弓形虫SRS29C基因敲除株的构建与鉴定
2023-03-08姜阜杉叶建军高传亮王宇冰艾晴晴宋淇淇
姜阜杉,叶建军,田 静,高传亮,王宇冰,艾晴晴,魏 瑶,宋淇淇
(天津农学院 动物科学与动物医学学院 天津市农业动物繁育与健康养殖重点实验室,天津 300392)
刚地弓形虫(Toxoplasmagondii)是一种专性细胞内寄生的机会性致病原虫,可感染包括人类在内的所有温血动物,呈世界性分布,是一种重要的人畜共患寄生虫[1]。孕妇感染弓形虫后除了造成流产和死胎之外,还可导致发育中的胎儿患上先天性弓形虫病,表现为心智发育不良、小头畸形、脑积水、视网膜脉络膜炎等症状[2]。家畜感染弓形虫后主要表现为流产、死胎、木乃伊胎等,同时导致肉制品和奶制品质量下降[3]。弓形虫给人类健康和养殖业的发展构成了严重的威胁。
作为一种细胞内寄生原虫,弓形虫具有大量表面抗原,这些抗原有作为诊断和疫苗候选分子的双重价值。目前,已知多种表面抗原均属于SRS蛋白超家族,多以糖基磷脂酰化形式锚定在细胞膜上[4]。弓形虫基因组的分析预测有161种独特的SRS蛋白[5],在弓形虫速殖子(急性感染)中,其表面的主要成分为SAG1、SAG2A、SAG3、SRS1、SRS2和SRS3,这些成为宿主免疫的主要目标[6]。而在缓殖子(慢性感染)中,SRS9是最丰富的SRS抗原之一,其对于弓形虫脑包囊的形成有重要作用[7]。SRS抗原一方面在入侵宿主细胞、免疫调节等方面发挥作用,另一方面,也为弓形虫在宿主环境中生存提供所需要的保护。
