PLZF调控小鼠早期T细胞发育和自我更新的功能①
2023-03-17李凤迪袁肇方刘新宇黄海岩杜江龙蔡葵蒸西北民族大学生命科学与工程学院兰州730030
李凤迪 袁肇方 刘新宇 黄海岩 杜江龙 蔡葵蒸 (西北民族大学生命科学与工程学院,兰州 730030)
早幼粒细胞白血病锌指基因也被称为Zbtb16基因,其调控的蛋白称为PLZF(the protein encoded by pro-myelocytic leukemia zinc finger,PLZF)。Zbtb16首次发现是在一位急性早幼粒细胞白血病患者中,这是一个以前未报道过的锌指基因,与其t(11;17)染色体易位有关,目前研究最多的是其炎症反应抑制因子的功能[1-2]。近年来,PLZF 已成为一种重要的免疫调节因子并在细胞生长中起关键作用,既能抑制反应性T 细胞的增殖和成熟也能诱导其凋亡[3-4]。PLZF 能在先天淋巴样细胞(innate lymphoid cells,ILCs)、CD8+T 细胞以及CD4+T 细胞中表达,尤其是在iNKT 细胞(invariant natural killer T cell,iNKT)中高表达,是一种标志性的转录调节因子,对iNKT 细胞分化、发育和功能的调控是必不可少的[5-7]。PLZF作为一种多功能的蛋白质,可调节多种生物过程:如精子发生、细胞增殖、分化、器官发育、干细胞维持和先天性免疫细胞发育以及肿瘤形成等[8]。目前,许多研究表明PLZF 参与癌症的发展,能够抑制许多癌细胞生长,包括黑色素瘤、恶性间皮瘤、前列腺癌和非小细胞肺癌等[5,9-11]。PLZF 之所以作为一个标志外周T 细胞淋巴瘤(PTCL)亚群(成熟T 细胞淋巴瘤的异质性疾病类型)的蛋白质,是因为其存在人类淋巴细胞中多于在小鼠中[12-13]。早期T 细胞系前体在胸腺中发育为T淋巴细胞是一个高度调控的过程,调节涉及外部信号和细胞内在机制[14]。最近研究发现PLZF 能调节造血干细胞功能,参与细胞周期调控,并在控制造血干细胞稳态中起关键作用[15]。本文通过PLZF-EGFP 报告基因小鼠和新生小鼠胸腺的肾移植模型,利用流式细胞术和细胞分选以及RT-PCR 技术,初步研究PLZF 在小鼠早期T细胞发育和自我更新过程中的功能,对PLZF 是否参与胸腺内T 细胞的早期发育及维持早期T 细胞系前体的自我更新进行系统性研究。
1 材料与方法
1.1 材料
1.1.1 实验动物 动物实验由西北民族大学实验动物伦理委员会审批。PLZF-/-、PLZF-EGFP 报告基因小鼠(简称为PEG 小鼠),其EGFP 的表达受PLZF启动子活性的调控,Rag2-/-和Rag2/γc-/-小鼠购自上海南方模式生物科技有限公司。所有小鼠均为C57BL/6 背景,在4~8 周龄时用于实验。实验中PLZF-/-小鼠的对照为条件许可时采用同窝出生的野生型小鼠,或年龄和性别一致的C57BL/6小鼠。
