水稻耐热新种质R203的创制与应用
2023-03-07刘刚夏快飞吴艳张明永张再君杨金松邱东峰
刘刚,夏快飞,吴艳,张明永,张再君,杨金松,邱东峰
水稻耐热新种质R203的创制与应用
刘刚1,夏快飞2,吴艳1,张明永2,张再君1,杨金松1,邱东峰1
1湖北省农业科学院粮食作物研究所/粮食作物种质创新与遗传改良湖北省重点实验室,武汉 430064;2中国科学院华南植物园,广州 510650
【目的】全球气候变暖导致水稻抽穗开花期受到高温热害的影响日益严重。鉴定抽穗期耐高温水稻种质,创制耐热新种质,为培育实用性耐热水稻新品种,降低高温热害对水稻生产的影响,保障中国乃至世界的粮食安全奠定基础。【方法】以广恢128(七桂早/测64//明恢63)为耐热亲本,通过杂交、复交以及系谱法,选育过程中连续多代筛选抽穗开花期处于高温阶段结实率较高且变异较小的株系,在高世代,利用人工气候室进行抽穗开花期耐热鉴定(处理盆栽于开花当天移入人工气候室,高温处理时间段为每日09:00—15:00,设置恒温38℃,15:01—次日08:59设置恒温28℃,相对湿度均为75%,处理7 d),结合农艺性状分析方法创制水稻耐热新种质。【结果】创制的新种质R203具有较好的耐热性,在正常条件下和高温条件下都有较高的结实率(常温结实率为94.5%、高温结实率为81.9%、相对结实率为86.7%),其对三系不育系恢复能力强,综合抗性好,稻米品质优,具有配制实际应用价值的耐热新组合的潜力。在自然高温条件下,以R203为父本、7个三系不育系为母本配制的杂交组合结实率为83.4%—99.4%,耐热性均表现较好。其中,育成的三系杂交中籼新品种泰优203的结实率为87.9%,综合相对耐热系数为1.11,耐热性达到1级,生产试验中,其产量比对照增加5.36%,增产点占85.71%,具有很好的丰产和稳产性,稻米品质达部标二级,具有较好的推广应用价值。【结论】当前耐热基础研究不足以支撑实用型耐热新品种选育,以高温湿热易发地区的种质为材料,通过表型选择创制了耐热新种质R203,并利用杂种优势,培育出实用型耐热水稻新品种泰优203。
水稻;耐热;种质创新;资源评价
0 引言
【研究意义】长江中下游地区为中国水稻主产区,水稻面积占全国50%以上。而高温热害是影响长江中下游水稻生产的主要灾害因子之一[1]。2013年IPCC第五次评估报告再次明确指出气候变暖比以前预测的更为严重[2],水稻生产将面临更为严峻的高温热害风险,尤其是开花期时的高温热害已成为中国水稻生产的主要自然灾害,因此,鉴定开花期耐高温水稻新种质、培育新品种,是降低高温热害对水稻生产影响的根本措施,对保障中国乃至世界粮食安全具有重要意义。【前人研究进展】水稻耐热性为多基因调控的数量性状,应用常规的遗传育种方法很难进行准确而有效的选择。近年来,随着测序技术的迅猛发展和功能基因组学研究的不断进步,水稻耐热分子机理研究取得较大进展。目前,已定位了165个耐热QTL[3-22],YE等[20]定位到高温胁迫下调控小穗育性的QTL(),其位于第4染色体,在开花期遭受高温胁迫时,能将小穗育性提高约15%。PS等[21]利用N22和IR64构建的重组自交系(recombinant inbred lines,RILs),以开花期小花育性和每株产量作为指标,定位到5个耐热性相关的QTL,分别位于第3、5、9和12染色体,能够解释6.27%—21.29%的表型变异。还有少数相关功能基因成功被克隆,Wang等[23]克隆了编码温度介导和节律调控的DEAD-Box RNA解旋酶的基因,该基因参与高温条件下正常rRNA前体的加工,是细胞核仁SSU复合体的伴侣蛋白,对高温下的细胞增殖十分重要,调控水稻耐热生长。Liu等[24]克隆了苗期耐高温基因,其编码环指蛋白,通过介导过氧化氢诱导的气孔关闭增强水稻耐热性。Zheng等[25]发现对高温下叶绿体发育以及PSⅡ功能的维持十分重要。Li等[26]研究表明通过有效降解有毒变性蛋白以及维持高温应答过程,进而保护植物细胞。Kan等[27]通过正向遗传学方法从水稻耐热遗传资源中定位克隆,其编码一个G蛋白γ亚基,负向调控水稻的耐热性。Zhang等[28]分离克隆水稻高温抗性新基因位点,揭示中存在由2个拮抗的基因和组成的遗传模块调控水稻高温抗性的新机制和叶绿体蛋白降解新机制。刘进等[22]检测到为新的主效QTL,对增强苗期耐热性具有较强的功效。【本研究切入点】目前,尽管已鉴定出一些水稻耐热相关QTL,成功克隆少部分耐热功能基因,但应用分子标记辅助选择创制耐热新种质,培育耐热新品种尚未见报道。