同为肉孢子虫科的新孢子虫,其与弓形虫在形态结构、生活史、基因和抗原方面都极为相似。新孢子虫NcSRS2、NcSAG1基因与刚地弓形虫SRS2(现重命名为SRS29C)、SAG1基因高度同源[8]。NISHIKWA等[9]分别用携带有NcSRS2和NcSAG1基因的重组痘苗病毒免疫BALB/c小鼠后,发现此疫苗可有效阻止小鼠中新孢子虫的侵袭。刘梦丽等[10]利用重组表达的NcSRS2和NcSAG1蛋白与弗氏佐剂混匀后免疫BALB/c小鼠,结果表明NcSRS2和NcSAG1重组蛋白可激发机体产生Th1型和Th2型免疫反应。鉴于NcSRS2在新孢子虫黏附及入侵宿主细胞过程中发挥的重要作用,以及其良好的免疫原性,推测与其具有高度同源性的弓形虫SRS2(现重命名为SRS29C)蛋白可能会在弓形虫中发挥相似的作用。因此,对弓形虫SRS29C蛋白进行系统的研究,有助于我们为弓形虫疫苗的研发提供新的靶基因。
1 材料与方法
1.1 虫体与细胞HFF细胞、弓形虫RH株、弓形虫ME49株由华中农业大学申邦教授惠赠。
1.2 主要试剂基因组提取试剂盒、质粒小提试剂盒购自天根生化科技(北京)有限公司;Q5定点突变试剂盒购自NEB公司;ClonExpress Ⅱ OneStep Cloning Kit购自南京诺唯赞生物科技股份有限公司;Transetta (DE3) Chemically Competent Cell、限制性内切酶购自北京全式金公司;乙胺嘧啶购自Sigma公司;二抗均购自碧云天公司;tgGT1-258700质粒、PUC19-(277760)质粒载体由华中农业大学申邦教授惠赠;pGEX-4T-1质粒由本实验室保存;Cytomix(120 mmol/L氯化钾、0.15 mmol/L氯化钙、5 mmol/L磷酸氢二钾、5 mmol/L磷酸二氢钾、2 mmol/L EDTA、5 mmol/L 6水合氯化镁均购自中国医药集团有限公司,25 mmol/L HEPES购自Sigma公司)。
