不同秋眠等级苜蓿根系性状对越冬的影响
2023-02-07王晓龙杨曌来永才李红钟鹏徐艳霞柴华李莎莎吴玥宋敏超周景明
王晓龙,杨曌,来永才,李红,钟鹏,徐艳霞,柴华,李莎莎,吴玥,宋敏超,周景明
(1.黑龙江省农业科学院博士后科研工作站,黑龙江 哈尔滨 150086;2.黑龙江省农业科学院畜牧兽医分院,黑龙江 齐齐哈 161005;3.黑龙江省农业科学院,黑龙江 哈尔滨 150086)
紫花苜蓿(Medicago sativa)是多年生豆科牧草,具有产量高、抗逆性强、适应性广等优良特性,已成为奶牛等草食动物的优质饲草[1]。紫花苜蓿种植面积的不断扩大,有利于积极推进“粮改饲”及“种养结合”模式试点,适当的利用部分耕地来种植青贮玉米(Zea mays)和紫花苜蓿,可以优先满足区域内的饲草供求,进而促进苜蓿产业持续、健康、稳定发展[2]。黑龙江省西部地区处于中温带,土壤肥沃,地势平坦,水源充沛,拥有全省得天独厚的黑土地资源,是发展苜蓿产业的重要区域,但由于该地区冬季低温、暴雪、寒潮等极端天气时有发生,苜蓿易遭受冷害或冻害威胁,导致苜蓿生产能力下降,越冬耐寒能力降低,对草牧业可持续生产造成巨大损失[3]。因此,苜蓿根系的越冬问题一直是影响我国北方一些极寒地区苜蓿产业发展的关键问题。
秋眠性(fall dormancy)是苜蓿对晚秋环境(日照时数减少)变化的适应性反应,植株表现为由向上生长转向匍匐生长,致使总产量减少的一种生长特性,它与产量和越冬性紧密相关,已经成为北美评定苜蓿品种生产性能的重要参考指标[2]。根系是养分和水分吸收、运输、存储的主要功能组织[4-5],特别是根系作为贮藏营养物质和供给植株生长的重要器官[6-7],其生长状况直接影响苜蓿的生产性能及持久利用[8-9]。正是因为根系生长情况直接影响地上生物量,而根系形态结构又一定程度上决定了根系生长状况[10],所以通过探究苜蓿根系表型性状来剖析苜蓿的生产能力及越冬性具有重要意义。大量研究表明,根系形态可以影响苜蓿的生物固氮[8]、水分利用率[11]、再生特性[12]及抗寒性[13],尤其是对苜蓿能否多年可持续利用起关键作用。