水稻优良耐热种质资源也相对匮乏,虽已鉴定出的耐热水稻资源N22、T226、996、R1056、WD-16343等,但都不能直接应用于新品种选育,如N22是较早鉴定出来的耐高温材料,其常温结实率为63.70%[29],低于生产上要求的75.00%标准,不适宜于直接应用;WD-16343在高温处理结实率为71.00%,相对结实率高达98.50%,但其在常温条件下结实率仅为72.10%,且株叶形态、穗粒结构、稻米品质等较多性状不理想,无法直接利用[29-30]。因此,在现阶段利用表型选择的方法创制农艺性状优良的耐热水稻新种质对于培育耐热新品种显得尤为重要。【拟解决的关键问题】本研究在水稻耐高温种质和可用于耐高温辅助选择的分子标记匮乏情况下,开展抽穗开花期耐热性鉴定,结合农艺性状分析,筛选并创制实用型水稻耐热新种质,为培育水稻耐热新品种奠定基础。
1 材料与方法
1.1 材料创制与杂交配组方法
自2000年开始在广州高温湿热易发地区,先后以轮回422、IR54、广恢128、银晶软占等为亲本(电子附表1),通过杂交、复交法以及系谱法培育新材料,选育过程中,连续多代比较自然条件下结实率的高低,结合气象分析,判定年际间高温发生时段,筛选抽穗开花期处于高温阶段结实率较高且变异较小的株系。在高世代,利用人工气候室进行高温鉴定,筛选耐高温株系。利用杂交水稻配组方法配制杂交组合。
1.2 功能基因分析
运用含有重要农艺性状的功能基因标记和单倍型标记的高密度水稻基因芯片GSR40K(武汉双绿源创芯科技研究院有限公司)进行功能基因分析。
1.3 耐热性鉴定与分析
2014年对80份资源以及创制材料进行人工气候室抽穗期耐热性鉴定。每份材料分为2个处理,处理1田间自然条件作为常温对照,2014年7月15日至8月15日平均最高气温32.5℃,平均最低气温24.3℃,平均相对湿度79%;处理2盆栽于开花当天移入人工气候室进行7d高温处理,高温处理时间段为每日09:00—15:00,设置恒温38℃,15:01—次日08:59设置恒温28℃,相对湿度均为75%。通过高温结实率、常温结实率的统计和相对结实率来评价耐热性,具体方法见参考文献[29-30]。
2019年在华中农业大学开展耐热鉴定,分为田间自然高温鉴定与玻璃温室高温鉴定,以高温下鉴定品种的结实率与对照品种(丰两优四号)的结实率之比作为相对耐热系数,耐热性分级以相对耐热系数高低为评价标准:强(≥1.1)、较强(0.9≤<1.1)、一般(0.7≤<0.9)、较弱(0.5≤<0.7)、弱(<0.5)。利用DSR-TH数字化温湿度记录仪(ZOGLAB Microsystem Co.Ltd)每5分钟实时记录温、湿度。田间自然高温鉴定按分期播种方法进行,在抽穗时按见穗期对分蘖分别挂牌,成熟时按见穗期将挂牌分蘖进行分类,分别考察结实率,按自然条件日均温32℃以上、最高温在35℃以上且持续时间5 d以上为受到高温胁迫,以这部分分蘖的平均结实率作为高温胁迫条件的结实率,以其他非高温时间段的最高结实率作为常温条件下的结实率;玻璃温室高温鉴定时,利用通风口闭(开)达到增(减)温效果,试验处理日均温32.2—34.5℃,日最高温在38.9—41.0℃,每个品种栽4盆,每盆保留3个抽穗一致的大分蘖,2盆不处理计算自然结实率,2盆处理计算高温结实率。
1.4 田间试验方法与分析
按文献[31]的方法调查播种期、始穗期等农艺性状,成熟后每小区连续抽样10株考种,用于分析各品种的产量性状。结合温度等气象数据(http://www.tianqihoubao. com/lishi/)分析各品种在正常条件和自然高温条件下的结实率和耐热性。采用EXCEL完成方差分析。
2 结果
2.1 R203的创制及配组能力分析
2000年,水稻耐热性种质资源的缺乏,耐热性鉴定尚处于起步阶段。选用轮回422为母本,IR54为父本进行杂交,后代按系谱法选育,至2003年得到中间材料R10;2003年以广恢128为母本,R10为父本进行杂交,后代按系谱法选育恢复系,至2006年得到中间材料R145;2006年以银晶软占为母本,以R145为父本进行杂交,2007年早造种植F1并进行混合收种,2007年晚造种植F2进行单株筛选,在2008-2010连续3年做早、晚造种植,比较自然条件下结实率高低,并结合气象数据分析,筛选抽穗开花期处于高温阶段结实率较高且变异较小的株系,至2011年早季获得主要农艺性状基本稳定的株系2011203。筛选株型紧凑,叶色清秀,后期自然褪色黄熟,稻米外观透明,垩白少的单株并扩繁,定名为R203,详细过程如图1所示。R203于2021年申请新品种保护,申请公告号20211006360。
