线粒体对肿瘤微环境中肿瘤细胞和免疫细胞代谢的影响
2023-03-23胡雪宁林冬静
田 静,佘 威,胡雪宁,王 爽,崔 莹,林冬静
(吉林医药学院:a.临床医学院,b.肿瘤靶向治疗与转化医学重点实验室,吉林 吉林 132013)
2020年,世界卫生组织国际癌症研究机构发布全球最新癌症负担数据,我国癌症负担新发人数占全球23.7%[1],远超其他国家,综合干预和靶向治疗仍是癌症研究的重点。
在肿瘤微环境(tumour microenvironment,TME)中,细胞代谢是维持活力和功能的关键因素,可作为肿瘤靶向治疗的重要方向。肿瘤细胞强烈依赖葡萄糖酵解途径获取能量。在代谢过程中,大量的氧化中间产物的积累导致戊糖磷酸活性升高,同时核糖五磷酸的产量大幅度提升,从而加速癌细胞的分裂。癌细胞快速生长消耗大量活性氧(reactive oxygen species,ROS),使大量乳酸堆积,它有能力在这种低氧、酸性微环境中生长,可能耗尽周围大量浸润免疫细胞所需要的关键营养物质,抑制免疫检查点对癌细胞的破坏[2]。在研究肺腺癌组织标本时,发现了肿瘤微环境中肿瘤细胞与免疫细胞相互调节机制[3]。
线粒体代谢为细胞代谢网络核心,如氧化磷酸化、三羧酸循环及氨基酸代谢等代谢过程都位于线粒体内。线粒体ROS家族的产生和氧化还原反应再平衡,对癌细胞增殖和侵袭起着刺激作用。研究人员发现,代谢能力与免疫水平呈正相关,T细胞的持久性随着线粒体生物量和备用呼吸能力的增加而增加,二者的增加为防止癌细胞的复发和根除癌细胞提供了必要的生物能量优势[4]。
本综述旨在讨论线粒体变化对TME中肿瘤细胞与免疫细胞代谢的影响,进而分析肿瘤细胞代谢重编程和抗肿瘤免疫反应的联系,前者如何调节后者将为抗肿瘤免疫治疗中以线粒体为目标靶点研究提供参考。
1 TME中肿瘤细胞和免疫细胞代谢主要特征
TME是指肿瘤发生、生长、转移与肿瘤细胞所在的复杂的内外环境,包含多种细胞和其他成分。肿瘤细胞利用葡萄糖酵解成乳酸,也被视为“代谢的重新编程”,拥有六大特性[5]:葡萄糖和氨基酸的摄取、失常;养分获得路径的投机性;利用糖酵解中间路径所形成的产物合成蛋白质、核酸及脂类;氮源供给增加;代谢产物启动基因表达失控;代谢产物与TME相互效用。
肿瘤细胞代谢异常突出体现为葡萄糖代谢的异常,尤其是三个与糖酵解关键酶(己糖激酶、磷酸果糖激酶和丙酮酸激酶)活性加强与同工酶谱的变更、糖异生关键酶活力下降有关。糖酵解许多中间路径所形成产物被肿瘤细胞应用,导致生成蛋白质、核酸和脂类增加,进而加速了肿瘤细胞的增进。另外,谷氨酰胺是人体血浆中含量最丰富的氨基酸,为嘌呤、嘧啶核苷酸的合成提供氮源,参与核苷酸生物合成,是谷胱甘肽合成必不可少的原料,参与维持ROS稳定。肿瘤细胞中谷氨酰胺的摄取和分解代谢显著增强,存在“谷氨酰胺依赖”现象[6]。