而在苜蓿根系形态结构中,靠近地面的根冠部为根颈,作为地上部分与根系的交接部位,其对低温具有较强的耐受性,这可能与品种的遗传特性及环境刺激等诸多因素有关[14],但是关于苜蓿根系性状对秋眠性及越冬抗寒性影响的报道却较少,以前有过一些研究和评估方法,如通过标准主轴分析(standardized major axis,SMA)法,解析不同秋眠级苜蓿根系性状在越冬过程中的差异化响应[3],采用偏最小二乘回归(partial least squares regression,PLSR)法,揭示根系性状对不同秋眠级苜蓿越冬率的影响程度及抗寒效应的关键因子[15]。鉴于目前仍不清楚苜蓿根颈、根系表型性状与秋眠性及越冬抗寒能力强弱的相互关系,本研究选取国内外不同秋眠级的苜蓿品种为试验材料,通过对苜蓿的地上、地下生物量、根系特征、越冬率及产量进行初步研究,力求探究苜蓿越冬耐寒性与根系特征和产量之间的关联,旨在为培育耐寒苜蓿品种提供优良种质资源,为苜蓿耐寒种质筛选提供参考与依据。
1 材料与方法
1.1 试验地概况
试验地位于黑龙江省齐齐哈尔市富拉尔基科研基地,E 123°24′36″,N 47°09′00″,最高温度38.5℃,最低温度-39.5℃,年均气温3℃,积温(≥10℃)为2722.1℃,年均降水量约480 mm,无霜期132 d左右。试验地土壤为黑风沙土,有机质含量19.89 g·kg-1,全氮1.19 g·kg-1,速效磷为10.60 mg·kg-1,速效钾为124.84 mg·kg-1,pH为7.4。
1.2 试验材料
选取国内外苜蓿材料8份,包括国内审定品种3份(草原3号、肇东、公农2号),国外引进品种5份(驯鹿、金皇后、420、三得利、赛迪),所有供试苜蓿材料的品种名称、秋眠级及来源信息见表1。

表1 供试苜蓿材料及来源Table 1 Source and materials of tested alfalfa
1.3 试验设计
试验小区面积为15 m2(3 m×5 m),2020年6月进行人工条播,行距30 cm,播种量15 kg·hm-2,采用完全随机区组设计,重复3次,每小区种植10行,各小区之间留60 cm间隔。播前精细整地,以确保种子发芽一致。试验期间实行常规田间管理,适时除草、病虫害防治,越冬前灌水1次,灌溉定额为225~375 m3·hm-2。建植当年9月下旬采用壕沟法,每小区随机选取30株重复,用铁锹挖取苜蓿根系,用自来水冲洗干净并吸干水分,带回实验室进行根系表型性状测量。2021年初花期(6月10日、7月20日、9月10日)进行3次刈割,留茬高度5 cm,测定产量。各项指标的具体测定方法如下:
