海马和右海马两个通道的神经电活动信号,对原始信号进行采样,采样率设为500 Hz。同时,记录小鼠在Y迷宫任务期间的85次行为训练的情况,观察训练的正确率与训练次数之间的关系,训练次数以步长为5的窗口滑动,计算并记录每10次训练正确率的平均值。
1.5.2 功率谱密度
使用matlab中pwelch函数,计算脑电信号的功率谱密度,脑电信号采样率为500 Hz,设置Hamming窗,窗长1 s,50%的重叠。计算正常对照组、AD组、tACS组和假刺激组脑电信号的功率谱密度,绘制功率谱密度曲线,频率设置为0~45 Hz(主要研究频率范围为0~45 Hz)。
1.5.3 格兰杰因果分析
格兰杰因果分析就是基于自回归模型分析时间序列X的过去是否有助于预测时间序列Y的现在。假设x1(t)和x2(t)是任意两个广义平稳的时间序列,x1(t)或x2(t)单独可以表示为

(1)
式中:p是模型阶数;aj是回归系数;ξn为时间序列的预测误差。通过x2(t)来预测x1(t)和通过x1(t)来预测x2(t)可表示为
(2)
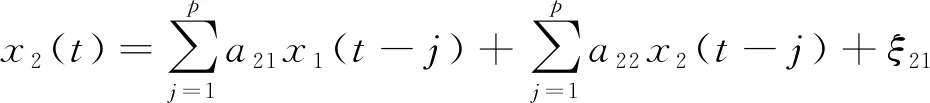
(3)
式中:ξ12和ξ21为时间序列的预测误差。从x1(t)到x2(t)的因果作用可表示为

(4)
式中:var(ξ1)和var(ξ12)是单变量和全模型中的预测误差。
本研究使用matlab工具箱(MVGC)分析格兰杰因果关系以及海马两个区域的因果关系,自定义阶数为50,计算θ频段和γ频段下双侧海马之间的格兰杰因果值。
1.6 统计学方法
采用GraphPad prism软件进行统计学分析,通过独立样本t检验和双因素方差分析,比较正常对照组、AD组、tACS组和假刺激组在不同频带下的统计学差异。统计分析中P表示显著性水平,*表示P<0.05,**表示P<0.01,***表示P<0.001,****表示P<0.000 1,ns表示无显著性差异,数据表示为均值±标准误差(SEM)。
2 行为及电生理结果
本文统计了正常对照组、AD组、tACS组和假刺激组小鼠在执行Y迷宫任务时的训练表现。计算各个组的局部场电位在主要频带上(0~45 Hz)的功率谱密度,绘制功率谱密度与频率的曲线,应用格兰杰因果的方法,计算各个组的左、右海马之间的格兰杰因果连接值(GC),分析各组之间脑网络有效连接的差异。
2.1 行为表现
图3记录了Y迷宫训练任务期间不同组小鼠的行为表现。随着训练次数的增加,各组小鼠的训练正确率都有明显增加,AD组的平均正确率(0.556±0.02)明显低于正常对照组的正确率(0.825±0.08),tACS组行为正确率(0.643±0.05)明显提高,在训练次数大于30次时接近正常对照组水平。

图3 各组小鼠在Y迷宫任务中的行为表现 Fig.3 Behavioral correct rate on the Y-maze task in each group of rat
2.2 功率谱密度分析
图4为正常对照组、AD组、tACS组和假刺激组的功率谱密度曲线,计算了在θ频段(4~12 Hz)和γ频段(30~45 Hz)的平均功率谱密度。在θ频段上,AD组的平均功率谱密度显著低于正常对照组(P<0.01),而tACS组的功率谱密度恢复至正常对照组的值。在γ频段上,AD组的功率谱密度显著低于正常对照组(P<0.000 1),tACS组的功率谱密度提高至正常对照组水平。tACS可以增强AD小鼠海马处θ频段和γ频段的功率。