2020—2021年,将R203与多个三系不育系配制的杂交组合作中稻种植于武汉,于5月10日播种,以丰两优四号为对照,各组合均在8月10日前始穗,其考种产量数据结果见表1。根据http://tianqi.2345. com/wea_history/57494.htm显示的武汉市历史天气,2020—2021年7月24日—8月18日同期日最高温度分别为23—37℃、21—38℃,日平均气温分别为24—32℃、22.5—32.5℃,气温正常。R203对各三系不育系恢复性较好,各组合结实率分布在76.77%—90.24%,各组合理论产量分布在7.68—13.16 t·hm-2,有8个组合的理论产量超过对照品种3.00%以上,且有6个组合比对照增产5.00%以上,增产极显著,表明R203具有较好的配组潜力。其中,以三系不育系泰丰A为母本,以R203为父本配组的杂交组合泰优203,结实率达89.03%,产量达9.83 t·hm-2,超过对照丰两优四号,增产9.48%,表现优异(表1)。
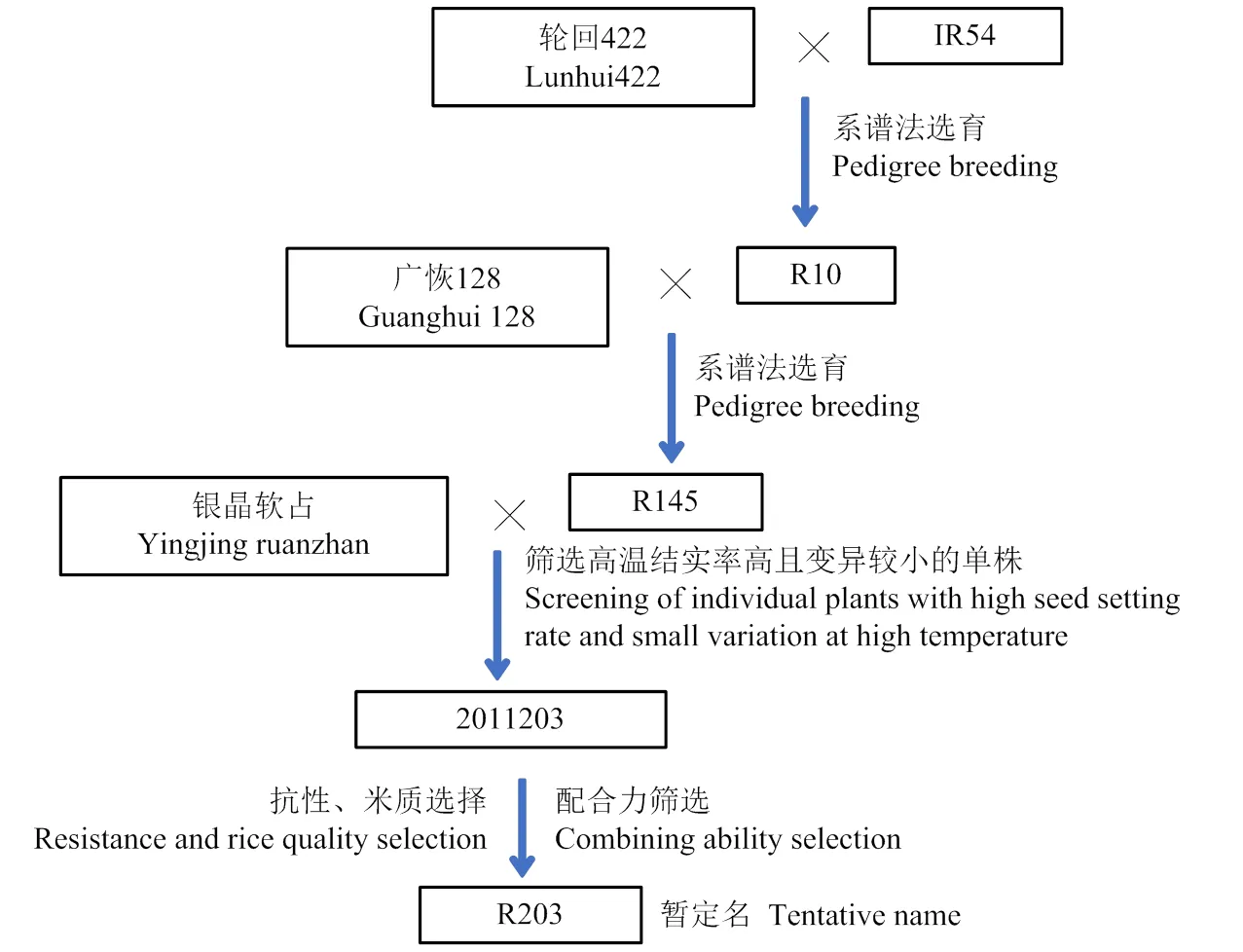
图1 R203的系谱图

表1 R203与不同三系不育系配组的产量及相关性状
*:参试品种与对照品种的理论产量差异显著(0.01<≤0.05);**:参试品种与对照品种的理论产量差异极显著(≤0.01)
*: significant difference in theoretical yield between tested varieties and control varieties in 0.05 probability level; **: significant difference in theoretical yield between tested varieties and control varieties in 0.01 probability level
2.2 R203的功能基因分析
依据发表文章结果,以及具体功能的验证,利用武汉双绿源的功能基因芯片对R203进行了部分功能基因分析,共检测到25个功能基因,包括产量基因2个、抗非生物逆境基因2个、抗生物逆境基因7个、品质基因7个、生育期基因3个、株型基因2个、育性基因1个、落粒性基因1个和耐热基因1个(表2)。同时未检测出耐热基因。结果表明,田间观察R203叶色清秀、综合抗性好,以及稻米外观品质好、垩白少等性状优良具有较好的分子基础。
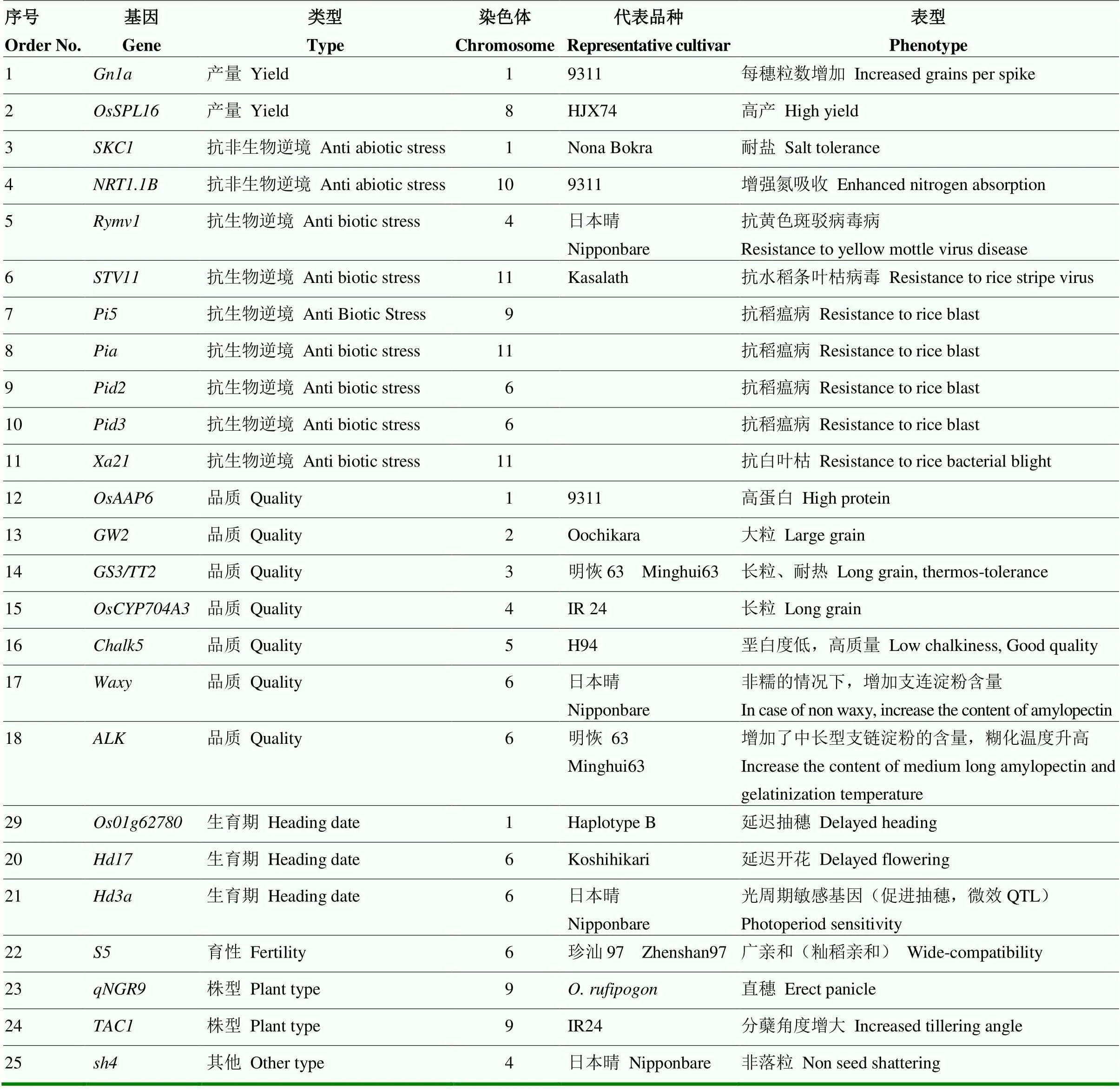
表2 R203的部分功能基因列表
2.3 R203及泰优203的耐热性鉴定
根据查中萍等[29]报道,抽穗期每日9:00—15:00,人工气候室38℃高温、相对湿度75%处理7 d,80份材料(以N22为对照)的常温结实率、高温结实率分别为54.00%—94.50%、0—83.00%,相对结实率分布范围为0—98.80%,说明高温处理有效。结果显示,对照材料N22的常温结实率、高温结实率、相对结实率分别为85.00%、63.70%和74.90%,资源材料WD-16343分别为72.10%、71.00%和98.50%,而R203(田间编号44079)则分别为94.50%、81.90%和86.70%,其常温结实率和高温结实率均显著高于N22和WD-16343,说明R203不仅具有较好的耐热性,还具有较高的利用价值。
以R203为父本配制的杂交组合在自然高温条件下结实率正常。2022年湖北武汉经历夏季异常高温,根据气象记载,自7月24日至8月23日,日均气温26.5—34℃,自7月31日至8月23日,最高气温均在35℃以上,有连续15 d的最高气温超过38℃,连续18 d的最低气温高于28℃,日均温33℃,这样的高温强度超过文献[29]的高温鉴定强度。通过田间调查及考种,R203配组的杂交组合与对照组合泰丰A×元恢236的抽穗期及结实率情况如表3和图2所示。各杂交组合在7月底到8月上旬抽穗,遭遇极端高温天气,对照泰丰A×元恢236的结实率仅为50.00%,以R203配组的杂交组合结实率均高于75.00%且高于对照,差异极显著。说明以R203配制的杂交组合具有较好的耐热性。通过对比泰丰A×元恢236与泰优203(泰丰A×R203),其结实率的差异可视为父本的影响,说明亲本的耐热性差异会导致杂交组合的耐热性不同,因此,在杂交配组中应该选用耐热性较好的亲本,才能培育出耐热性好的杂交水稻,同时,也说明R203在水稻耐热育种中具有重要价值。