采用根系分析系统(WinRHIZO)测量根系总长度、根颈体积、根尖数量等指标。
采用Marquez-Ortiz等[16]的方法测定根系形态性状。根颈直径:根颈膨大处直径采用游标卡尺测量;根颈入土深度:测量从地表到根颈上端距离;主根直径:根颈以下10 cm处主根直径;侧根数量:当距离主根0.5 cm处的侧根直径≥0.1 cm时,可计入侧根总数量,当直径<0.1 cm时,则不计入总数量;侧根直径:一级侧根距离主根2 cm处为侧根直径;侧根位置:地表与第一个侧根的距离;生物量:取30株单株苜蓿,分离茎秆(含叶片)和根系,在65℃烘箱内烘干24 h,称重后再烘6 h至恒重,即茎秆和根系干重。
产量:初花期测定每小区的鲜草产量,从刈割后苜蓿中称取1.0 kg,3次重复,65℃烘箱内烘干24 h,称重后再烘6 h至恒重,折算每hm2产量。
越冬率:每小区随机选取样段3处,样段长为1 m,越冬前及次年返青分别统计样段中植株总数,越冬率(%)=返青的植株数量/越冬前植株的总数量×100。
1.4 统计分析
采用Excel 2010进行数据处理,采用SAS 9.0软件进行方差分析、相关性分析和主成分分析。
应用模糊数学隶属函数法[17-18]对所测指标进行综合评价,求各项指标隶属函数值,若测定指标与苜蓿越冬耐寒性呈正相关,则

测定指标与越冬耐寒性呈负相关,则

式中:Fij为i草种j指标值;Xij为鉴定i草种j指标隶属值;Ximax为所鉴定i草种j指标最大值;Ximin为所鉴定i草种j指标最小值。将i草种各项指标函数值进行累加,求平均值,均值越大则苜蓿品种耐寒性越强。最后将8个苜蓿品种的耐寒性进行比较,得出品种间抗寒强弱顺序。
2 结果与分析
2.1 越冬率和根颈特征
由表2可知,不同品种之间越冬率存在显著差异,以品种公农2号、肇东和草原3号越冬率较高,分别为96.73%、95.48%、95.31%,显著高于其他品种(P<0.05);其次是驯鹿、金皇后、420和三得利苜蓿,分别是越冬率最低(赛迪)苜蓿的3.12、2.74、2.38、2.32倍。整体上看,秋眠等级越高的品种,其越冬率越低。根颈是苜蓿重要的贮藏器官,其直径大小是维持苜蓿抗寒能力的重要功能性状。随秋眠级的增加,根颈直径、体积、入土深度均呈降低的变化趋势。