1.3 SRS29C多克隆抗体的制备在NCBI网站已经公布的刚地弓形虫ME49虫株全基因组信息中下载SRS29C相关基因信息,其全长2 752 bp,其中第484~1 602位共1 119 bp为此基因的CDS序列,根据此CDS序列设计扩增全长基因序列的引物,上、下游引物分别带有Sal Ⅰ、Not Ⅰ酶切位点(表1)。PCR反应条件:95℃ 5 min;95℃ 30 s,55℃ 30 s,72℃ 90 s延伸,共30个循环;72℃ 5 min,15℃ 5 min。构建成功后转化进全式金Transetta (DE3) Chemically Competent Cell中大量表达,利用包涵体洗涤法纯化蛋白,浓缩后用新西兰大白兔进行免疫,获得的多克隆抗体用Western blot检验抗体免疫原性。
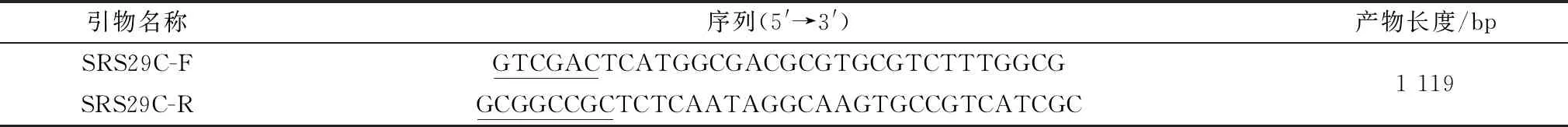
表1 SRS29C基因PCR扩增引物
1.4 SRS29C基因敲除重组质粒的构建利用E-CRISP在线软件对弓形虫SRS29C基因确定靶向gRNA序列,设计gRNA引物。用Q5定点突变试剂盒将tgGT1-258700质粒定点突变。利用Vazyme在线引物设计软件设计多片段同源重组引物,将SRS29C上游5′UTR、下游3′UTR及DHFR抗性基因表达盒连接到PUC19-(277760)质粒骨架上构建PUC19-(277760)5H-loxp-DHFR-loxp-3H同源模板质粒。
1.5 弓形虫的转染按照常规方法培养弓形虫速殖子约80%虫体逸出时,刮下虫体和细胞,5 mL注射器反复吹吸8~10次,破碎细胞。3 μm滤膜过滤纯化虫体,3 000 r/min离心5 min,收集虫体。