表3 2022年不同杂交组合的抽穗期及在高温条件下结实率
杂交组合泰优203在参加长江中下游汇丰企业联合体中籼迟熟组区试中由华中农业大学开展了耐热性鉴定。2019年和2021年的耐热性鉴定分别以丰两优四号为对照,其中,2021年4个品种耐热级别均为3级,综合相对耐热系数小于对照或与对照差异不显著,表明其耐热性不强于对照相当。2019年鉴定结果显示,两优新月丝苗和桂香优086的综合相对耐热系数小于对照丰两优四号,表明它们的耐热性弱于对照,泰优203的综合耐热系数为1.11,显著高于对照,泰优203的耐热性级别为1级,为参试品种中最好,且其在大田高温条件下的结实率最高,达到76.74%,超过国家审定标准的最低结实率(≥75.0%,表4),说明泰优203在耐热性方面表现优异。
2.4 泰优203的利用价值分析
2017年夏季在湖北武汉进行组合筛选试验,泰优203平均产量为10.24 t·hm-2,比对照丰两优四号增产2.26%,全生育期133 d,比对照短1.2 d;株高119.0 cm,有效穗数261.00万/hm2,每穗总粒数212粒,结实率78.00%,千粒重24.90 g,谷粒长宽比为3.95。2018年进行品种比较试验,平均产量为10.33 t·hm-2,比对照丰两优四号增产1.72%;全生育期134.0 d,与对照相当;株高122.0 cm,有效穗数273.00万/hm2,每穗总粒数209粒,结实率81.30%,千粒重23.70 g,谷粒长宽比为4.00。综合结果表明泰优203分蘖力强,成穗率高,产量稳定,后期熟相好,稻米外观品质好。

A:1070A×R203;B:垦2001S×R203;C:EK2S×R203;D:N25S×R203;E:泰香A×R203;F:野香A×R203;G:泰优203;H:泰丰A×元恢236(CK)
泰优203于2019—2020年参加长江中下游中汇丰企业水稻联合体籼迟熟组区试,区试点18个,分布在安徽、福建、河南、湖北、湖南、江苏、江西和浙江8个省(表5)。2年产量分别比对照丰两优四号增产6.70%和3.35%,增产点率分别为100.00%和94.12%,全生育期131.4 d,比对照短2.3 d。2021年开展生产试验,产量比对照增加5.36%,7个点中有6个点增产,增产点占85.71%。主要农艺性状表现:有效穗数256.50万/hm2,株高115.7 cm,穗长24.7 cm,每穗总粒数187.1粒,结实率88.10%,千粒重24.00 g。抗性:稻瘟病综合指数4.4级,穗瘟损失率最高级5级;白叶枯病7级;褐飞虱9级。米质主要指标:出糙率79.00%,精米率69.50%,整精米率65.50%,粒长6.7 mm,长宽比3.50,垩白粒率7.00%,垩白度2.50%,直链淀粉含量15.30%,胶稠度62.0 mm,碱消值6.4,透明度1级。稻米品质部标优二级。
多年多点试验数据综合显示泰优203产量高,增产点率高,表明其稳产性好,稻米品质优良,且耐热1级,具有较大的推广利用价值。泰优203于2021年申请新品种保护,申请公告号20211004973,于2022年通过国家农作物品种审定(国审稻20220178)。
3 讨论
3.1 耐热新种质R203具有实际应用价值和基础研究价值
本研究创制的耐热新种质R203,一方面具有很好的实际生产应用价值,亲本主要来源于南方稻区的种质资源,且主要培育地在广州,连续多代筛选抽穗开花期处于高温阶段结实率较高且与正常温度条件下变异较小的株系,使培育的R203不仅有较好的耐热性,在正常条件下和高温条件下都有较高的结实率,其农艺性状、稻米品质和综合抗性等方面均达到生产应用标准,为培育耐热新品种奠定了基础。已经育成的三系杂交中籼新品种泰优203,耐热性1级,且丰产性和稳产性达到生产利用的标准,稻米品质部标二级,具有较好的推广应用价值。另一方面,具有很好的基础研究价值。经检测最近克隆的水稻耐热基因[25]和[26]在R203中的基因型,表明R203不含有基因型且具有的热敏感基因型,同样含有基因型的明恢63在耐热性鉴定中表现为热敏感[29, 32],可见R203的耐热性或有新的基因型控制,可用来定位或克隆有实用价值的水稻开花期耐热新QTL/基因,解析水稻耐热分子机理,为水稻耐热的分子标记辅助选择育种提供基因资源。

表4 长江中下游汇丰企业联合体中籼迟熟组生产试验参加品种耐高温鉴定结果