各品种根颈直径为4.75~5.20 mm;根颈入土深度为2.88~3.17 cm;根颈体积为0.265~0.299 cm3。不同品种间根颈直径、体积及入土深度差异较大,其中公农2号和肇东苜蓿的根颈直径、入土深度均显著高于其他品种(P<0.05);公农2号的根颈体积最大,肇东苜蓿(0.298 cm3)和草原3号(0.296 cm3)次之,赛迪苜蓿最小。
2.2 生物量
随秋眠级的增加,苜蓿建植当年地上生物量变化的总趋势表现为增高态势。由表2可知,品种金皇后、三得利、赛迪和420地上生物量显著高于公农2号、草原3号和肇东苜蓿(P<0.05),其中赛迪苜蓿地上生物量(37.59 g·plant-1)最高,尤其比地上生物量最低的公农2号高1.25倍。各品种的根系生物量为20.55~30.05 g·plant-1,以品种金皇后的根系生物量最高,其次为三得利、赛迪和420苜蓿,与公农2号相比分别增加了46.23%、42.19%、37.57%、25.01%。上述结果表明建植当年苜蓿地上生物量和根系生物量之间的关联程度很高,随秋眠等级增加,各品种的地上、根系生物量均表现为增加趋势,秋眠等级较高的品种地上生物量较高,则相应的地下生物量也较高。
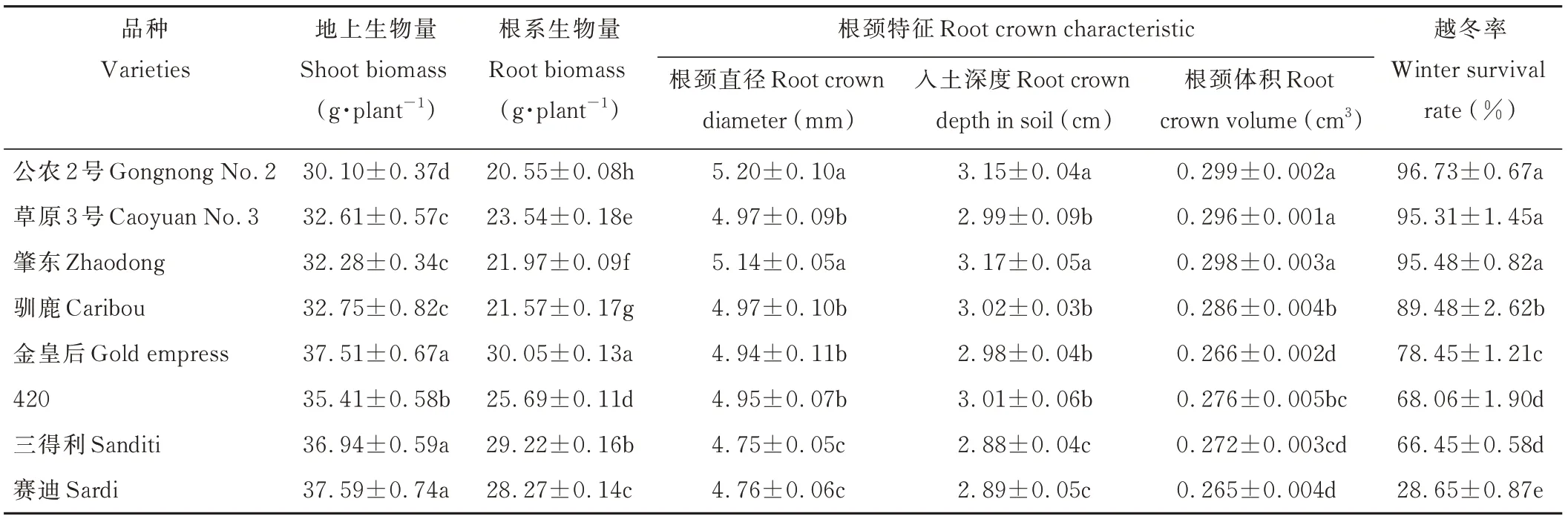
表2 不同苜蓿的地上、根系生物量、根颈特征和越冬率比较Table 2 Comparison of shoot biomass,root biomass,root crown characteristic and winter survival rate of different alfalfa
2.3 根系特征
苜蓿属于直根系,是由主根和侧根组成,在土壤状况良好或地下水位偏低的地区,根系深达2~3 m。由表3可知,不同品种间根系长度、主根直径和根尖数量差异较大,其中草原3号的根尖数量(2128.51)和根系长度(208.21 cm)最高,主根直径以品种金皇后、赛迪、420和三得利较高,分别为4.55、4.41、4.38、4.25 mm,显著高于其他品种(P<0.05)。各品种间侧根位置、侧根直径和侧根数量也存在显著差异,品种420、金皇后和赛迪的侧根位置显著高于其他品种(P<0.05);侧根直径以金皇后最大,比侧根直径最小的公农2号高25.35%;草原3号的侧根数量(3.20)最多,显著高于其他品种(P<0.05)。