用1 mL Cytomix洗涤虫体,3 000 r/min离心5 min,弃上清,最终用250 μL Cytomix重悬沉淀。在4 mm电转杯中加入同源片段1 500 ng和CRISPR质粒7 500 ng,加入虫体,终体积500 μL,混匀进行电转化。转化条件如下:1 600 V,25 μF,50 Ω,电转2~3次。电转后的虫体加入到新的HFF细胞中正常培养,同时培养未经电转的虫体作为对照组。培养12 h后加入终浓度为1 μmol/L的乙胺嘧啶继续培养。
1.6 基因敲除型虫株的筛选与鉴定
1.6.1基因敲除株虫库的鉴定 当培养瓶中有80%虫体逸出、细胞内纳虫泡较大时进行传代,传代过程中加入终浓度为1 μmol/L的乙胺嘧啶,直至空白对照的虫体全部死亡后,提取其DNA进行PCR鉴定。PCR引物见表2,PCR反应条件:95℃ 3 min;95℃ 30 s,55℃ 30 s,72℃ 1 min,30个循环;72℃ 5 min,15℃ 5 min。
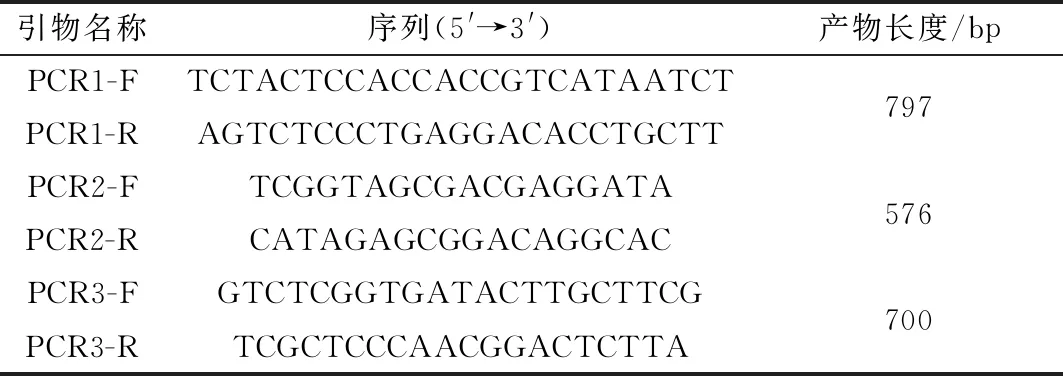
表2 SRS29C基因敲除株PCR鉴定引物
1.6.2基因敲除株的单克隆筛选 鉴定为阳性后,利用限制性稀释法在长满HFF细胞的96孔板中进行单克隆筛选,37℃、5% CO2条件下培养7 d。将筛选获得的单克隆接种到铺满HFF细胞的24孔板中扩大培养,之后对所有单克隆进行初步鉴定。鉴定方法同1.6.1。初步鉴定正确的单克隆敲除株接种到T25中扩大培养,进行Western blot鉴定。
1.7 基因敲除型虫株的Western blot鉴定收集等量的扩大培养的单克隆虫株及野生株,用裂解液将虫体充分裂解,此后用BCA法测定裂解产物的浓度。通过Western blot验证弓形虫SRS29C蛋白的表达情况,一抗为 Rabbit anti-SRS29C-GST IgG和Mouse anti-tublin IgG;二抗为辣根过氧化物酶(HRP)标记的山羊抗兔 IgG(H+L)和HRP标记的山羊抗小鼠 IgG(H+L)。