(a)θ频段

(b)γ频段
2.3 θ频段和γ频段的格兰杰因果分析
行为学表现和功率谱密度分析表明,AD组与正常对照组之间存在差异,tACS可以恢复这种差异。使用格兰杰因果分析来进一步研究正常对照组、AD组、tACS组和假刺激组双侧海马之间的因果连接。
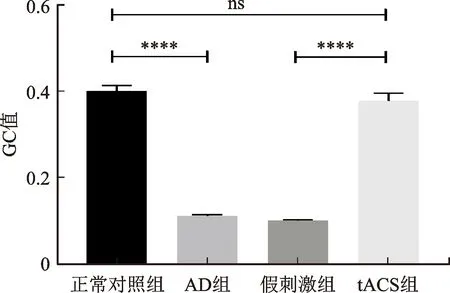
在θ频段上,统计学分析各组双侧海马之间平均GC值的差异(见图5)。在左海马到右海马的信息流向上,4个组的平均GC值分别是0.400±0.013、0.111±0.002、0.377±0.017、0.099±0.002(见图5(a))。在右海马到左海马的信息流向上,4个组的平均GC值分别是0.208±0.007、0.109±0.003、0.185±0.005、0.044±0.002(图5(b))。从左海马到右海马和从右海马到左海马的信息流向上,AD组的GC值均显著小于正常对照组(P<0.000 1),tACS组的GC值均显著大于假刺激组的GC值(P<0.000 1)。在左海马到右海马的信息流向上,tACS组的GC值与正常对照组无统计学差异,在右海马到左海马的信息流向中,tACS组的GC值也接近正常对照组水平。图5(c)是分析各个组双侧海马之间不同信息流向的统计学差异。正常对照组的GC值在左、右海马之间不同的信息流向上存在统计学差异(P<0.000 1),AD组和假刺激组的GC值在双侧海马的不同信息流向上无统计学差异。与正常对照组相比,AD组从左海马到右海马GC值的变化是大于右海马到左海马的GC值变化程度(0.289>0.099),而tACS组在不同的信息流向上GC值的差异与正常对照组相近。
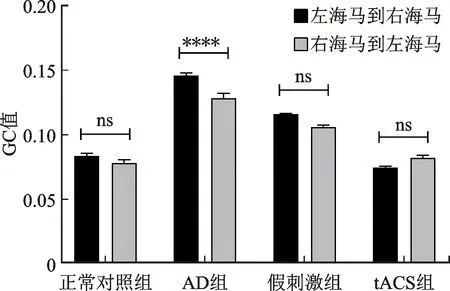
图6是γ频段上正常对照组、AD组、tACS组和假刺激组的平均GC值比较,在左海马到右海马的信息流向上,4个组的平均GC值分别是0.083±0.002、0.145±0.002、0.074±0.001、0.115±0.001(见图6(a))。在右海马到左海马的信息流向上,4个组的平均GC值分别是0.077±0.002、0.127±0.003、0.082±0.002、0.105±0.001(见图6(b))。从左海马到右海马和右海马到左海马的信息流向上,AD组的GC值均显著大于正常对照组(P<0.000 1),tACS组的GC值均显著小于假刺激组的GC值(P<0.000 1),tACS组的GC值与正常对照组之间无显著性差异,tACS刺激使AD小鼠的GC值恢复到正常对照组水平。图6(c)是分析各个组左、右海马之间不同信息流向上的统计学差异。AD组的GC值从左海马到右海马与右海马到左海马的信息流向存在统计学差异(P<0.000 1),其他组双侧海马之间不同的信息流向无统计学差异。与正常对照组相比,AD组从左海马到右海马的GC值比右海马到左海马的GC值变化程度更大(0.062>0.05),tACS组在不同的信息流向上GC值的差异与正常对照组相近。
(a)左海马到右海马