数据来源于区试结果;**:泰优203与丰两优四号差异极显著(≤0.01) Data from regional trials; **: significant difference between Taiyou203 and Fengliangyousihao at≤0.01
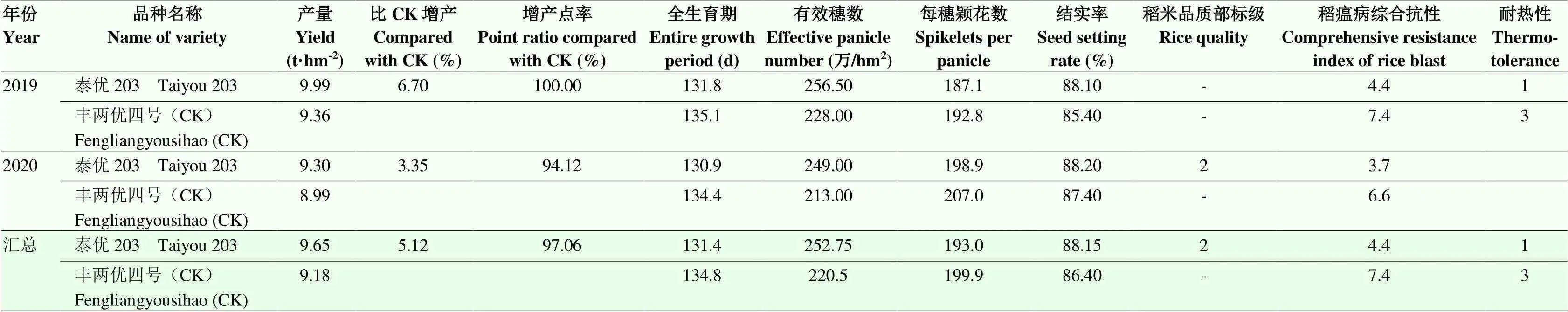
表5 泰优203在区试中的产量及主要性状表现
区试汇总数据的算法:2年数据产量及相关性状取均值、稻米品质取最优值、稻瘟病抗性取最差值,耐热性鉴定仅开展1次
According to the district test standard, the algorithm of comprehensive value is respectively: the mean value of yield and related characters, the optimal value of rice quality and the worst value of rice blast resistance were taken as the two years' data. The heat resistance was identified only once
3.2 表型选择创制新种质提高水稻耐热选择的准确性
尽管目前已鉴定到一些水稻耐热相关QTL,但是水稻耐热功能基因的分离克隆还较少,且克隆的基因多数局限于分子机理研究,也鲜见有效利用分子标记辅助选择创制耐热种质、培育耐热新品种的报道。这主要有以下原因:一是水稻耐热性为多基因控制的数量性状,遗传机理复杂;二是热胁迫对水稻各发育时期都会造成影响,不同时期的水稻对热胁迫的敏感性不同,因而造成的影响也不同,这导致难以对水稻耐热性进行精确鉴定;三是水稻耐热性受多种环境条件影响,鉴定条件较难控制。通过表型选择创制水稻耐热新种质,性状鉴定准确可靠,不仅可以培育实用性强的耐热新品种解决生产实际问题,还可以耐热新种质创制为突破口,在优异耐热新种质的基础上开展实用的、关键的耐热基因的定位与克隆,挖掘其优异等位基因,同时可加强表型选择与基因型选择相互印证,提高耐热选择的准确性。
4 结论
采用耐热亲本广恢128等高温湿热易发地区的种质资源创制耐热种质,选育过程中连续多代筛选抽穗开花期处于高温阶段结实率较高且变异较小的株系,在高世代利用人工气候室进行抽穗开花期耐热鉴定,结合农艺性状分析,创制了实用型耐热新种质R203,并以此为基础,利用杂种优势,培育出丰产、稳产、优质的耐热水稻新品种泰优203。
[1] CHEN C Q, VAN GROENIGEN K J, YANG H Y, HUNGATE B A, YANG B, TIAN Y L, CHEN J, DONG W J, HUANG S, DENG A X, JIANG Y, ZHANG W J. Global warming and shifts in cropping systems together reduce China’s rice production. Global Food Security, 2020, 24: e100359.
[2] IPCC. Climate Change 2013: The physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2013, Cambridge, United Kingdom and New York, NY, USA, 1535.
[3] JAGADISH S V K, CAIRNS J, LAFITTE R, WHEELER T R, PRICE A H, CRAUFURD P Q. Genetic analysis of heat tolerance at anthesis in rice. Crop Science, 2010, 50(5): 1633-1641.
[4] JAGADISH S V K, MUTHURAJAN R, OANE R, WHEELER T R, HEUER S, BENNETT J, CRAUFURD P Q. Physiological and proteomic approaches to address heat tolerance during anthesis in rice (L.). Journal of Experimental Botany, 2010, 61(1): 143-156.