表3 不同苜蓿的根系特征比较Table 3 Comparison of root characteristics of different alfalfa
2.4 产量
产量是苜蓿生产能力及耐寒适应性的综合体现,为探究建植次年苜蓿产量对越冬性的响应,测定2021年苜蓿总产量,由图1可知,各品种全年总产量为1465.28~12746.73 kg·hm-2,以肇东苜蓿产量最高,草原3号(11921.24 kg·hm-2)和公农2号(11416.77 kg·hm-2)次之,赛迪苜蓿最低;在饲草产量构成中,第1茬产量为808.25~6311.06 kg·hm-2;第2茬为470.91~4870.21 kg·hm-2;第3茬为186.11~1565.45 kg·hm-2,由此看出,第1茬产量明显高于第2和3茬。第1茬苜蓿产量占全年总产量的44%~56%;第2茬占32%~45%;第3茬占10%~13%。第1茬产量中,草原3号和肇东苜蓿显著高于其他品种(P<0.05),第2和3茬产量中,肇东苜蓿的产量均为最高,与其他品种相比差异显著(P<0.05)。该结果表明,品种产量高低可充分反映苜蓿耐寒性强弱,耐寒性强的品种越冬率较高,建植次年第1茬苜蓿产量较高,说明苜蓿产量与越冬率之间具有一定的相关性。

图1 不同苜蓿产量比较Fig.1 Comparison of yield of different alfalfa
2.5 综合评价分析
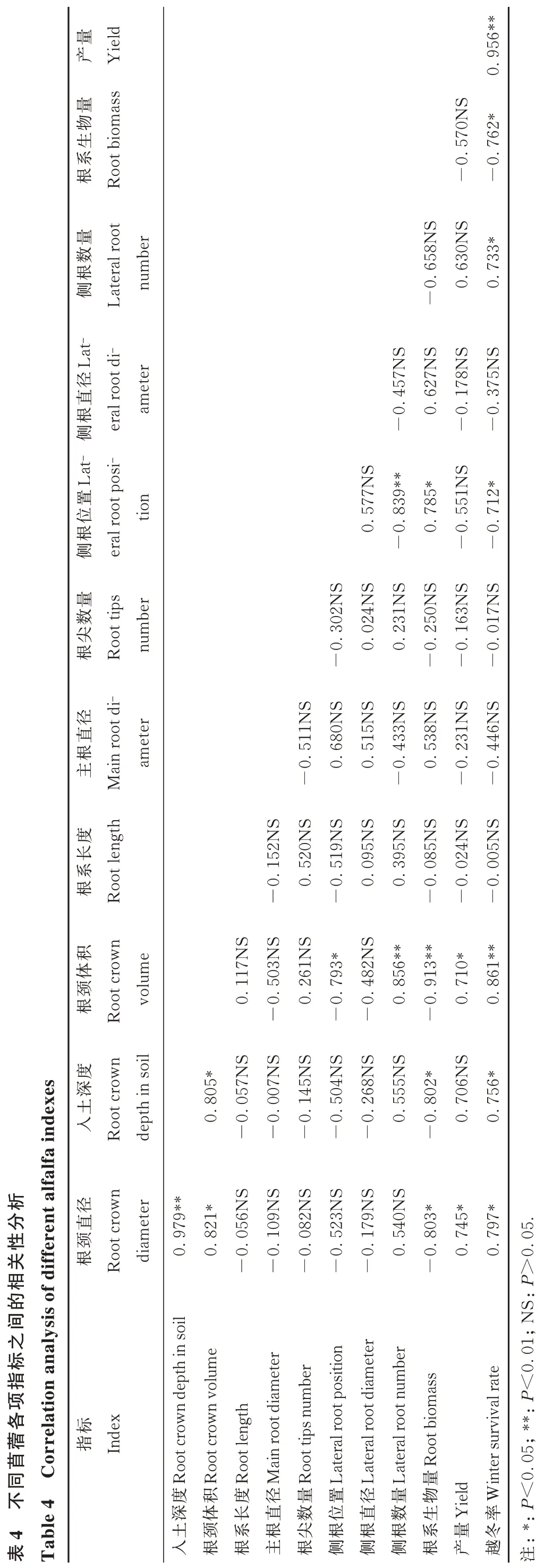
相关性分析结果表明,苜蓿的越冬率与根颈直径、入土深度之间存在显著正相关(P<0.05),相关系数为0.797、0.756,说明苜蓿根颈直径越大、入土越深,则苜蓿的越冬耐寒能力越强。苜蓿根颈直径与根颈体积、入土深度之间存在显著或极显著正相关,相关系数0.821和0.979,表明品种的根颈直径越大,则根颈入土越深。苜蓿越冬率与根系生物量呈显著负相关(-0.762;P<0.05),与产量呈极显著正相关(0.956;P<0.01),说明秋眠级越高的苜蓿品种地下生物量越大,其越冬能力越弱,则翌年越冬存活植株数越少、产量越低(表4)。