2 结果
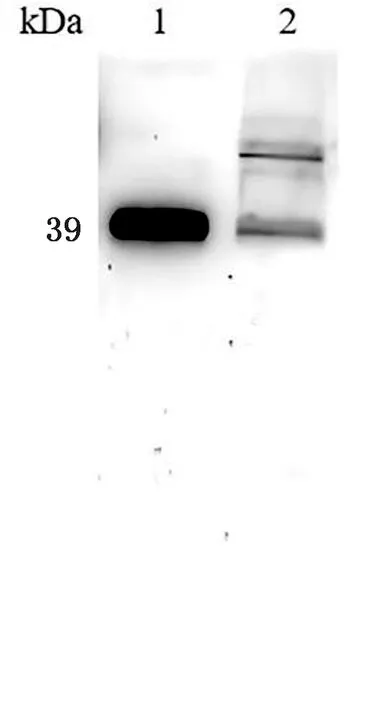
2.1 SRS29C多克隆抗体的制备重组原核表达质粒进行双酶切鉴定,结果显示,3,4,6号样品均获得4 957,1 126 bp的片段,为酶切正确样品(图1)。将测序正确的菌液标记为pGEX-4T-1-SRS29C。将正确的质粒进行原核表达,结果显示在65 kDa左右的位置有明显条带,表明重组蛋白可在Transetta (DE3) Chemically Competent Cell中表达,且以包涵体的形式表达(图2)。蛋白纯化结果表明,目的蛋白经过3次洗涤后基本将杂质清除(图3)。将纯化后的蛋白透析复性浓缩后进行免疫,获得的多克隆抗体利用Western blot进行抗体特异性检测,结果显示,均能在39 kDa左右显示出1条特异性条带,与SRS29C天然蛋白预测大小一致,表明制备的Rabbit anti-SRS29C-GST IgG免疫原性良好(图4)。
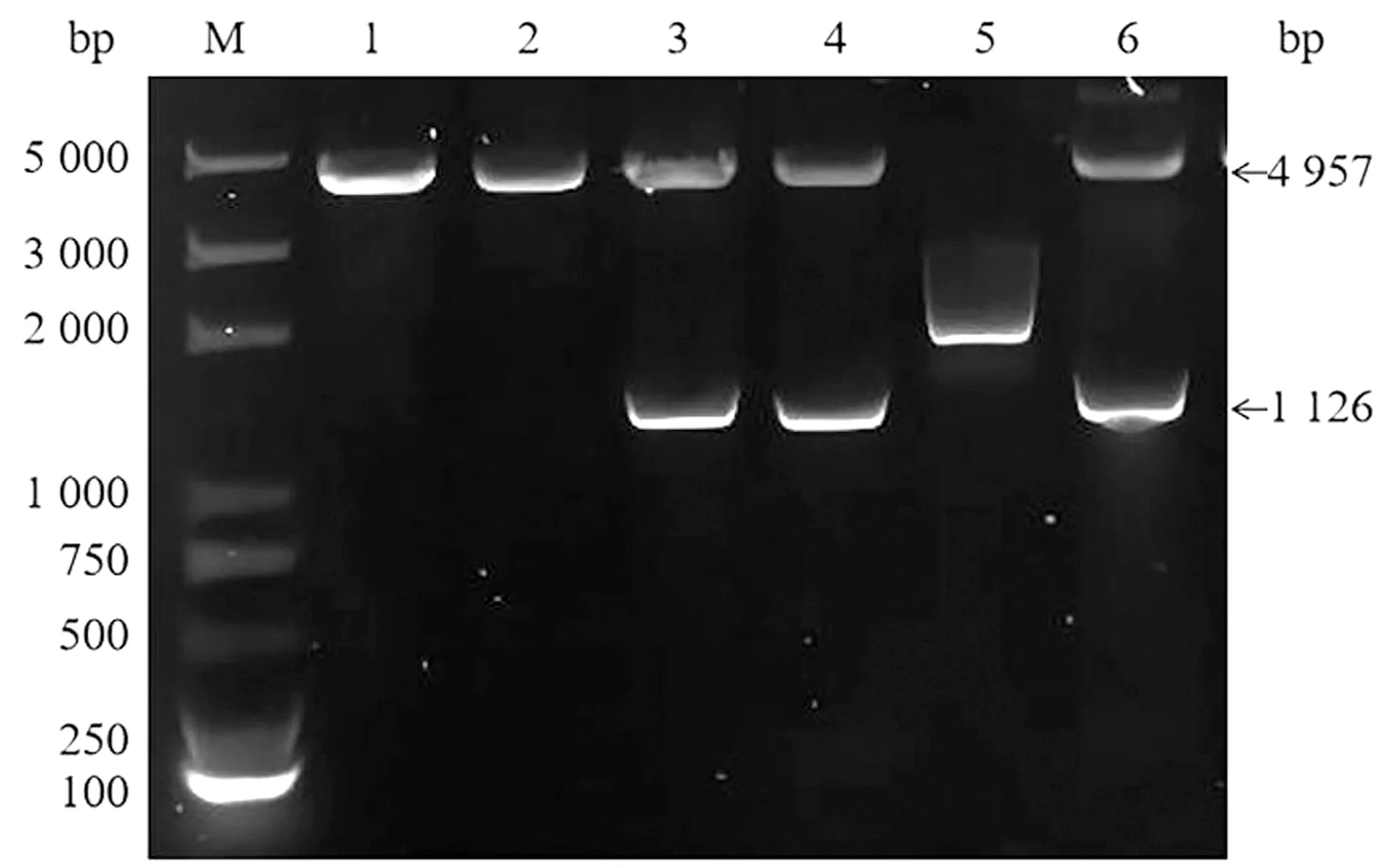
M.Trans2K© Plus DNA Marker;1~6.6个样品酶切产物
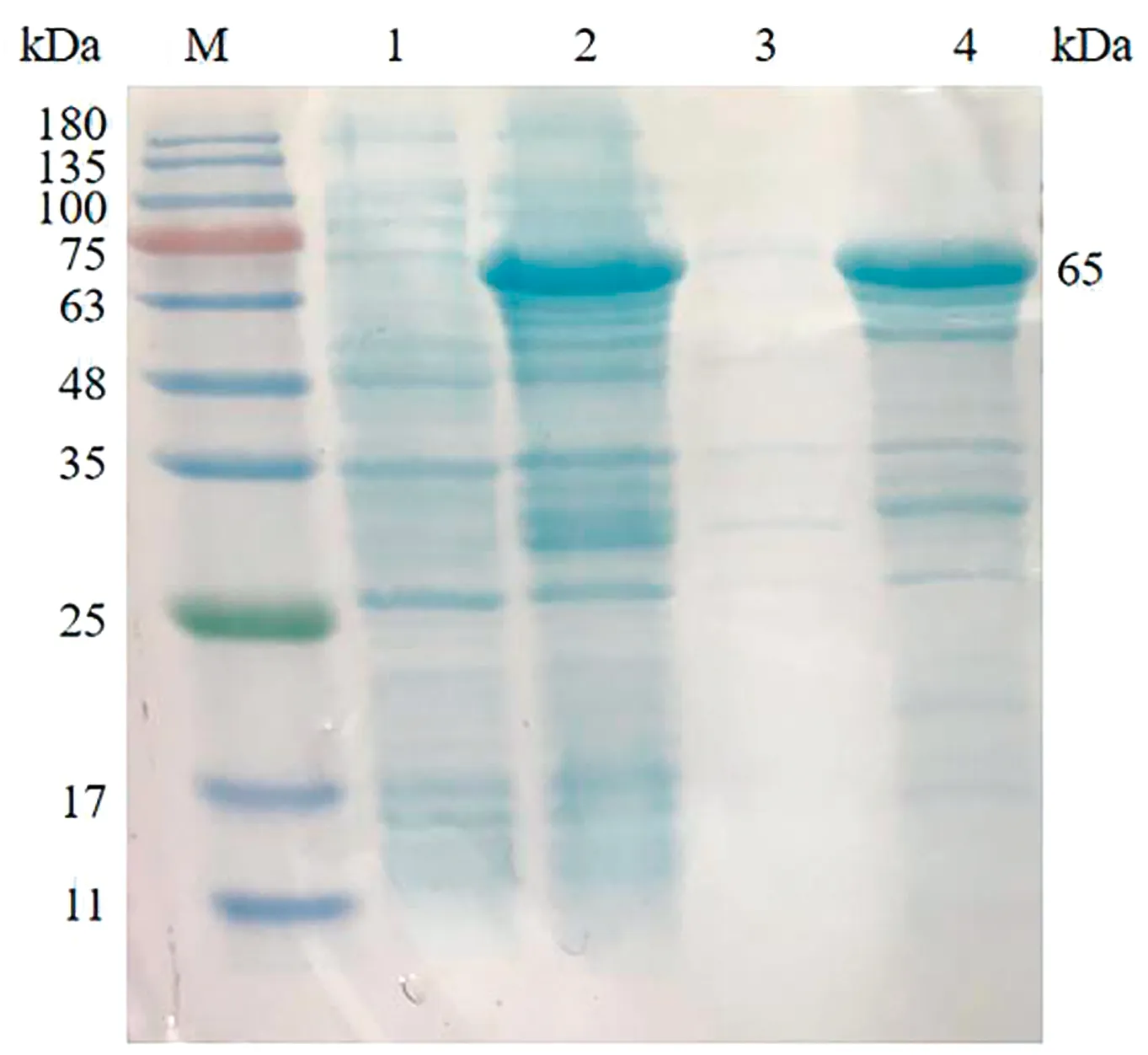
M.彩虹180广谱蛋白Marker;1.未诱导全菌;2.诱导后全菌;3.诱导破碎后上清液;4.诱导破碎后包涵体
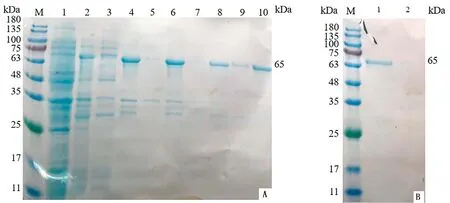