[5] KILASI N L, SINGH J, VALLEJOS C E, YE C R, JAGADISH S V K, KUSOLWA P, RATHINASABAPATHI B. Heat stress tolerance in rice (L.): identification of quantitative trait loci and candidate genes for seedling growth under heat stress. Frontiers in Plant Science, 2018, 9: 1578.
[6] LEI D Y, TAN L B, LIU F X, SUN C Q. Identification of heat- sensitive QTL derived from common wild rice (Griff.). Plant Science, 2013, 201/202: 121-127.
[7] LI M M, LI X, YU L Q, WU J W, LI H, LIU J, MA X D, JO S M, PARK D S, SONG Y C, SHIN D J, HAN L Z. Identification of QTLs associated with heat tolerance at the heading and flowering stage in rice (L.). Euphytica, 2018, 214(4): 1-11.
[8] POLI Y, BASAVA R K, PANIGRAHY M, VINUKONDA V P, DOKULA N R, VOLETI S R, DESIRAJU S, NEELAMRAJU S. Characterization of a Nagina22 rice mutant for heat tolerance and mapping of yield traits. Rice (New York, NY), 2013, 6(1): 36.
[9] TAZIB T, KOBAYASHI Y, KOYAMA H, MATSUI T. QTL analyses for anther length and dehiscence at flowering as traits for the tolerance of extreme temperatures in rice (L.). Euphytica, 2015, 203(3): 629-642.
[10] XIAO Y H, PAN Y, LUO L H, ZHANG G L, DENG H B, DAI L Y, LIU X L, TANG W B, CHEN L Y, WANG G L. Quantitative trait loci associated with seed set under high temperature stress at the flowering stage in rice(L.). Euphytica, 2011, 178(3): 331-338.
[11] YE C R, TENORIO F A, ARGAYOSO M A, LAZA M A, KOH H J, REDOÑA E D, JAGADISH K S V, GREGORIO G B. Identifying and confirming quantitative trait loci associated with heat tolerance at flowering stage in different rice populations. BMC Genetics, 2015, 16: 41.
[12] ZHU S, HUANG R L, WAI H P, XIONG H L, SHEN X H, HE H H, YAN S. Mapping quantitative trait loci for heat tolerance at the booting stage using chromosomal segment substitution lines in rice. Physiology and Molecular Biology of Plants, 2017, 23(4): 817-825.
[13] 曹立勇, 朱军, 赵松涛, 何立斌, 颜启传. 水稻籼粳交DH群体耐热性的QTLs定位. 农业生物技术学报, 2002, 10(3): 210-214.