TME内有免疫激活细胞群和炎性细胞群,还有免疫抑制细胞群(表1)[5]。这些细胞可以通过分泌细胞因子和扰乱代谢,抑制或消除抗肿瘤效应细胞的功能(图1)[5]。

图1 TME中的癌细胞和免疫抑制代谢
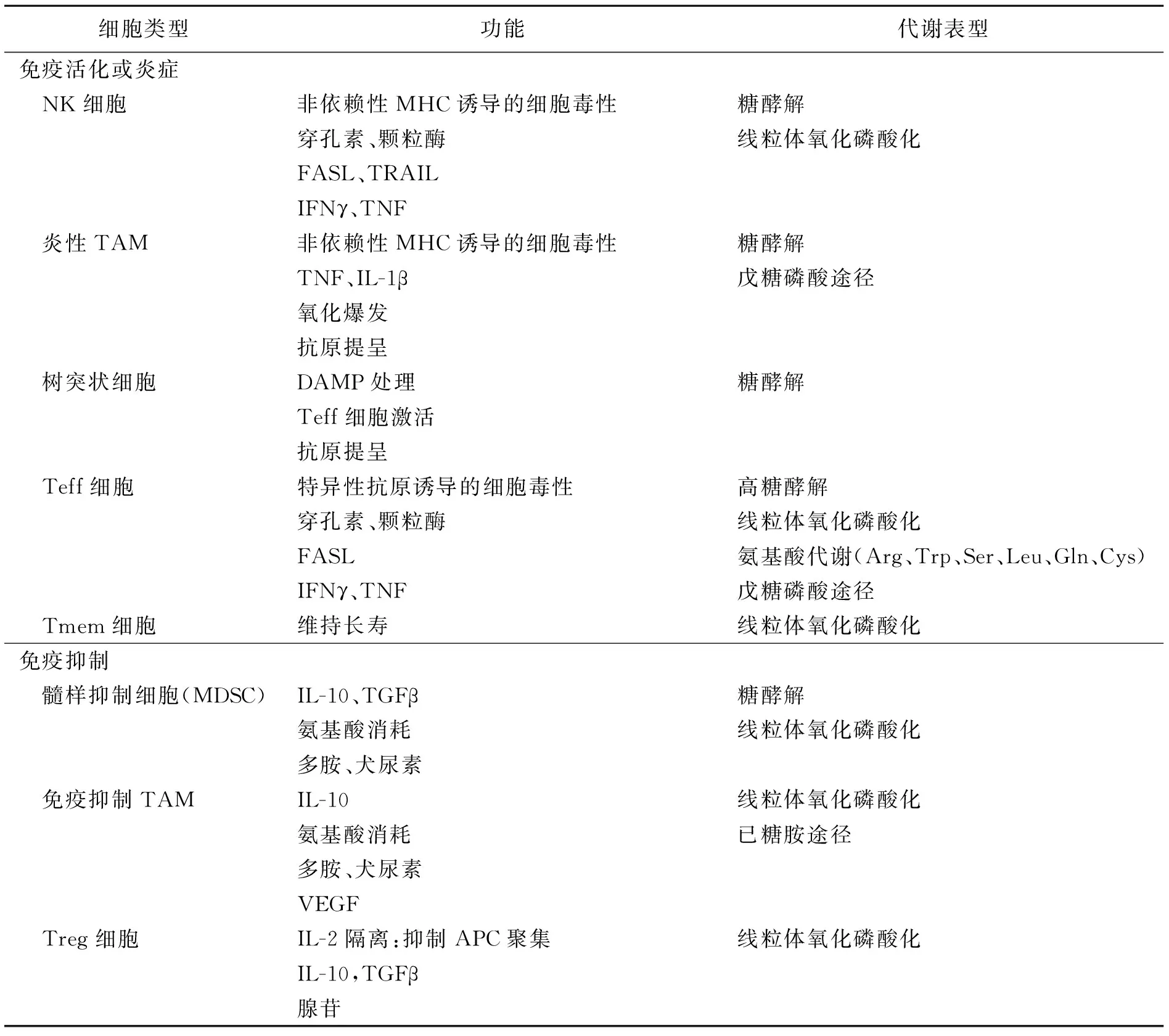
表1 TME中的免疫细胞类型、功能及代谢
TME中肿瘤和基质细胞乳酸分泌的增加,加快了肿瘤细胞的增进,调整了调节性T细胞(regulatory T cell,Treg)免疫机制,进而促进了肿瘤细胞隐匿免疫抑制。免疫细胞在增长、分化、执行效应器功能中会发生代谢的重新编程,对免疫应答起决定性效用[6-7]。肿瘤细胞通过排放代谢的产物(乳酸、PGE2、精氨酸等)影响免疫分子,比如细胞外囊泡向免疫细胞远距离释放精氨酸1,进而影响精氨酸代谢,从而达到降低机体抗肿瘤免疫反应的作用[8]。肿瘤细胞、Treg细胞通过强化免疫细胞磷脂酶A2-IVA表达,指引免疫细胞脂质代谢的重新编程与老化,从而减少了对肿瘤细胞的阻抑,减低抗肿瘤免疫应对[9]。
2 线粒体对肿瘤细胞和免疫细胞代谢的影响
在肿瘤发生进展过程中,糖代谢、脂肪酸代谢及氨基酸代谢异常[10]。同时,线粒体代谢性产物也对免疫细胞的代谢产生影响(图2)[5]。肿瘤细胞中异常水平的ROS会破坏线粒体DNA(mitochondrial DNA,mtDNA),进而影响线粒体膜呼吸链与生产腺苷三磷酸酶系统的正常进行,ROS水平的急剧上升导致线粒体功能加速被破坏,从而导致肿瘤细胞通过糖酵解途径获取腺苷三磷酸(ATP),发生代谢重编程[11]。
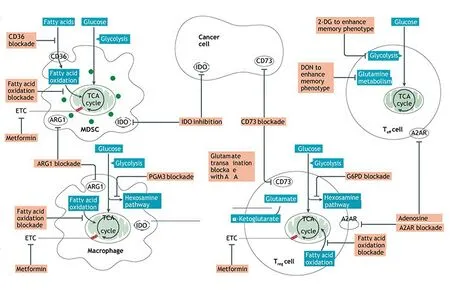
图2 TME中不同类型免疫细胞线粒体代谢特征
2.1 线粒体代谢改变
2.1.1糖代谢
肿瘤细胞特有的代谢产物为其蛋白质、核酸、脂类的合成与代谢提供原料,进而促进了肿瘤细胞存活,如糖酵解中间产物3-磷酸甘油和丙酮酸可用于合成非必需氨基酸[12]。另外,肿瘤细胞通过糖酵解会产生大量的乳酸,被释放到细胞外后,形成了有利于侵袭性高的肿瘤细胞生长繁殖的局部酸性微环境,还可通过抑制肿瘤免疫来达到促进肿瘤发展的目的[13]。有学者以小鼠为研究对象,发现抗肿瘤CD8+T细胞的效应表达与葡萄糖的可获得性成正比,而抗肿瘤CD4+T细胞的葡萄糖摄取和功能表达与肿瘤细胞的糖酵解活性成反比[14]。
2.1.2脂肪酸代谢
早在胚胎发育时脂肪酸的从头合成就很活跃,但大部分正常细胞会选择优先利用外源的脂肪酸合成功能性脂类。与之不同的是,外源脂肪酸的水平无论高低都不会影响肿瘤细胞进行大量内源性脂肪酸的合成。柠檬酸裂解酶促进线粒体中柠檬酸转化成细胞基质中的乙酰辅酶A,从而促进脂肪生成,来维持肿瘤的生长。肿瘤细胞中脂肪酸从头合成增强通过诱导内源性ATP柠檬酸裂合酶活性,促进了B细胞增殖、分化和抗体的分泌[15]。