1.1.2 主要试剂及仪器 RNA 提取试剂(QIAGEN公司);实时荧光定量PCR 试剂(Bio-Rad 公司);胎牛血清(Gibco 公司);BSA(Amresco 公司);DMEM、胰蛋白酶(兰州民海生物公司);Lipofectamine 2000(Thermo Fisher Scientific 公司);阿维汀、甲醇、冰醋酸、无水乙醇、NaHCO3、异丙酮(上海生工公司); 细胞培养皿(corning 公司);流式细胞术用到的抗 体如下:CD45.1(A20)、CD45.2(104)、Sca-1(E13-161.7)、c-Kit(2B8)、Flt3(A2F10)、IL-7Ra(A7R34)、CD93(AA4.1)、B220(RA3-6B2)、CD19(6D5)、CD11b(M1/70)、Gr-1(RB6-8C5)、CD44(IM7)、CD25(PC61)、Thy1.2(30-H12)、CD4(RM4-5)、CD8α(53-6.7)、TCR-β(H57),均购自Biolegend 公司;流式细胞仪(LSR Ⅱ and FACSAria cell sorter,BD公司)。
1.2 方法
1.2.1 新生小鼠胸腺移植模型的建立 将新生小鼠(CD45.2)的胸腺取下,移植于同系受体小鼠(CD45.1)的肾被膜下,用阿维汀(2.5%溶液,10 U/g体质量)腹腔注射麻醉。在腹腔做一个5 mm的切口,体式显微镜下暴露左肾,用显微解剖钳将单个新生鼠的两个胸腺叶置于肾被膜下,随后缝合伤口。
1.2.2 流式细胞术和细胞分选 胸腺移植的小鼠饲养3 周后,解剖小鼠并制备胸腺和骨髓单细胞悬液,使细胞悬浮于含0.1%BSA和0.08%叠氮化钠的PBS 缓冲液中(染色缓冲液)。于4 ℃下将抗体在染色缓冲液中孵育20 min。封闭Fc 受体:使用流式Staining Buffer 洗涤板孔2 次,200 µl/孔,在4 ℃、 1 500 r/min 下离心5 min;然后加入Staining Buffer 稀释受体阻断剂,1 µg/100 µl 中细胞106个,4 ℃放置 15 min 封闭阻断受体。在Fc 受体封闭期间,配制表面染色抗体混合体系,封闭完成后使用Staining Buffer 洗涤板孔2 次,200 µl/孔,4 ℃、1 500 r/min 离心5 min,弃液体,振荡重悬,加入表面染色抗体混合物 50 µl/孔,轻敲混匀,4 ℃条件下避光孵育30 min。将带有生物素标记的抗体CD3ε、B220、TER-119、CD11bGr-1 混合在一起制成抗体混合物,命名为Lineage。将新鲜分离的胸腺细胞与Lineage 共同孵育来分选造血干细胞LSK。为了保留阴性细胞,用磁珠分离(MACS)将Lineage 阳性(Lin+)细胞去除。细胞富集后,用表面标志物对Lineage阴性细胞进行染色。然后上机分析,并采用FloJo进行数据分析。
1.2.3 RNA 提取和实时定量PCR RNA 提取采用RNA 试剂盒操作,反转录采用iScriptTMcDNA Synthesis Kit 试剂盒操作。其中反转录反应条件设定:25 ℃ 5 min,反转录46 ℃ 20 min,RT失活95℃ 1 min,4 ℃保存。待检基因的蛋白表达水平用HPRT (hypoxanthine phosphoribosyltransferase)mRNA 比对进行计算[16]。RT-PCR 所使用的引物如下:HPRT 正向引物5'-CTGCGGAAAACGGTTCCTG-3',反向引物5'-GTGCCAGTATGGGTCTGTCT-3'。
1.3 统计学处理 本研究中的数据使用GraphPad Prism 5(San Diego,CA,USA)进行数据统计学分析。组间比较采用独立样本t检验,P<0.05 为差异有统计学意义。
