不同恢复年限草地群落水平植物功能性状及功能多样性分析
2023-02-07李江文何邦印李彩回虹燕刘博张晓曦樊慧苏文钰
李江文,何邦印,李彩,回虹燕,2,刘博,2,张晓曦,2,樊慧,苏文钰
(1.延安大学生命科学学院,陕西 延安 716100;2.陕西省红枣重点实验室,延安大学,陕西 延安 716100)
草地植物群落功能多样性反映了群落内物种间功能特征的数值和范围,与物种多样性不同,功能多样性考虑到植物群落中冗余种和种间互补作用[1]。目前,主要以植物功能性状表征或描述生态系统功能,将植物功能性状与生态系统功能联系起来,评价群落功能多样性变化规律[2]。群落加权平均性状(community weighted mean trait,CWM)是反映草地植物群落样本中期望函数值的度量,也被定义为植物性状的总价值,其变化来源包括群落物种组成变化和种内性状变异[3]。因此,综合考虑群落物种组成与群落水平植物功能性状对草地生态系统功能多样性的影响,从不同层次考虑群落物种多样性及功能多样性的变化,成为研究草地植物群落功能变化的新途径。
物种组成是确定草地植物群落类型的依据,决定了植物群落的性质和功能。已有研究表明,随着恢复年限的延长,草地群落结构和功能将产生负面效应,草地群落物种多样性有下降的趋势[4]。长期封育后,可能会改变灌木与草本的种间关系,导致草地出现灌丛化现象[5]。灌丛化显著改变草地植物群落物种组成、多样性及生产力,进而对草地植物群落结构、功能及其稳定性产生影响[6]。而灌丛化的不断扩增降低了群落中草本植物分布的均匀性,对草本植物多样性产生负面影响[7]。草地植物群落物种之间以及物种与生境之间关系复杂,这些关系所反映的结果可以通过植物功能性状的变化来体现[8]。将群落物种组成与群落水平功能性状变化相结合,分析草地植物群落功能多样性动态变化,对于研究草地生态系统功能变化具有重要意义[9]。
黄土高原因人类过度利用与开发,造成其水土流失严重,生态环境极其脆弱,其也是我国最早实施退耕还林、还草工程的地区。为恢复退化草地,黄土高原地区广泛实施了多种封育措施,形成了以实施退耕还林、还草为主的封育区和以农田撂荒地为主的撂荒草地。基于此,本研究以黄土高原丘陵沟壑区不同恢复年限人工刺槐林(Robinia pseudoacacia)林下草地和自然撂荒草地作为研究对象。通过对不同恢复年限两种草地类型草本植物群落进行调查,并结合植物功能性状指标,研究不同恢复年限两种草地类型植物群落物种组成、群落加权平均性状、物种多样性及功能多样性的动态变化规律,以期揭示不同恢复年限黄土高原丘陵沟壑区林下草地与撂荒草地群落水平植物功能性状及功能多样性动态变化,为黄土高原丘陵沟壑区草地生态系统功能评估提供理论依据。
1 材料与方法
1.1 研究区概况
本研究采样区位于陕西省安塞区纸坊沟流域(109°14′36″-109°16′03″E,36°42′42″-36°46′28″N),该区域属于黄土高原丘陵沟壑区第二副区,面积约为8.27 km2。属暖温带半干旱气候,年平均气温8.8℃,年平均降水量483 mm,降水主要集中在6-9月,约占全年总降水量的70%以上。流域内沟壑密集,地面起伏较大,海拔1010~1431 m。土壤类型以黄绵土为主。植被类型处于暖温带阔叶林向干草原过渡的森林草原带,草地植被主要以黄刺玫(Rosa xanthina)、铁杆蒿(Artemisia sacrorum)、狗尾草(Setaria viridis)、猪毛蒿(Artemisia scoparia)、野菊(Chrysanthemum indicum)、披碱草(Elymus dahuricus)等为主。