2.1.3氨基酸代谢
谷氨酰胺是血浆中最多的氨基酸,其代谢是否活跃直接决定了肿瘤细胞氨基酸代谢。在线粒体功能障碍或缺氧甚至无氧的条件下,肿瘤细胞对谷氨酰胺的吸收与代谢异常活跃,大概是其他氨基酸的10倍,生成α-酮戊二酸,为嘌呤、嘧啶和DNA的合成提供氮,为脂质合成和肿瘤细胞快速增殖提供原料[16]。此外谷氨酰胺也被免疫细胞充分利用,特别是支持淋巴细胞和巨噬细胞的免疫反应。谷氨酰胺的缺乏可使T细胞的增殖和细胞因子的产生受到抑制[17],这也暗示了肿瘤微环境中高氨基酸含量可增强免疫细胞的杀伤作用。
2.2 线粒体结构异常
2.2.1mtDNA的突变
线粒体基因组是一个小的环状DNA分子,在每个线粒体的基质中mtDNA复制与细胞周期无关,可以多次进行。由于没有核膜包被暴露在基质中,缺乏保护的同时线粒体中缺乏DNA修复系统,所以mtDNA与核基因相比来说发生突变的概率更大,受损后序列发生改变,造成编码蛋白构象改变,线粒体功能发生障碍。对肿瘤mtDNA的检测中发现,在结肠癌、乳腺癌、肝癌、肺癌等多种肿瘤中均有mtDNA突变发生,其中mtDNA拷贝数的变化与肿瘤类型有关,小细胞肺癌mtDNA突变引起了能量代谢改变[18]。
2.2.2线粒体膜稳定性
线粒体膜稳定性对细胞凋亡以及肿瘤的发生发展有着较大影响。线粒体外膜上有Bcl-2蛋白和电压依赖性阴离子通道(voltage-dependent anion channel,VDAC)蛋白,前者是从滤泡B细胞瘤中获得的第一个抗凋亡蛋白,后者主要负责线粒体和细胞质之间代谢物质流通。研究发现Bcl-2家族蛋白可通过维持VDAC的开放状态来影响线粒体外膜的稳定性,从而抑制凋亡的发生[19]。线粒体的形态控制着T细胞的代谢,并可通过增强T细胞识别从而破坏肿瘤细胞。激活的效应T细胞拥有分裂的线粒体,而记忆T细胞维持它们的线粒体为融合网状物[20]。这证实了线粒体结构对调节免疫细胞新陈代谢至关重要。
2.2.3线粒体ROS紊乱
ROS以超氧化物和羟基自由基以及过氧化氢的形式,在生理状态下由物质代谢反应产生。各个氧化代谢中的ROS可通过氧化丙酮酸激酶M2来驱动戊糖途径的通量,增加谷胱甘肽的水平,促进肿瘤细胞发生发展。细胞ROS产生的主要位点是线粒体复合物Ⅰ和Ⅲ,与此同时,线粒体也是其损伤位点之一。例如,它们攻击线粒体膜和靠近它们形成地方的mtDNA,这导致线粒体功能障碍和更多的ROS产生。Maillet等发现结肠癌动物模型中,能够通过ROS依赖途径,引起线粒体形态和功能的一系列改变,从而触发肿瘤细胞凋亡,抑制肿瘤生长[21]。
2.2.4线粒体钙离子紊乱