A.包涵体洗涤结果(M.彩虹180广谱蛋白Marker;1.未诱导全菌;2.诱导后全菌;3.诱导破碎后上清;4.诱导破碎后包涵体;5,7,9.包涵体第1,2,3次洗涤上清;6,8,10.包涵体第1,2,3次洗涤沉淀);B.包涵体溶解结果(M.彩虹180广谱蛋白Marker;1.第3次洗涤后溶解上清;2.第3次洗涤后溶解沉淀)
1.ME49全虫蛋白;2.RH全虫蛋白
2.2 SRS29C基因敲除质粒的构建将重组导向质粒利用Sal Ⅰ酶切鉴定(图5),结果显示在6 024,3 650 bp 处显示条带,与预期大小一致。同源模板质粒利用EcoRⅠ酶切鉴定,结果显示在4 548,2 038,1 194 bp处与321 bp处显示条带,与预期大小一致(图6)。将测序正确的质粒保存并用于后续试验。

M.DL15000 DNA Marker;1,2.pSAG1-Cas9-U6-gSRS29C酶切产物
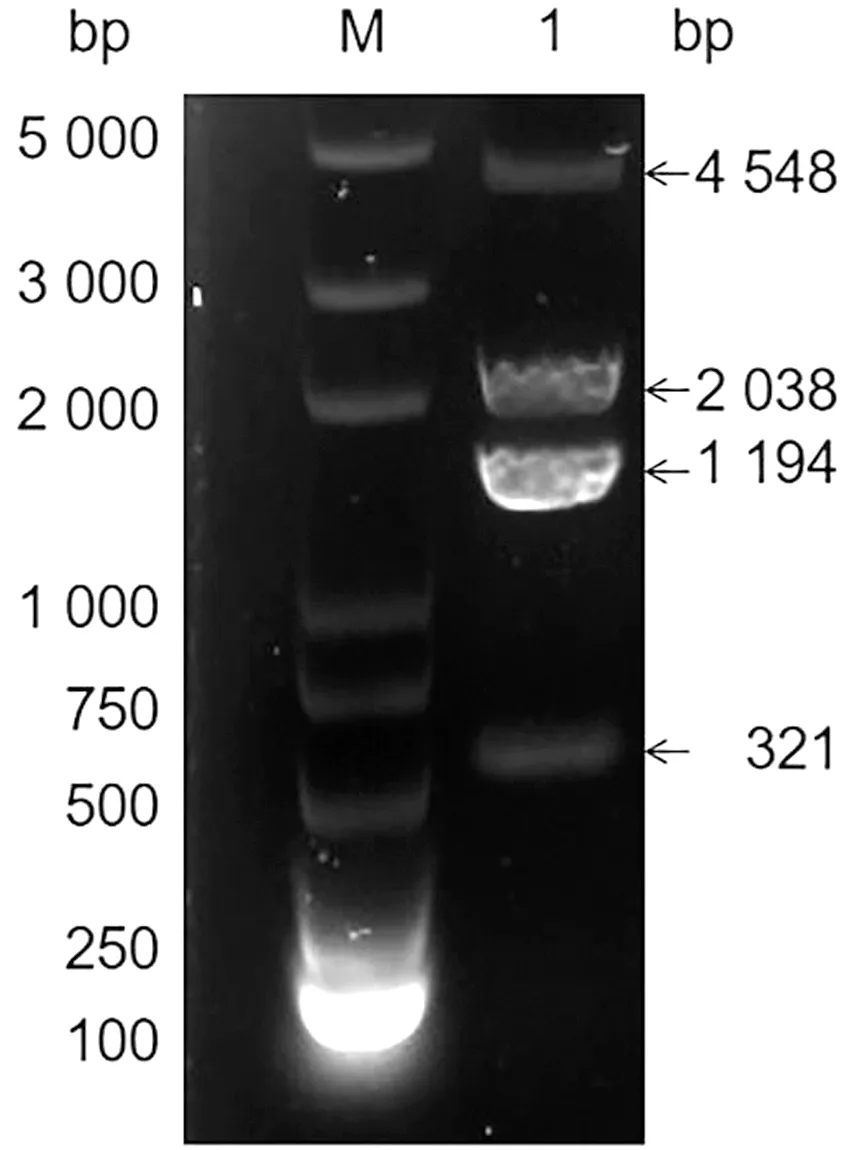
M.Trans2K® Plus DNA Marker;1.pSRS29C-DHF质粒酶切产物
2.3 基因敲除虫株的筛选与鉴定PCR鉴定电转后的虫株,结果显示,敲除株可扩增出797 bp的5′UTR片段和576 bp的3′UTR片段,与预期大小一致,未能扩增出用于鉴定的700 bp的SRS29C基因片段,野生株能扩增出此基因片段(图7)。经单克隆筛选的虫株PCR鉴定均与虫库鉴定结果相同(图8)。结果表明,已成功获得SRS29C基因敲除株,命名为弓形虫ME49△SRS29C及RH△SRS29C株。
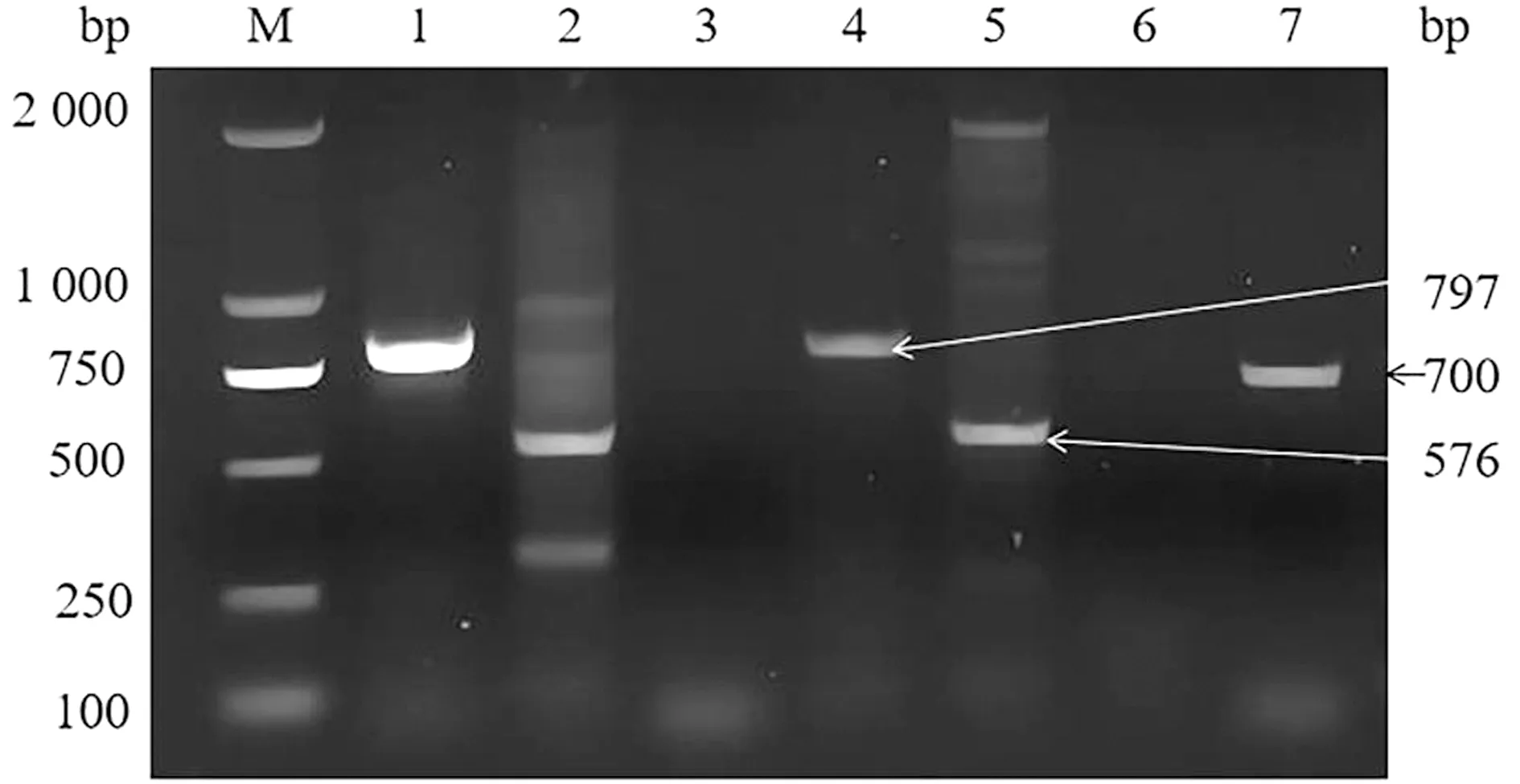