1.2 研究方法
2021年8月,在研究区选取10、20、30和45 a(不同恢复年限,表1)立地条件基本一致的林下草地和撂荒草地,其中林下草地人工林品种为刺槐。在不同恢复年限的林下草地和撂荒草地分别设置3个重复样地(32 m×32 m),共计24个(林下草地12个、撂荒草地12个),每个重复样地设置4个1 m×1 m固定样方,用于调查草地植物群落物种组成及植物功能性状。同时,采集每个固定样方中所有植物的叶片,要求采集叶片新鲜、完整、无病虫害,用于测量植物叶片面积及叶片质量。

表1 研究地基本情况Table 1 The basic situation of the research site
1.3 数据统计与分析
2020年8月,基于样方法调查固定样方内群落物种组成,测量样方内所有物种盖度、高度、密度及生物量,并归类群落物种生活型、纲及所属科。采集固定样方内所有物种完整叶片,每个物种采集6株,每株取叶片12~20个,使用叶面积仪(LI-3000C,美国)测定所有植物叶片面积,取平均值为该种植物叶面积大小。待测完植物叶面积后,将对应叶片分类装进信封袋内并带回实验室,置于65℃烘箱中烘干至恒重,使用万分之一电子天平(BSA224S,中国)称取单株采集叶片干重,取平均值即为叶片质量。
本研究选用α物种多样性指数反映群落内物种组织化水平,具体选用指数及计算方法参考如下:物种丰富度指数(species richness index)、Shannon-Wiener指数[10]、均匀度指数(evenness index)[11]和优势度指数(dominance index)[12]。
使用F-Diversity软件计算功能多样性指数[13-15],F-Diversity软件构建了一个方便使用的界面,以开放的源代码例程来评估和分析功能多样性指数,开放源代码的计算平台是统计软件R[16]。本研究采用以下4个多样性指标:Rao二次熵指数(Rao’s quadratic entropy)、功能丰富度指数(functional richness index)、功能均匀度指数(functional evenness index)和离散度指数(functional divergence index),具体计算公式及方法参照Mason等[17]。
本研究使用群落加权平均性状反映草地群落水平植物功能性状的变化规律。群落加权平均性状(CWM)由每个物种的特征代表值(平均值、中位值和模式)和相对丰度计算所得。计算公式如下:
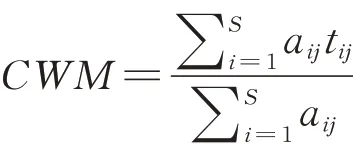
式中:S为群落j中的物种数目,aij表示物种i在群落j中的相对多度,tij表示物种i在群落j中的性状值。
利用SPSS 16.0统计软件分析不同恢复年限和不同草地类型群落加权平均性状、物种多样性和功能多样性之间的差异性,显著性检验水平为0.05,使用SigmaPlot 12.5对分析结果构图。
2 结果与分析
2.1 不同恢复年限草地植物群落物种组成动态分析