线粒体中含有大量的Ca2+,并且控制细胞内钙离子的吸收和摄取。线粒体Ca2+信号与细胞生长和代谢密切相关,通过激活TCA循环的多种成分(IDH、aKGDH和PDH)构成存活前机制,从而为电子传递链和ATP产生提供能量[22]。减少线粒体Ca2+转移可以使瘤细胞避免凋亡,但线粒体Ca2+摄取的减少仍然能够激活TCA循环,使供给电子传递链的减少当量的产生变得平滑,并维持足够的ATP以满足细胞需求[23]。乳腺癌、结肠癌、胃癌细胞中Ins P3R表达显著增多,导致线粒体Ca2+超载,肿瘤细胞代谢紊乱,抑制了细胞凋亡的发生,促进了肿瘤的增殖和存活[24]。但线粒体钙稳态失衡影响肿瘤发生发展的具体作用机制目前仍不清楚。
3 靶向肿瘤线粒体是干预热点
线粒体是细胞的“动力工厂”,作为很多药物或毒物靶细胞器,可以有效提高药物的治疗效果。科学家进一步探究以线粒体为靶点的药物与抗肿瘤之间的作用机制。目前已有一些靶向线粒体代谢的抗肿瘤药物在临床或临床前开发中,如WZB117和紫杉醇或顺铂联合治疗可以发挥协同抗肿瘤作用。这类药物的作用机理是阻断葡萄糖转运,通过阻断葡萄糖转运蛋白1活性,减少细胞内葡萄糖的转运效率,降低肿瘤内糖酵解的代谢[25],进而逆转肿瘤细胞的高糖酵解状态,阻止肿瘤细胞摄取葡萄糖。某些肿瘤细胞钠-葡萄糖转运体2(sodium glucose transporter 2,SGLT2)水平明显增高,采用FDA批准的小分子抑制剂靶向SGLT2降低血糖水平,可以降低肺腺癌小鼠模型肿瘤的生长,延长生存时间[26]。另一种药物是P-糖蛋白,它是一种具有ATP结合域的单向转运跨膜蛋白[27],这个药物主要通过水热合成法、薄膜分散法构建了线粒体靶向,应用ATP水解释放能量将药物主动排出,进而避免了细胞内药物浓度过高,解决了肿瘤细胞的耐药性。这一系列研究表明靶向肿瘤线粒体可以有效地提高药物对肿瘤细胞的毒性以及改善药物的活性,同时靶向肿瘤线粒体可以有效地克服药物的耐药性。
4 总结与展望
肿瘤细胞可通过物质代谢抑制机体抗肿瘤免疫反应;而小分子抑制剂可促进T细胞增殖,减轻髓样细胞介导的免疫逃逸,抑制肿瘤生长。在探究肿瘤细胞和免疫细胞的相互作用时,线粒体靶向治疗的研究与发现无疑是这场肿瘤生态系统中代谢竞争的“关键一物”。研究线粒体对肿瘤细胞代谢以及免疫细胞的影响机制,对线粒体与二者之间的相互关系做出探讨。研究发现肿瘤及免疫细胞代谢与线粒体代谢、结构的改变密切相关,当线粒体处于不同糖代谢、脂肪酸代谢、氨基酸代谢的变化时,二者的代谢过程也随之发生不同的改变。同时,随着线粒体中Ca2+稳态的改变,ROS水平与肿瘤细胞增殖代谢高度联系。因此,在个体化治疗基础上,线粒体能量代谢特征与肿瘤细胞代谢特征不容小觑,靶向肿瘤线粒体作为肿瘤细胞的“十环之靶心”更为肿瘤细胞免疫治疗机制提供新的策略方法。