主成分分析是将12项指标转化成3个综合指标(F1、F2、F3)来弥补单一指标对苜蓿抗寒性评价的不足,根据累积贡献率≥85%原则,第一主成分(F1)特征值为7.019,贡献率58.50%,特征向量中数值较大的指标为根颈体积(0.368)、越冬率(0.331)和侧根数量(0.313);第二主成分(F2)特征值为2.146,贡献率17.89%,对应特征向量主要表现在根颈直径(0.321)、入土深度(0.349)和主根直径(0.386)。第三主成分(F3)特征值为1.223,贡献率10.19%,数值较大的指标是根系长度(0.522)和侧根直径(0.614)。上述3个主成分的累积贡献率达86.58%,基本包含所测指标的大部分信息(表5)。
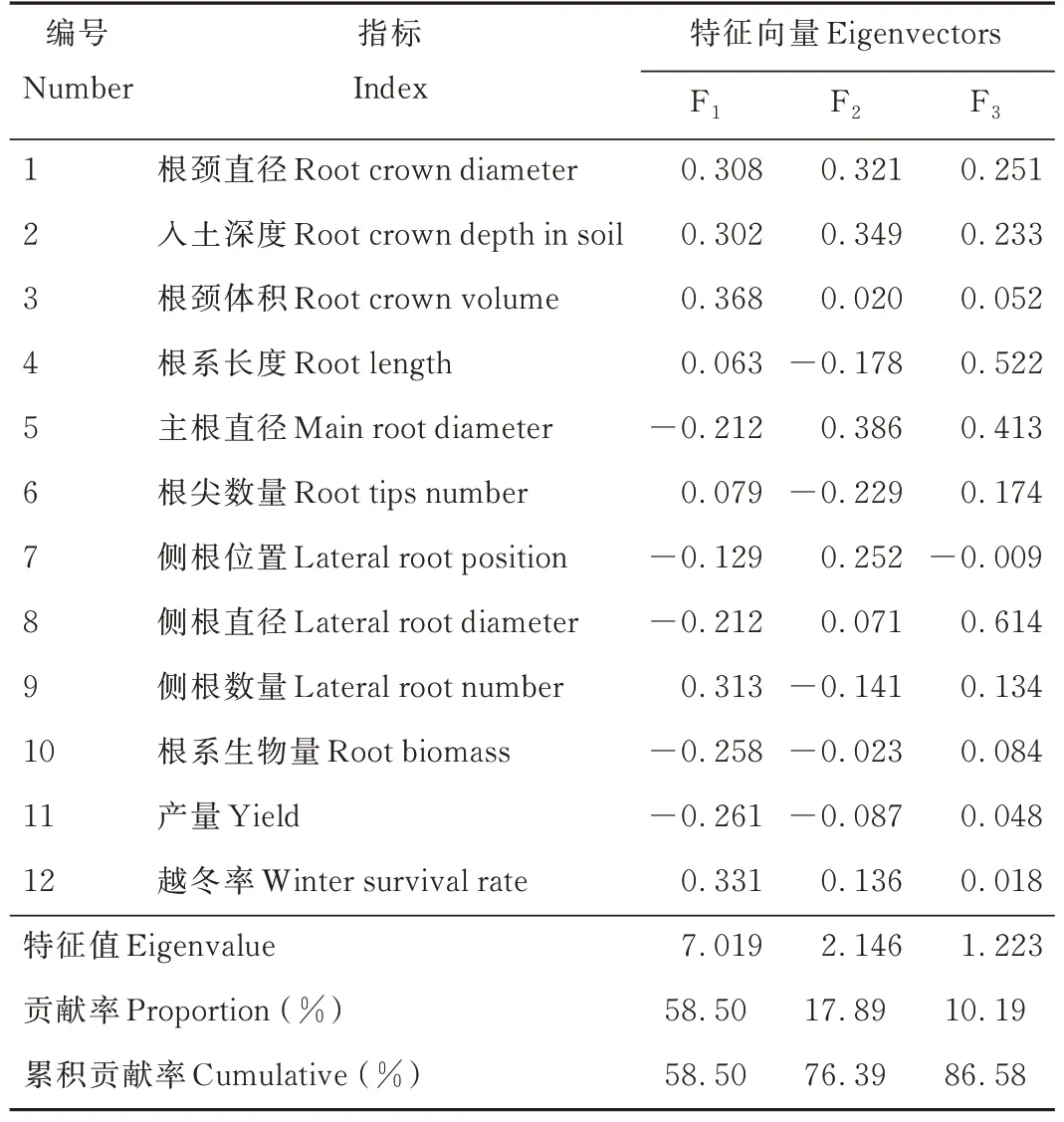
表5 主成分分析Table 5 Principal component analysis
通过对12项指标进行相关性及主成分分析,筛选出根颈直径、入土深度、根颈体积、根系长度、主根直径、侧根直径、侧根数量、越冬率作为评价耐寒种质材料的重要参考指标。采用隶属函数法对不同苜蓿品种的抗寒性进行综合评价,最终得出8个苜蓿品种的抗寒强弱顺序依次为:肇东(0.848)>公农2号(0.666)>草原3号(0.650)>金皇后(0.551)>驯鹿(0.505)>420(0.366)>三 得 利(0.293)>赛 迪 (0.268)(表6)。
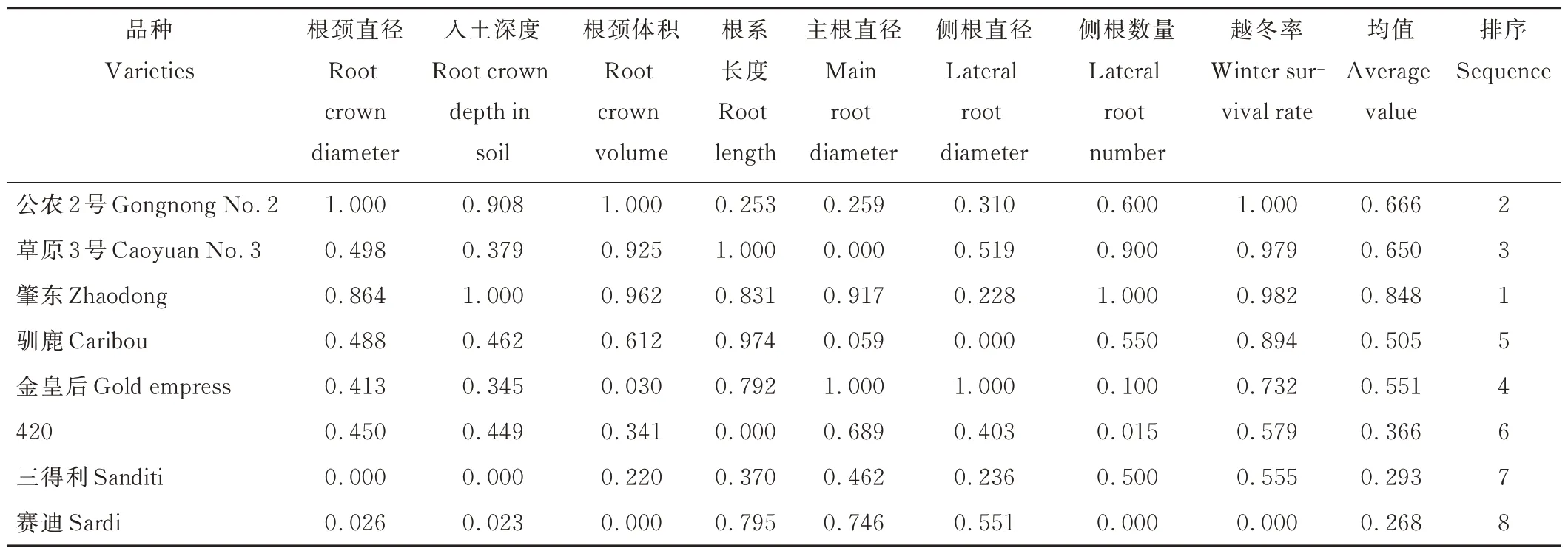
表6 不同苜蓿各项指标隶属函数分析Table 6 Analysis of the membership function of different alfalfa indexes
?
3 讨论
紫花苜蓿的抗寒性既受自身遗传因素影响,又与生长环境紧密相关。本研究发现,不同秋眠级苜蓿品种的越冬率存在明显差异,秋眠等级低的肇东、公农2号和草原3号越冬率均大于95%,而秋眠等级较高的品种如420、三得利、赛迪(fall dormancy 4~7)越冬率却较低,卢欣石[19]通过对国内审定苜蓿品种(23个)及地方品种(92个)的秋眠等级进行研究发现,苜蓿秋眠级别与其耐寒能力强弱有直接关系,这与Wang等[20]和Brummer等[21]研究得出的规律相一致,说明秋眠等级低的品种越冬能力强,更适应当地寒冷的气候环境[2],究其原因可能与苜蓿根系中积累了较多的棉子糖和氨基酸(如苏氨酸、组氨酸、脯氨酸和甘氨酸)有关[22]。尽管秋眠性与抗寒性在一定程度上存在关联,但二者并不属于同一性状,今后可以利用杂交育种方法将弱秋眠和强抗寒基因重新组合,进而选育出弱秋眠耐寒性强的新品种。