调查不同恢复时期刺槐林林下草地与撂荒草地草本植物组成,依据分属科排序可知(图1),林下草地共发现草本植物43种,分属18科36属,撂荒草地发现草本植物38种,分属16科35属。茜草科、葡萄科、毛茛科、车前科、伞形科和蝶形花科仅出现在林下草地,百合科、大戟科、远志科和堇菜科仅出现在撂荒草地。林下草地与撂荒草地植物群落物种主要以菊科、禾本科和蔷薇科为主。其中10~30 a林下草地物种以菊科为主,分别占比24%、44%和34%,45 a林下草地物种以禾本科为主,达26%;10~45 a撂荒草地物种以菊科为主,依次占比35%、33%、29%和40%。10~45 a,林下草地以菊科为主的植物群落逐渐演替到以禾本科为主的植物群落,而撂荒草地物种始终以菊科为主要物种。

图1 不同恢复年限草地植物分属科排序Fig.1 Ordination of grassland plant families in different restoration years
依据植物生活型差异分析不同恢复年限草地植物群落物种组成(图2A,B),林下草地与撂荒草地物种均以多年生杂类草为主,数量随恢复年限的延长呈增加-降低-增加的变化趋势,且以20 a物种数量最多,占比分别达30%和38%。不同恢复年限灌木和半灌木与多年生杂类草数量动态变化规律正好相反,且以20 a物种数量最少,林下草地灌木和半灌木占比仅10%,而撂荒草地未出现灌木与半灌木物种,但随着恢复年限的延长,撂荒草地灌木和半灌木比例增加。不同恢复年限林下草地一、二年生草本和多年生禾草数目动态变化幅度较小,但撂荒草地一、二年生草本植物数量随恢复年限的延长呈先降低后增加的变化趋势(转折点为30 a),多年生禾草的数目呈先增加后降低的变化趋势(转折点为20 a),且动态变化幅度较大。依据植物所属纲排序分析群落物种组成(图2C,D),随着恢复期的延长,林下草地双子叶植物数目逐渐减少,单子叶植物数目逐渐增加,而撂荒草地双子叶植物和单子叶植物数目呈先增加后降低的变化趋势,且20 a物种平均数量最多,分别占比43%和29%。
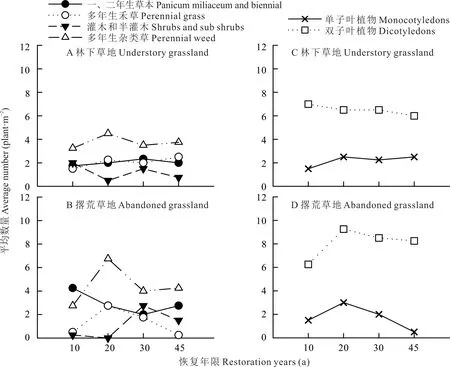
图2 不同恢复年限草地植物生活型、纲排序动态变化Fig.2 The dynamic changes of plant life forms and class ordination in different restoration years
2.2 草地群落水平植物功能性状差异性分析
群落加权平均性状差异性分析结果显示(表2),45 a撂荒草地的CWM盖度、CWM密度和CWM地上生物量最高,且CWM盖度显著高于20 a(P<0.05),CWM密度显著高于20和30 a(P<0.05),CWM地上生物量显著高于10和20 a(P<0.05)。30 a林下草地CWM地上生物量最高,且显著高于20 a(P<0.05)。10 a林下草地CWM叶面积显著高于其他年份(P<0.05),10 a林下草地CWM叶片质量显著高于20 a(P<0.05)。30 a撂荒草地CWM叶面积和CWM叶片质量最高,且显著高于10和45 a(P<0.05)。林下草地与撂荒草地群落加权平均性状差异性分析结果显示,林下草地与撂荒草地之间CWM高度无显著差异。10 a林下草地CWM密度显著低于撂荒草地(P<0.05),但林下草地CWM叶面积和CWM叶片质量显著高于撂荒草地(P<0.05)。45 a林下草地CWM盖度、CWM密度和CWM地上生物量显著低于撂荒草地(P<0.05),而林下草地CWM叶面积和CWM叶片质量显著高于撂荒草地(P<0.05)。