CAO L Y, ZHU J, ZHAO S T, HE L B, YAN Q C. Mapping QTLs for heat tolerance in a DH population fromcross of rice (). Journal of Agricultural Biotechnology, 2002, 10(3): 210-214. (in Chinese)
[14] 曹志斌, 谢红卫, 聂元元, 毛凌华, 李永辉, 蔡耀辉. 水稻抽穗扬花期耐热QTL ()定位及其遗传效应分析. 中国水稻科学, 2015, 29(2): 119-125.
CAO Z B, XIE H W, NIE Y Y, MAO L H, LI Y H, CAI Y H. Mapping a QTL () for heat tolerance at the heading stage on rice chromosome 5 and its genetic effect analysis. Chinese Journal of Rice Science, 2015, 29(2): 119-125. (in Chinese)
[15] 陈庆全, 余四斌, 李春海, 牟同敏. 水稻抽穗开花期耐热性QTL的定位分析. 中国农业科学, 2008, 41(2): 315-321.
CHEN Q Q, YU S B, LI C H, MOU T M. Identification of QTLs for heat tolerance at flowering stage in rice. Scientia Agricultura Sinica, 2008, 41(2): 315-321. (in Chinese)
[16] 奎丽梅, 谭禄宾, 涂建, 卢义宣, 孙传清. 云南元江野生稻抽穗开花期耐热QTL定位. 农业生物技术学报, 2008, 16(3): 461-464.
KUI L M, TAN L B, TU J, LU Y X, SUN C Q. Identification of QTLs associated with heat tolerance of Yuanjiang common wild rice (griff.) at flowering stage. Journal of Agricultural Biotechnology, 2008, 16(3): 461-464. (in Chinese)
[17] 盘毅, 罗丽华, 邓化冰, 张桂莲, 唐文邦, 陈立云, 肖应辉. 水稻开花期高温胁迫下的花粉育性QTL定位. 中国水稻科学, 2011, 25(1): 99-102.
PAN Y, LUO L H, DENG H B, ZHANG G L, TANG W B, CHEN L Y, XIAO Y H. Quantitative trait loci associated with pollen fertility under high temperature stress at flowering stage in rice. Chinese Journal of Rice Science, 2011, 25(1): 99-102. (in Chinese)
[18] 张昌泉, 陈飞, 洪燃, 李钱峰, 顾铭洪, 刘巧泉. 利用染色体片段代换系定位水稻抽穗开花期耐热性QTL. 江苏农业科学, 2016, 44(12): 120-123.
ZHANG C Q, CHEN F, HONG R, LI Q F, GU M H, LIU Q Q. Mapping QTL for heat tolerance at heading and anthesis stage in rice using chromosome segment substitution lines. Jiangsu Agricultural Sciences, 2016, 44(12): 120-123. (in Chinese)
[19] 赵志刚, 江玲, 肖应辉, 张文伟, 翟虎渠, 万建民. 水稻孕穗期耐热性QTLs分析. 作物学报, 2006, 32(5): 640-644.
ZHAO Z G, JIANG L, XIAO Y H, ZHANG W W, ZHAI H Q, WAN J M. Identification of QTLs for heat tolerance at the booting stage in rice (L.). Acta Agronomica Sinica, 2006, 32(5): 640-644.(in Chinese)
[20] YE C R, TENORIO F A, REDOÑA E D, MORALES-CORTEZANO P S, CABREGA G A, JAGADISH K S V, GREGORIO G B. Fine-mapping and validatingto increase spikelet fertility under heat stress at flowering in rice. Theoretical and Applied Genetics, 2015, 128(8): 1507-1517.
[21] PS S, SV A M, PRAKASH C, MK R, TIWARI R, MOHAPATRA T, SINGH N K. High resolution mapping of QTLs for heat tolerance in rice using a 5K SNP array. Rice (New York, NY), 2017, 10: 28.
[22] 刘进, 胡佳晓, 马小定, 陈武, 勒思, Jo Sumin, 崔迪, 周慧颖, 张立娜, Shin Dongjin, 黎毛毛, 韩龙植, 余丽琴. 最新录用:水稻RIL群体高密度遗传图谱的构建及苗期耐热性QTL定位. 中国农业科学, 2022, 55(22): 4327-4341.
LIU J, HU J X, MA X D, CHEN W, LE S, JO S, CUI D, ZHOU H Y, ZHANG L N, SHIN D, LI M M, HAN L Z, YU L Q. Construction of high density genetic map for RIL population and QTL analysis of heat tolerance at seedling stage in rice (L.). Scientia Agricultura Sinica, 2022, 55(22): 4327-4341. (in Chinese)
[23] WANG D, QIN B X, LI X, TANG D, ZHANG Y E, CHENG Z K, XUE Y B. Nucleolar DEAD-box RNA helicaseregulates thermotolerant growth as a pre-rRNA chaperone in rice. PloS Genetics, 2016, 12(2): e1005844.
[24] LIU J P, ZHANG C C, WEI C C, LIU X, WANG M G, YU F F, XIE Q, TU J M. The RING finger ubiquitin E3 ligaseenhances heat tolerance by promoting H2O2-induced stomatal closure in rice. Plant Physiology, 2015, 170(1): 429-443.
[25] ZHENG K L, ZHAO J, LIN D Z, CHEN J Y, XU J L, ZHOU H, TENG S, DONG Y J. The ricegene encoding a novel Degprotease protein is essential for chloroplast development under high temperatures. Rice (New York, NY), 2016, 9(1): 13.