2 结果
2.1 PLZF-/-小鼠的胸腺细胞总数与野生型小鼠相比减少 分别取12 只PLZF+/+和12 只PLZF-/-小鼠的胸腺,对其胸腺细胞总数进行计数。结果显示,PLZF-/-小鼠的胸腺细胞总数明显少于野生型小鼠的胸腺细胞总数(图1)。由GraphPad Prism 5.0 软件进行数据统计学分析,其P=0.002<0.01,差异具有统计学意义。
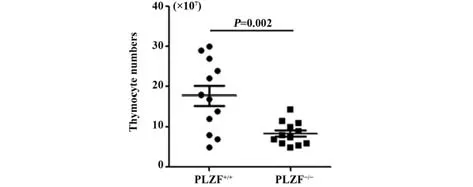
图1 PLZF+/+和PLZF-/-小鼠的胸腺细胞总数比较Fig.1 Comparison of total number of thymocytes in PLZF+/+ and PLZF-/- mice
2.2 小鼠骨髓造血干细胞(hematopoietic stem cell,HSC)和胸腺DN2a 细胞中PLZF 呈高表达 为进一步研究PLZF 在T细胞发育过程中的表达情况,利用PEG 转基因小鼠模型。分别获取C57BL/6和PEG 小鼠的骨髓细胞,利用流式细胞仪对LSK 细胞(Lin-Sca-1+c-Kit+)进行开窗分析,根据其Flt3 的表达情况,将LSK 细胞分为HSC(LSK Flt3lo)和淋巴预处理多能祖细胞(lymphoid-primed multipotent progenitor,LMPP)(LSK Flt3hi)两群。发现PEG 小鼠的HSC细胞的GFP 表达水平较高,而LMPP 细胞的GFP 水平与C57BL/6 小鼠表达无明显差异(图2A)。说明PLZF 在骨髓的HSC 中表达,而随后在LMPP 中表达沉默。紧接着又分别获取C57BL/6和PEG小鼠的胸腺细胞,利用流式细胞术对Lin-CD44+c-Kit+细胞进行开窗分析,根据其CD25 的表达情况,将Lin-CD44+c-Kit+细胞分早期T 细胞系前体(early T cell pro-genitors,ETP)(Lin-CD44+c-Kit+CD25-)和(CD4 CD8 double negative,DN)DN2a(Lin-CD44+c-Kit+CD25+)两群。发现PEG 小鼠ETP 细胞的GFP 水平与C57BL/6小鼠相当,而PEG 小鼠DN2a 细胞的GFP 水平显著升高(图2B)。综合PEG 小鼠骨髓和胸腺细胞的实验结果,表明PLZF 在HSC 中表达,当其分化为LMPP 时表达下降乃至沉默,一直到迁移至胸腺发育为DN2a细胞时又重新表达(图2C)。通过流式细胞术分选LSK、ETP、DN2(CD4-CD8-CD44+CD25+)和DN3(CD4-CD8-CD44-CD25+)细胞,利用RT-PCR 检测PLZF 的转录水平,发现PLZF 在造血干细胞LSK和DN2细胞中有较高的转录水平。但当T细胞发育至DN3时期时,PLZF的转录又降至极低水平。上述结果表明,PLZF 在T 细胞发育过程中是一个动态调控的过程(图2D)。
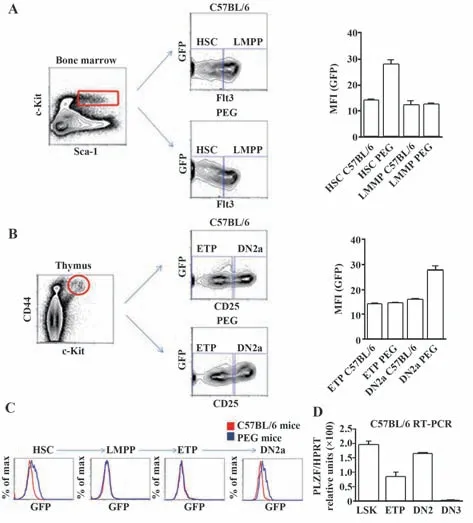
图2 T细胞发育过程中PLZF的表达情况Fig.2 Expression of PLZF during development of T cells