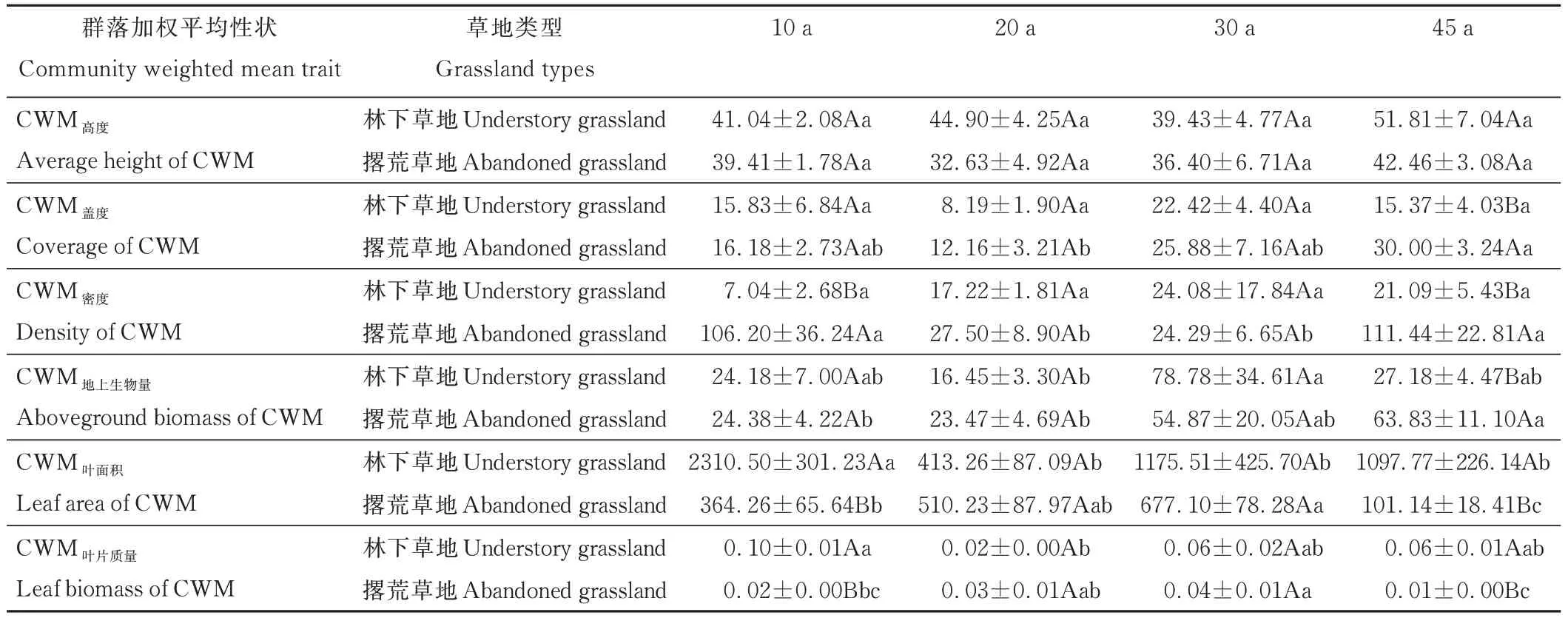
表2 不同恢复年限草地植物群落加权平均性状的差异性Table 2 Difference in community weighted mean trait(CWM)of grassland plant communities in different restoration years
2.3 草地群落物种多样性及功能多样性差异性分析
不同恢复年限草地植物群落物种多样性差异性分析结果显示(图3),20 a撂荒草地物种丰富度指数和Shannon-Wiener指数最高,分别为12.25和2.32,且显著高于10和45 a(P<0.05),撂荒草地20和30 a物种优势度指数显著高于10 a(P<0.05)。而不同恢复年限林下草地物种多样性指数(物种丰富度、Shannon-Wiener指数、均匀度指数和优势度指数)无显著性差异,且林下草地与撂荒草地之间均匀度指数也无显著性差异(P>0.05)。20和30 a撂荒草地物种丰富度指数显著高于林下草地(P<0.05),20 a撂荒草地Shannon-Wiener指数和物种优势度指数显著高于林下草地(P<0.05)。整体而言,林下草地物种多样性低于撂荒草地。双因素方差分析结果显示,恢复年限对物种丰富度指数产生显著影响(P<0.05),恢复类型对香农维纳指数产生显著影响(P<0.05),此外,恢复年限、恢复类型及两者交互效应对均匀度指数和优势度指数均无显著影响(P>0.05)。
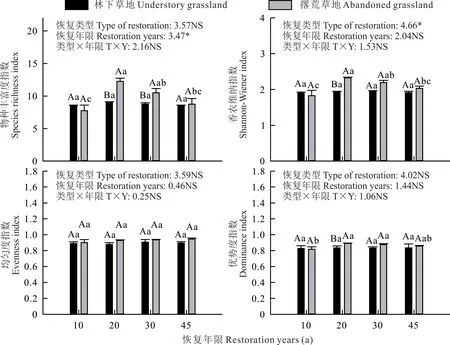
图3 不同恢复年限草地植物群落物种多样性的差异性Fig.3 Differences in species diversity of grassland plant communities in different restoration years
植物群落功能多样性差异性分析如图4所示,20 a撂荒草地的Rao二次熵指数和功能丰富度指数最高,分别为0.50和0.002,且20和30 a撂荒草地的Rao二次熵指数显著高于10 a(P<0.05),20 a撂荒草地功能丰富度指数显著高于10和45 a(P<0.05)。而不同恢复年限之间林下草地植物群落功能多样性指数均无显著性差异(P>0.05)。不同恢复年限撂荒草地功能均匀度指数和功能离散度指数无显著性差异(P>0.05)。10 a林下草地Rao二次熵指数显著高于撂荒草地(P<0.05),20 a林下草地Rao二次熵指数和功能丰富度指数显著低于撂荒草地(P<0.05)。整体而言,林下草地功能多样性低于撂荒草地,林下草地较撂荒草地生态效益更低。双因素方差分析结果显示,恢复年限、恢复类型及两者交互效应对功能丰富度指数产生显著影响(P<0.05),而对Rao二次熵指数、功能均匀度指数和功能离散度指数均无显著影响(P>0.05)。