根系不仅具有养分和水分吸收、运输、贮藏功能,而且还是与越冬或抗寒性密切相关的主要功能器官[16]。在寒冬气候条件下,不同苜蓿根系性状表现出对冬季寒冷的不同响应,本研究所有供试品种中,主根直径对秋眠等级的响应尤为突出,随着秋眠级的升高,主根直径显著增大,这与徐大伟[23]的研究结果相一致。随着秋眠级升高,建植当年苜蓿根系生物量呈现逐步增高的变化趋势,且与越冬率呈较强的负相关,说明秋眠等级高的品种秋季根系贮藏的营养物质有向主根深处转移的倾向,致使主根过度消耗营养物质用于根系继续生长,导致根系总生物量增加,越冬率下降。侧根则呈现相反的规律,品种的侧根数量与越冬率呈显著正相关,这与Smith[24]的研究结果趋于一致,说明抗寒品种具有较多的侧根。同时Johnson等[25]研究发现,苜蓿品种由秋眠型至非秋眠型,侧根数量呈从多到少的变化趋势,原因在于侧根数量越多,则越有利于苜蓿从土壤中吸收较多水分和养分元素来供给越冬植株对营养物质的需求。另外,随侧根位置的加大即侧根距离地表越远,品种的越冬率呈下降趋势,可能由于越冬前地上茎叶组织的营养物质向地下深处转移,而转移的养分则在地下表现为逐级递减的规律,即侧根位置越靠下吸收的养分就越少,越冬率下降。本研究所用品种的根长、主根直径、根尖数、侧根直径与越冬率之间均未达到显著相关,说明根系表型特征并不是判定苜蓿抗寒能力强弱的关键因子,而抗寒性强弱还取决于根颈性状[26]。
根颈为连接植物茎部与根部的过渡结构,是连接地上部和地下部的关键部位,对低温的反应较为敏感,与苜蓿越冬及返青密切相关。本研究表明,随着秋眠级升高,根颈直径呈下降趋势,秋眠性较强的品种更倾向于增大根颈直径,这与刘志英[15]的研究结果基本一致,呈现这种变化规律可能与生长时期、遗传特性和环境因子的相互作用有关。相关性分析结果表明,根颈直径与越冬率呈现出很强的正相关,耐寒能力强的品种如肇东苜蓿、草原3号、公农2号根颈直径较大,可见根颈对低温有较强的适应性[27]。为适应寒冬,秋眠性较强的品种通过增大根颈直径来抵御低温,而秋眠性较弱甚至非秋眠型品种根颈直径增粗幅度较小,抗寒性较弱[23]。