2.3 PLZF-/-小鼠的ETP数目减少 为检测PLZF对T 细胞早期发育的影响,分别获取C57BL/6 和PLZF-/-小鼠的胸腺细胞,利用流式细胞术对CD4-CD8-TCRβ-CD44+c-Kit+细胞进行开窗分析。结果显示,PLZF-/-似乎对胸腺细胞中ETP和DN2a细胞的比例影响不大(图3A)。但ETP 细胞的绝对数目在PLZF-/-小鼠中较C57BL/6 小鼠显著减少,仅有50%左右(图3B)。
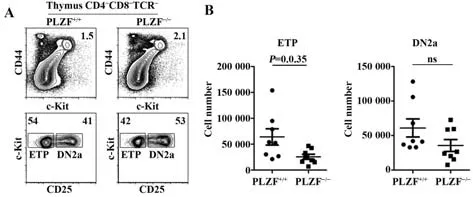
图3 PLZF-/-对小鼠ETP细胞的影响Fig.3 Effect of PLZF-/- on ETP cells in mice
2.4 PLZF-/-小鼠的胸腺细胞减少并非细胞内在性的 为进一步判断PLZF-/-小鼠的胸腺细胞总数减少是否是造血干细胞内在性导致的,采用竞争性骨髓嵌合小鼠模型。分别将PLZF+/+(Ly5.1)和PLZF-/-小鼠经免疫磁珠分选后富集的Sca-1+骨髓细胞按1∶1等量混合作为供体细胞,经尾静脉注射给预先经亚致死剂量辐射过的受体小鼠(Rag2/γc-/-),饲养8 周后取胸腺进行流式细胞术分析。Rag2 和γc 双基因缺陷使得受体小鼠自身的T、B 和NK 细胞均无法正常发育,只能完全来源于外源的供体细胞。结果表明,经骨髓移植的受体小鼠胸腺细胞中,PLZF+/+和PLZF-/-来源的细胞比例大致相等,且均能在胸腺中进行正常的T 细胞发育(图4A)。此外,在移植后的8周、24周取受体小鼠的脾脏进行分析,发现脾脏中B 细胞和T 细胞的PLZF+/+(Ly5.1)来源的细胞比例基本保持恒定,维持在50%左右,PLZF-/-细胞在竞争中保持水平(图4B)。以上结果证明PLZF-/-小鼠的胸腺细胞减少不是细胞内在性的原因。
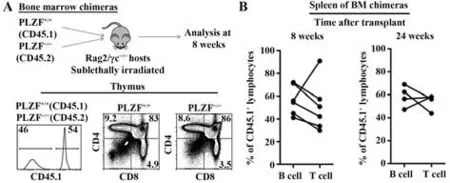
图4 PLZF-/-小鼠胸腺细胞减少的原因Fig.4 Causes of thymocytopenia in PLZF-/- mice
2.5 PLZF 在新生小鼠胸腺移植模型的移植物DN1细胞中高表达 为进一步研究PLZF 在T 细胞发育过程中的DN1 表达情况,将新生C57BL/6 或PEG 小鼠的胸腺取下,移植于受体小鼠Rag2/γc-/-的肾被膜下,4周后取胸腺移植物进行流式细胞术分析(图5A)。对CD4-CD8-TCRβ-(简称为TN)的胸腺细胞进行开窗分析,将DN1 细胞(CD44+CD25-TN)按γc 的表达水平分为供体细胞(γc+)和受体细胞(γc-)。对来源于PEG 小鼠的供体细胞进行GFP 荧光强度分析发现,PLZF 在胸腺移植物的DN1 细胞中高表达,甚至高于阳性对照iNKT细胞(图5B)。
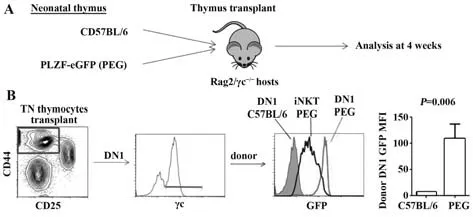
图5 新生小鼠胸腺移植模型的移植物DN1 细胞中PLZF高表达Fig.5 PLZF is high expression in DN1 cells of thymus transplantation model in neonatal mice