M.DL2000 DNA Marker;1.ME49△SRS29C PCR1结果;2.ME49△SRS29C PCR2结果;3.ME49△SRS29C PCR3结果;4.RH△SRS29C PCR1结果;5.RH△SRS29C PCR2结果;6.RH△SRS29C△SRS29C PCR3;7.弓形虫野生株PCR3结果
2.4 基因敲除型虫株的Western blot鉴定对弓形虫SRS29C基因敲除株与野生型虫株进行Western blot验证,结果显示在弓形虫ME49△SRS29C株、RH△SRS29C株、ME49株及RH株中,内参Tubulin抗体均能在50 kDa左右识别出1条特异性条带,与预期大小一致,目的基因SRS29C抗体在野生虫体中能识别出1条39 kDa左右的特异性条带,与SRS29C蛋白质大小一致,但在ME49△SRS29C株与RH△SRS29C株中未能识别出条带(图9)。结果表明,弓形虫ME49△SRS29C株与RH△SRS29C株不能表达SRS29C蛋白,进一步确定基因敲除型虫株构建成功。
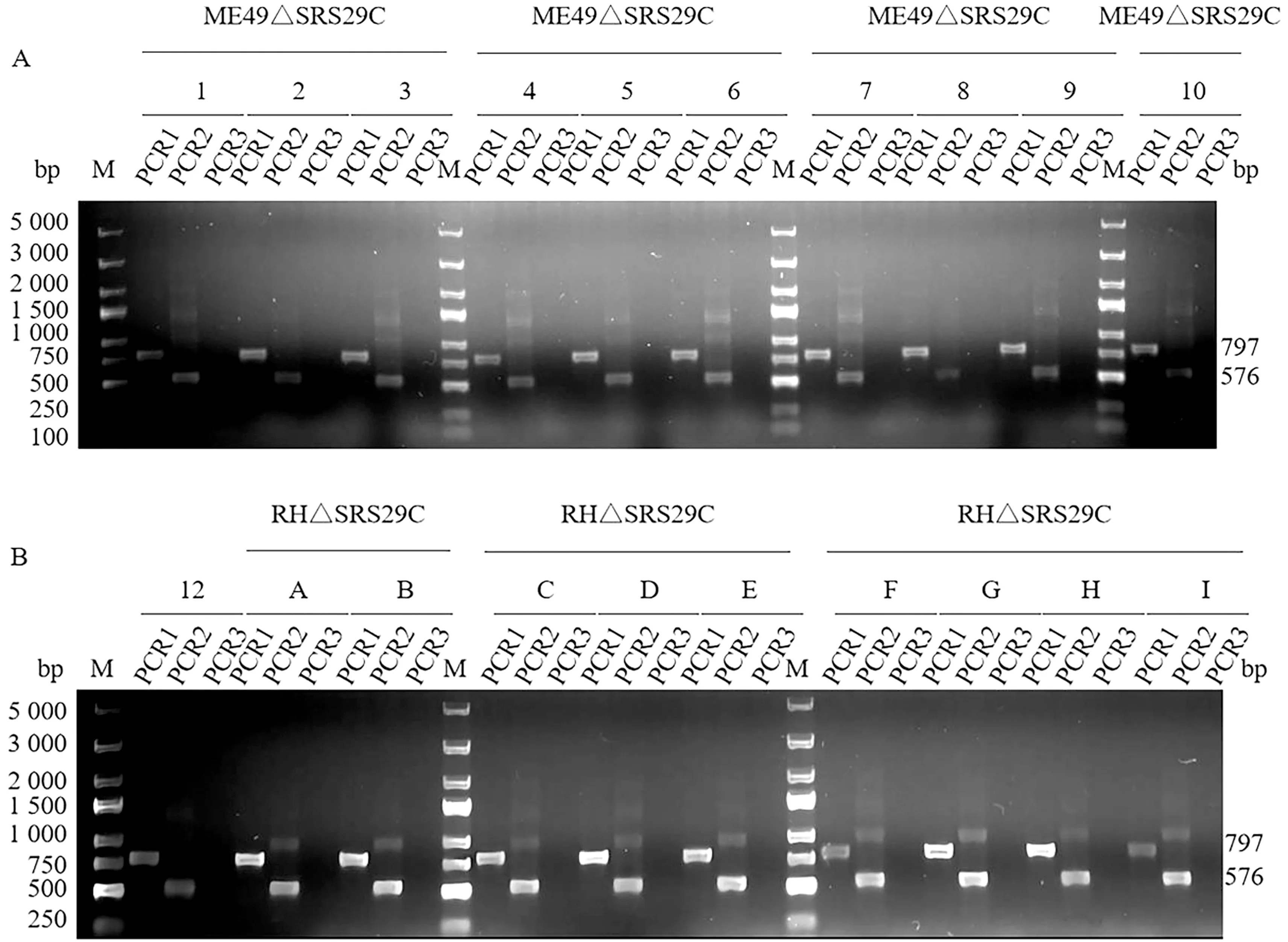
A.ME49△SRS29C-1~10号基因敲除单克隆株PCR鉴定结果(M.DL5000 DNA Marker);B.ME49△SRS29C-12及RH△SRS29C-A-I号基因敲除单克隆株PCR 鉴定结果(M.DL5000 DNA Marker)
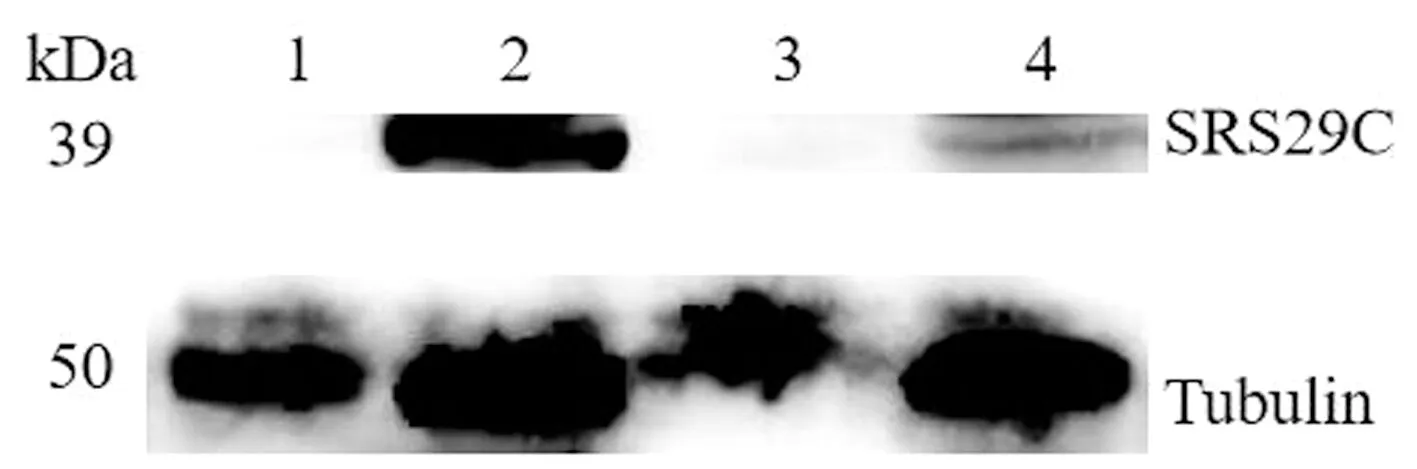
1~4.弓形虫ME49△SRS29C株、ME49株、RH△SRS29C株、RH株全虫蛋白
3 讨论