韩清芳等[28]研究发现,苜蓿的秋眠性还与根颈入土深度紧密相关,秋眠级越低的品种根颈入土深度越深,同时抗寒性与根颈入土深度也存在一定的关联,如抗寒性较强的公农2号和肇东苜蓿根颈入土较深,即抗寒品种根颈入土深度增加,越冬耐寒能力增强[4,28]。由此得出,苜蓿越冬率与根颈的大小及入土深度密切相关,根颈越大、根颈入土深度越深,则翌年苜蓿的越冬率越高。
产量是衡量苜蓿生产性能的重要指标,不同品种在同一生态区的产量存在明显差异[20],本研究中,品种肇东、草原3号和公农2号的产量明显高于其他品种,而秋眠等级为4~7级的420、三得利、赛迪苜蓿不仅越冬率较低,而且产量较低,表明品种的越冬率与秋眠性和产量之间存在很强的正相关,而秋眠等级为2~3级的驯鹿和金皇后苜蓿的产量亦较低,这与王晓龙等[2]和Malinowski等[29]的研究结果相一致,说明品种的秋眠等级与生产能力并无必然关联,究其原因可能与该地区气候、土壤及品种的遗传特性有关,因而秋眠等级并不是影响苜蓿产量的关键因子,而确定适合的种植(播种量、行距、播种深度)及收获(刈割时期、刈割高度)因子才是决定苜蓿高产的主要因素[29-31]。有研究指出,产草量因刈割茬次不同而差异明显,苜蓿第1茬产量对全年总产量贡献率大于第2和3茬[32]。本研究结果也得出同样规律,产量表现为第1茬>第2茬>第3茬,各茬苜蓿产量占全年总产量比例分别为50%、38%、12%,说明第1茬产量对苜蓿全年总产量十分重要,即使第1茬苜蓿的生长受到冬春季低温影响,但对全年总产量贡献仍最大,因此做好第1茬苜蓿田间管理是实现增产的关键。
4 结论
1)通过相关性及主成分分析确定根颈直径、入土深度、根颈体积、根系长度、主根直径、侧根直径、侧根数量、越冬率可作为评价耐寒种质材料的重要参考指标。
2)苜蓿品种由秋眠型、半秋眠型至非秋眠型,越冬率呈现逐步下降的变化;建植当年苜蓿主根直径和根系生物量呈现增高的变化;侧根数量呈现由多到少的变化趋势;根颈直径和根颈入土深度表现为下降的变化趋势。
3)采用模糊数学隶属函数法评价得出8个不同秋眠等级苜蓿抗寒性强弱顺序为:肇东>公农2号>草原3号>金皇后>驯鹿>420>三得利>赛迪。
4)综合比较得出公农2号、草原3号和肇东苜蓿的产量和越冬性均表现较好,较适宜黑龙江省西部地区种植推广。