[26] LI X M, CHAO D Y, WU Y, HUANG X H, CHEN K, CUI L G, SU L, YE W W, CHEN H, CHEN H C, DONG N Q, GUO T, SHI M, FENG Q, ZHANG P, HAN B, SHAN J X, GAO J P, LIN H X. Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice. Nature Genetics, 2015, 47(7): 827-833
[27] KAN Y, MU X R, ZHANG H, GAO J, SHAN J X, YE W W, LIN H X.controls rice thermotolerance through-dependent alteration of wax biosynthesis. Nature Plants, 2022, 8(1): 53-67.
[28] ZHANG H, ZHOU J F, KAN Y, SHAN J X, YE W W, DONG N Q, GUO T, XIANG Y H, YANG Y B, LI Y C, ZHAO H Y, YU H X, LU Z Q, GUO S Q, LEI J J, LIAO B, MU X R, CAO Y J, YU J J, LIN Y S, LIN H X. A genetic module at one locus in rice protects chloroplasts to enhance thermotolerance. Science, 2022, 376(6599): 1293-1300.
[29] 查中萍, 殷得所, 万丙良, 焦春海. 水稻种质资源开花期耐热性分析. 湖北农业科学, 2016, 55(1): 17-19.
ZHA Z P, YIN D S, WAN B L, JIAO C H. Analyzing of rice germplasm heat resistance during flowering stage. Hubei Agricultural Sciences, 2016, 55(1): 17-19. (in Chinese)
[30] LIU G, ZHA Z P, CAI H Y, QIN D D, JIA H T, LIU C Y, QIU D F, ZHANG Z J, WAN Z H, YANG Y Y, WANG B L, YOU A Q, JIAO C H. Dynamic transcriptome analysis of anther response to heat stress during anthesis in thermotolerant rice (L.). International Journal of Molecular Sciences, 2020, 21: 1155.
[31] 邱东峰, 葛平娟, 刘刚, 杨金松, 陈建国, 张再君. 优质水稻新种质ZY56 的创制及评价. 中国农业科学, 2021, 54(6): 1081-1091.
QIU D F, GE P J, LIu G, Yang J S, Chen J G, ZHANG Z J. Breeding and evaluation of elite rice line ZY56. Scientia Agricultura Sinica, 2021, 54(6): 1081-1091. (in Chinese)
[32] 周伟辉, 薛大伟, 张国平. 高温胁迫下水稻叶片的蛋白响应及其基因型和生育期差异. 作物学报, 2011, 37(5): 820-831.
ZHOU W H, XUE D W, ZHANG G P. Protein response of rice leaves to high temperature stress and its difference of genotypes at different growth stage. Acta Agronomica Sinica, 2011, 37(5): 820-831. (in Chinese)
Breeding and Application of a New Thermo-tolerance Rice Germplasm R203
LIU Gang1, XIA KuaiFei2, WU Yan1, ZHANG MingYong2, ZHANG ZaiJun1, YANG JinSong1, QIU DongFeng1
1Food Crop Institute, Hubei Academy of Agricultural Sciences/Hubei Key Laboratory of Food Crop Germplasm and Genetic Improvement, Wuhan 430064;2South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650
【Objective】The global warming has led to the increasingly serious heat damage on the heading and flowering stage of rice. To reduce the impact of heat damage on rice production and to ensure food security in China and even the world, new rice germplasms with thermo-tolerance on heading stage should be identified and new thermo-tolerance varieties need be bred. 【Method】Guanghui 128 (Qiguizao/Ce64//Minghui 63) was used as the heat resistant parent, through hybridization, multiple crossing and pedigree selection, the lines with high seed setting rate and small variation on heading and flowering stage during the high temperature were screened out for several generations’ breeding process. Then the selected higher generation lines were identified to create new thermos-tolerance rice germplasms inartificial climate chamber (The treated plants will be moved into the chamber on the flowering day, high temperature treatment is 9:00-15:00, 38℃, 15:01-8:59 28℃, the relative humidity is 75%, and the treatment lasts for 7 days), with analysis of agronomic trait. 【Result】The new germplasm R203 has stronger thermo-tolerance and higher seed setting rates under both normal and high temperature conditions (94.5% at normal temperature, 81.9% at high temperature, and 86.7% at relative). Its agronomic traits, quality and comprehensive resistance all meet the production standards. Above all, R203 has the potential to breed new thermos-tolerance hybrid rice varieties. The seed setting rates of 7 hybrid combinations with R203 as the male parent and seven three-line male sterile lines as the female parent were between 83.4%-99.4% under natural high temperature conditions. Among them, Taiyou 203, a new three-line medium indica hybrid rice has good qualities, the seed setting rate was 87.9%, the comprehensive relative heat resistance coefficient was 1.11, and the heat resistance reached level 1. In the production test, the yield increased by 5.36% compared with the control, and the yield increase point accounted for 85.71%. It has good high and stable yield, and the rice quality reached the second level of the ministerial standard. Thus Taiyou 203 has good promotion and application value. 【Conclusion】Currently, basic research on heat resistance is not enough to support the breeding of new practical heat resistant varieties,the rice resources in areas prone to high temperature and humidity are preferred as materials for breeding new heat tolerance lines,a new heat-resistant rice variety R203 was created by phenotypic selection, and a practical heat-resistant rice variety Taiyou 203 was developed by using heterosis.
rice; thermo-tolerance; germplasm enhancement; evaluation of germplasm

10.3864/j.issn.0578-1752.2023.03.001
2022-09-08;
2022-11-08
国家重点研发计划(2016YFD0100101-05)、国家作物种质资源库(湖北分库)(NICGR2022-31)
刘刚,Tel:18627862923;E-mail:liug1112@163.com。通信作者邱东峰,Tel:18672779158;E-mail:qdflcp@163.com
(责任编辑 李莉)