图4 不同恢复年限草地植物群落功能多样性的差异性Fig.4 Differences in functional diversity of grassland plant communities in different restoration years
3 讨论
3.1 不同恢复年限草地植物群落物种组成
本研究选取不同恢复年限刺槐林林下草地与撂荒草地,研究黄土高原丘陵沟壑区草地植物群落功能多样性动态变化,具有典型代表性。刺槐作为陕北地区退耕还林主要建植树种,在黄土高原地区分布广泛,而撂荒草地是退牧还草、农田撂荒后形成的黄土高原区又一典型草地类型。因形成背景不同,林下草地与撂荒草地的生境不同,造成两种草地类型群落物种组成差异较大。本研究分析指出,随恢复年限增加,林下草地从以双子叶为主的菊科逐渐演替到以单子叶为主的禾本科,而撂荒草地始终以菊科物种占比最高。主要原因在于随着刺槐林龄的增加,刺槐林的覆盖度和生物量随之增加,林下光照资源减少、刺槐生态位扩大,导致林下部分耐旱性较差、资源获取能力较低的植物优势地位逐渐衰退,而喜荫、耐旱性较强的物种入侵并逐渐成为优势种[18-19],水分和光照成为影响林下群落物种组成变化的关键因素[20]。以人工方式建植的刺槐林,可以很快地改变土壤理化性质和林下小气候,从而为林下草地植被提供新的生存环境和资源[21]。林下草地双子叶植物数目随着恢复年限的延长减少,单子叶植物数目增加的主因,正是耐受性较强物种逐渐取代耐受性较低物种的体现。而撂荒草地光照、水分等因素不受人工林干扰,不同恢复年限之间非生物环境与林下草地比较变化较小,对优势物种的影响需要更长的时间跨度,因此本研究结论指出群落优势物种仍以菊科为主。
不同功能属性物种对环境的适应性存在较大差异,造成不同恢复年限草地不同功能属性物种为适应特定生存环境,数量发生变化[22]。林下草地与撂荒草地物种以多年生杂类草为主,体现了多年生杂类草具有较强的适应性。土壤质地、水分、透气性等为不同植物提供了生长和繁殖的条件[23],而多年生禾草的数量随恢复年限的延长呈增加-降低-增加的趋势,且以20 a多年生杂类草的数量最多,灌木和半灌木数量最少,主要受土壤条件、环境因子、种内斗争和种间竞争的共同影响[24]。同样,坡向、海拔等立地条件不同也可能导致20 a草地物种数量变化,需进一步研究讨论。不同恢复年限多年生杂类草与灌木和半灌木数量动态变化规律正好相反,撂荒草地多年生禾草与双子叶植物、单子叶植物同步增加后再降低,灌木与半灌木数量随恢复年限延长整体呈增加的趋势,都体现了群落在恢复过程中的灌丛化趋势[25]。
3.2 草地群落水平植物功能性状动态变化
群落水平植物功能性状是将植物个体水平功能性状与物种相对多度相结合,用于描述群落对环境变化的响应规律[26]。群落加权平均性状能够对环境过滤作用作出响应,可以与环境变化、生态系统服务相联系,影响生态系统功能[27-29]。本研究指出,林下草地10 a植物叶片性状的群落加权平均值最大,而30 a地上生物量的群落加权平均值最大。群落水平植物功能性状因环境因素不同会产生较大的差异[3],10 a刺槐林覆盖度较低,造就了林下植物群落叶片性状的群落加权平均值较大,而30 a林下草地地上生物量、撂荒草地叶面积及叶片质量的群落加权平均值最大,则可能归因于土壤水分含量的突增及采样地坡向位置,需结合不同坡向及海拔等其他干扰因素进一步讨论研究。