CRISPR/Cas9技术在弓形虫的研究领域应用广泛,如参与弓形虫入侵的微线体蛋白MIC10、MIC11[11-12]、致密颗粒蛋白GRA16[13]、棒状蛋白ROP5和ROP18[14]等。在弓形虫代谢相关研究方面,如糖酵解途径涉及的3-磷酸甘油酸激酶1,3-磷酸甘油醛脱氢酶2,烯醇酶2[15]、核苷酸合成过程中腺苷酸琥珀酸裂解酶和腺苷酸激酶[16]、氨基甲酰磷酸合成酶 Ⅱ[17]等,使得弓形虫的研究越来越深入。而弓形虫的毒性是复杂多样的,具有多种不同的调节因子,可协同发挥作用,使得不同种类的弓形虫具有不同的毒力,SRS蛋白家族在此方面起的作用可能多种多样。部分已报道的弓形虫SRS蛋白家族如TgSRS1对小鼠有一定的保护作用[18];TgSRS3具有良好的诊断效果[19];TgSRS47D蛋白被预测富含丰富的B、T细胞表位但并未进行具体研究[20];高俊莹等[21]研究发现,TgSRS-SAG1参与黏附宿主细胞表面的硫化肝素并促进虫体入侵。梁凯等[22]将弓形虫TgSRS-SAG1与TgSRS-SAG4重组蛋白单独及联合诱导昆明鼠,发现其均能诱导小鼠产生抗弓形虫感染的免疫保护作用,且联合免疫保护效果更佳。WASMUTH等[23]针对TgSRS-SRS29B基因及TgSRS-SRS34A基因,构建了ΔSRS29B、ΔSRS34A单敲除株以及ΔSRS29BΔSRS34A双敲除株,此后通过免疫印迹试验及流式细胞技术检测其他存在于速殖子期SRS家族成员的表达情况,两项试验结果均显示在ΔSRS29BΔSRS34A双敲除株中SRS29C表达量明显增高。而这种SRS29C表达量过高的虫株在体内毒性试验中展现了很好的保护力。因此,本试验构建了弓形虫表面抗原SRS29C基因缺失株,为弓形虫SRS29C基因功能的深入研究奠定了基础,有助于探索SRS蛋白家族之间是否存在代偿机制,以及为弓形虫疫苗的研发提供新的靶基因。