(b)右海马到左海马

(c)双侧海马之间

(a)左海马到右海马

(b)右海马到左海马
(c)双侧海马之间
3 讨 论
本文记录了正常对照组、AD组、tACS组和假刺激组在Y迷宫任务期间左、右海马的局部场电位。通过行为表现,功率谱密度分析和左、右海马之间的格兰杰因果分析的研究结果发现:①AD小鼠行为训练的正确率低于正常小鼠,在θ和γ频段上的功率谱密度低于正常小鼠;②AD小鼠的双侧海马之间有效连接异常;③tACS干预后,AD小鼠的行为正确率提高,θ振荡和γ振荡的功率增强,且tACS改善了AD小鼠海马双侧的异常脑网络连接。
在Y迷宫行为训练期间,AD组训练正确率明显低于正常对照组,表明AD小鼠工作记忆损伤,这与已有的研究一致[19]。tACS可以提高AD小鼠行为训练的正确率,推测tACS可以改善AD小鼠的记忆损伤。
海马在学习和记忆中起着重要作用,因此本文分析了各组小鼠在执行工作记忆任务期间海马处的功率谱密度,进一步探究AD小鼠记忆损伤的神经机制。研究发现,与正常对照组相比,AD组的θ频段和γ频段的功率谱密度均降低。40 Hz tACS增强了γ振荡和θ振荡的能量,推测40 Hz tACS增强了γ振荡,同时调制了θ振荡,对θ频段振荡有促进作用。之前有研究发现γ频段的tACS会调制β的振荡[20],表明tACS可能通过调节不同频段振荡来改善AD的记忆功能。
海马处异常的θ和γ振荡可能是阿尔兹海默症的重要特征。目前,在AD小鼠模型中已经发现γ活性降低[13],同时AD患者中也存在γ振荡受损[14]。γ振荡(30~100 Hz)与学习、记忆等认知活动有关,主要反映抑制性中间神经元和锥体细胞之间的信息传递。视觉刺激诱导神经退行性疾病小鼠模型中的γ活性可以改善突触功能,改善小鼠的行为表现,防止神经退行性病变[21]。AD小鼠γ振荡能量异常可能影响双侧海马神经环路的信息整合。θ振荡(4~12 Hz)与探索行为有关,在工作记忆期间发挥重要作用,θ振荡反映了记忆信息的处理和存储[22]。海马处的θ振荡和γ振荡受损可能是AD小鼠工作记忆受损的神经机制之一。
行为学表现和功率谱密度分析之后,通过格兰杰因果分析,研究海马左、右侧之间的有效连接,从脑网络连接的角度更好反馈记忆信息传递机制。研究发现,在θ频段下,AD组左、右海马之间的GC值均小于正常对照组,在γ频段下 AD组左、右海马之间的GC值均大于正常对照组,表明在θ和γ频段上AD小鼠双侧海马之间的有效连接存在异常,这与之前的研究一致[23]。AD患者的脑网络有效连接存在异常,脑网络连接存在异常可能与AD患者大脑区域之间的信息交换效率较低有关[23]。XUE等研究表明,静息态下正常对照组、轻度认知障碍组和AD受试者的海马体与其他大脑区域之间网络连接存在差异,这种现象可能与认知功能有关,且异常连接是单向的[24]。大脑网络有效连接的异常变化可能与神经退行性过程有关,也可能涉及海马及其他大脑区域。支持学习和记忆的信息传输通路异常可能导致认知障碍和记忆损伤,AD小鼠双侧海马的有效连接异常可能影响到记忆信息的传输,影响到AD患者的行为表现。双侧海马之间有效连接异常可能是AD小鼠工作记忆受损的神经机制之一。与正常对照组相比,AD组从左海马到右海马信息流向上的GC值比右海马到左海马的GC值变化程度大,AD组双侧海马之间的网络连接异常,存在左、右不对称性。Watanab等研究表明,轻度认知障碍和AD患者双侧海马中的代谢物浓度发生不对称变化,左侧海马代谢物浓度变化与轻度认知障碍和AD患者的记忆功能障碍有关[11]。这与本文的结果一致。
已有多项研究表明,tACS可以改善AD患者的认知功能。Moussavi等研究表明,tACS可以改善老年痴呆患者的认知功能,可能对长期记忆有改善作用[25]。Dhaynaut等研究表明,tACS靶向的颞叶区域的p-tau蛋白沉积显著减少,40 Hz的tACS可以改善AD患者的症状[26]。
目前,tACS干预AD的研究大多限于刺激靶区相关的局部脑区的研究,本研究联合分析海马靶点区和靶点对侧之间的有效连接,进而揭示tACS干预AD的神经机制。研究表明,tACS恢复了AD小鼠双侧海马之间异常的有效连接,tACS靶点区和靶点对侧之间的有效连接均使异常的有效连接恢复至正常水平。海马是一个有机整体,尽管左、右海马之间在解剖上[27-28]和功能[29]上存在一定的差异,但tACS靶向刺激右海马后,从右海马到左海马的信息流向与左海马到右海马的信息流向上的有效连接发生一致变化,左、右海马之间的连通性变化是一致的。这表明,40 Hz tACS可以通过改善AD双侧海马之间的有效连接来改善AD小鼠的记忆功能。
在后续研究中,需要进一步探究文中结果确切的神经生物学机制。本文没有单独考虑训练任务正确和错误情况下的脑网络连接,后续将继续研究训练任务的正确与否对小鼠双侧海马网络连接的影响。本研究存在样本量较小的限制,后续通过增加样本量来进一步探究tACS的效果和长期作用。
4 结 论
多项研究表明,tACS对AD患者的症状有改善作用,针对tACS干预AD的神经机制的研究仅限于刺激靶点相关的局部脑区。本文采用格兰杰因果分析的方法,研究海马靶点侧和靶点对侧的脑网络有效连接,从相连脑区的角度探究tACS干预AD的神经机制。研究发现,AD小鼠双侧海马之间的网络连接出现异常,而以右海马为靶点的40 Hz的tACS通过改善双侧海马间的有效连接,可以恢复海马区γ和θ的异常振荡,改善了AD小鼠在记忆任务中的表现。tACS可以恢复阿尔兹海默病存在的异常脑网络连接,从而改善AD患者的认知功能,为AD的治疗提供一种神经调控方法。