草地群落加权平均性状的变化一方面是种内性状变异导致,另一方面还在于群落物种组成及群落动态变化发挥作用[30]。随着恢复年限增加,撂荒草地地上生物量的群落加权平均性状显著增加,不同物种个体生物量存在较大差异,而群落物种组成的改变将导致地上生物量的加权平均性状发生改变。45 a撂荒草地盖度、密度和地上生物量的群落加权平均性状显著大于林下草地,这与群落物种多样性的变化规律一致,因此群落物种组成可能是影响两种草地类型盖度、密度和地上生物量的群落加权平均性状出现显著性差异的主因。而10和45 a林下草地叶面积和叶片质量的群落加权平均性状显著大于撂荒草地,可能主要由种内性状差异导致。
3.3 草地植物群落物种多样性与功能多样性动态变化
林下草地与撂荒草地从根本上讲,分属于森林生态系统和草地生态系统[31],这意味着将原有的农田生态系统转换为能够自我更新的草地生态系统,必然引起地上与地下生物环境与非生物环境的不同[32-33]。不同恢复年限之间林下草地物种多样性与功能多样性并无显著变化,一方面由于林下草地双子叶植物数量降低后,单子叶植物数量增加;另一方面在于林下生境未影响群落水平高度和密度功能性状的显著变化,其他性状的改变未对功能多样性指数变化产生较大影响。而20 a撂荒草地物种多样性(丰富度指数、Shannon-Wiener指数和优势度指数)、功能多样性(Rao二次熵指数和功能丰富度指数)显著增加,与此同时,20 a撂荒草地多年生植物平均数量显著增多,而一、二年生草本植物数量明显下降。这可能是由于水分亏缺缓解了群落内的种间竞争,提高了非优势种的丰富度和生物量,从而在短期内增加了物种多样性[34]。
本研究指出,物种丰富度与功能丰富度指数、Rao二次熵与Shannon-Wiener指数、物种均匀度与功能均匀度指数变化同步,这与前期研究物种多样性与功能多样性的关系主要呈现正相关结果一致[1]。物种多样性与功能离散度指数之间变化规律并无显著相关性也有一定的依据[35],由于物种多样性可能主要是由资源可利用程度决定,而功能多样性可能是由于具有相似功能性状的物种能够通过环境的筛选,引起种间竞争等种间作用产生生态位分化所致,因此造成物种多样性与功能离散度之间无相关性[36]。撂荒草地物种多样性及功能多样性高于林下草地,一方面在于林下草地除草本植物外,刺槐可拦截降水、削弱降水侵蚀,进而促进水源涵养,增加物种生存空间,提升草地植物群落地上净初级生产力,从而影响植物群落物种组成及功能变化[37-38]。另一方面,撂荒草地相较于人工林恢复地,在较长的时间能够获得更大的生态效益[39],因此其多样性指标要高于林下草地。另外,30 a撂荒草地物种多样性和功能多样性处于较高水平,可能原因在于30 a草地高生物量物种比例增加(如灌木与半灌木),导致群落功能多样性改变。
4 结论
黄土高原不同恢复年限草地植物群落主要以多年生菊科和蔷薇科为主。随着恢复年限增加,林下草地植物从以双子叶为主逐渐演替到以单子叶为主,从以菊科为主逐渐演替到以禾本科为主。而撂荒草地物种始终以菊科物种为主,在恢复过程中逐渐演替为以灌木和半灌木为主的群落,且双子叶植物和单子叶植物同步增加后再降低。
林下草地群落水平叶片功能性状高于撂荒草地,而群落水平盖度、密度和地上生物量性状低于撂荒草地。林下草地物种多样性和功能多样性指数低于撂荒草地,撂荒草地相较于林下草地具有更高的生态效益。