2.6 PLZF-/-可能影响新生小鼠胸腺移植模型的供体DN1 细胞自我更新 将新生PLZF+/+或PLZF-/-小鼠的胸腺取下,移植于受体小鼠Rag2/γc-/-的肾被膜下,4周后取胸腺移植物进行流式细胞术分析(图6A)。对TN 胸腺细胞进行开窗分析,将DN1 细胞(CD44+CD25-TN)按γc 的表达水平分为供体细胞(γc+)和受体细胞(γc-)。结果显示,在胸腺移植物中DN1 细胞来源于供体,与PLZF-/-小鼠胸腺作为供体的比例远低于PLZF+/+小鼠胸腺作为供体(图6B)。随后将PLZF-/-小鼠胸腺作为供体与PLZF+/+小鼠胸腺进行比较,胸腺移植物中DN1 细胞来源于供体的绝对数量减少,而来源于受体小鼠的绝对数量增多(图6C)。PLZF-/-并不影响胸腺移植物中供体T 细胞的正常发育,且胸腺移植物的绝对细胞数量与野生型差异无统计学意义(图6D)。上述结果表明,PLZF-/-可能对T 细胞的正常发育和分化没有影响,但很可能参与维持早期T细胞系前体的自我更新。
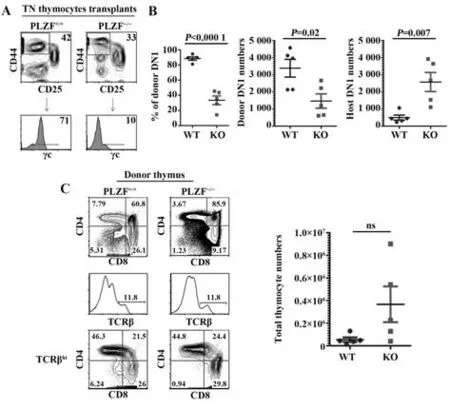
图6 PLZF-/-可能影响新生小鼠胸腺移植模型的供体DN1细胞自我更新Fig.6 PLZF-/- may affect donor DN1 cell self-renewal in neonatal thymus transplantation model
3 讨论
本研究通过竞争性骨髓嵌合小鼠模型,发现经骨髓移植的受体小鼠胸腺细胞中,PLZF野生型和突变型来源的细胞比例大致相等,且均能在胸腺中进行正常的T 细胞发育,表明PLZF 对T 细胞分化发育的影响不是造血干细胞内在性的。但在新生小鼠胸腺移植模型中,Rag2/γc-/-作为受体小鼠,PLZF-/-会导致胸腺移植物中DN1 细胞来自供体的比例显著减少。当受体来源的早期T细胞系前体能够分化至超越DN2 阶段,供体胸腺细胞最终从胸腺移植物中消失。但当没有持续的早期T 细胞系前体供应(如受体小鼠同时缺乏SCF 和IL-7 信号),或早期T 细胞系前体发育阻滞于DN3 阶段之前(如受体小鼠 IL-7信号缺陷),供体胸腺细胞转变为在胸腺移植物中持续自主产生T细胞的模式。由于此受体小鼠缺乏IL-7 信号,“DN3 生态位”缺乏宿主来源的祖细胞的空间竞争,故供体胸腺细胞具备了自我更新的特征,而PLZF参与供体胸腺细胞的自我更新。
由于胸腺细胞自身寿命短,且缺乏具有自我更新能力的ETP 或长期驻留的胸腺细胞,所以胸腺的功能完全依赖于从骨髓来源的早期T 细胞系前体[17]。PEAUDECERF 等[18]研究发现胸腺细胞能被骨髓迁移而来的早期T 细胞系前体迅速取代,即使骨髓前体细胞不能产生T 细胞,但也会发生这种取代,如Rag1/2-/-或严重联合免疫缺陷(SCID)的小鼠。相反的是,MARTINS 等[19]研究了将新生小鼠胸腺移植于缺失IL-7 的小鼠中,发现新生小鼠胸腺具有自我更新的细胞群,且独立于骨髓的任何能维持胸腺细胞群体,并能迅速重建外周T 细胞群,以及清除T细胞缺陷小鼠的受感染能力。因此,这就能解释既往的试验中虽能观察到供体胸腺细胞消失但不能归结为胸腺细胞寿命短的原因,当新出现的受体来源DN3 胸腺细胞和已产生的供体来源的胸腺细胞间存在竞争时,供体来源的胸腺细胞消失[20]。
本研究有助于全面揭示PLZF 调控早期T 细胞发育和自我更新的功能,为PLZF 在T细胞分化发育的功能提供了科学依据。
