褐飞虱神经肽及其受体基因的功能筛查
2023-01-19王斯亮罗序梅张传溪
王斯亮,罗序梅,张传溪,3*
(1.浙江大学农业与生物技术学院昆虫科学研究所,杭州 310058;2.温州科技职业学院农业与生物技术学院,浙江 温州 325006;3.宁波大学植物病毒学研究所,农产品质量安全危害因子与风险防控国家重点实验室/农业农村部和浙江省植物保护生物技术重点实验室,浙江 宁波315211)
神经肽广泛存在于低等和高等生物中,作为激素、神经递质、神经调质发挥生理功能。褐飞虱(Nilaparvata lugens)是我国南方稻区的首要害虫,主要吸食水稻叶鞘韧皮部的汁液,田间褐飞虱种群的为害造成“虱烧”现象,严重时会引起大面积水稻枯秆倒伏[1]。对于害虫的绿色防治,利用天敌、病原微生物以及特异性环境友好型农药是重要的方法。得益于褐飞虱基因组和转录组数据的解析以及褐飞虱系统性RNA干扰操作体系的建立[2-3],褐飞虱的功能基因组的研究正在逐步开展,其中,分析鉴定得到的褐飞虱重要生理功能基因,具有成为防治褐飞虱的特异性农药靶标的潜力。神经肽在昆虫中承担着调控多种重要生理功能的作用,包括蜕皮、取食、求偶、繁殖、形体表观发育等[4-5]。一些神经肽(如影响蜕皮的神经肽)的功能缺失或失调会造成昆虫重要的形态或生理缺陷,或使昆虫死亡。
随着生物信息学的发展,越来越多的昆虫神经肽通过组学方法被识别。TANAKA等[6]通过对褐飞虱转录组的分析,搜索鉴别到一系列神经肽以及相应的G蛋白偶联受体的同源基因序列。与此相同,本研究对褐飞虱神经肽及其受体基因进行RNA 干扰,对神经肽生理功能进行筛查验证,期望得到具有敲减后致死致畸表型的神经肽基因,以及对尚未明确功能的神经肽进行功能分析。本文也概览了褐飞虱神经肽的生理功能,筛选出对褐飞虱存活率和发育有明显影响的神经肽及其受体基因,为害虫的绿色防控提供了新的潜在靶标。
1 材料与方法
1.1 供试虫源
褐飞虱试虫于2008 年采集自浙江大学华家池校区(30°16′35.21″ N,120°11′37.16″ E),传代饲养至今。用于饲养褐飞虱的水稻品种为‘秀水134’。饲养条件为:温度(25±1)℃,相对湿度60%~70%,光照周期16 h/8 h(光照/黑暗)。
1.2 序列查找
利用TANAKA 等[6]报道的褐飞虱神经肽及其受体基因cDNA 序列,通过Blast 2.2.29+程序,在褐飞虱转录组和基因组中搜索比对,并搜索可变剪接本的存在,通过聚合酶链反应(polymerase chain reaction,PCR)和克隆测序验证,得到用于设计双链RNA(double-stranded RNA, dsRNA)的褐飞虱神经肽及其受体基因的模板序列。
1.3 RNA 干扰(RNA interference,RNAi)
在特异性引物的5′端添加T7 启动子,利用MEGAscript T7 转 录 试 剂 盒(MEGAscript T7 Transcription Kit,美国Ambion 公司)合成目标基因的dsRNA(根据不同基因的开放阅读框长度,dsRNA 长度范围设计为200~500 bp),用无核酸酶的水将dsRNA溶液稀释为5 μg/μL。注射绿色荧光蛋白(green fluorescent protein,GFP)基因的dsRNA(dsGFP)组作为阴性对照。使用FemtoJet显微注射仪(德国Eppendorf公司)将dsRNA注射到褐飞虱体内,每组选取50~100 头若虫进行注射,3 龄若虫每头注射20 nL,5 龄若虫每头注射100 nL。干扰后2~3 d取虫样提取总RNA,采用定量PCR验证干扰效果。为了排除实验中不同批次的水稻苗的营养条件差异造成的背景干扰,本研究采用“成虫相对存活率”来代表实验组中褐飞虱成虫的存活率,其中,成虫相对存活率=实验组成虫存活率/对照组成虫存活率。
1.4 总RNA 提取与反转录
采用Trizol 法提取褐飞虱的总RNA。利用cDNA 第一链反转录试剂盒[HiScriptⅡ1st Strand cDNA Synthesis Kit(+gDNA wiper)](南京诺唯赞生物科技股份有限公司)合成cDNA第一链,反应体系为20 μL,加入1 μg总RNA进行反应。
1.5 荧光定量PCR 分析
用荧光定量PCR试剂盒(SYBR Green PCR Kit,美国Bio-Rad公司)制备荧光定量PCR反应体系,在Bio-Rad CFX-96实时荧光定量PCR仪中反应。内参基因为褐飞虱18S rRNA[7](Nl18S,GenBank登录号为JN662398.1),正向引物为5′-CGCTACTACCGA TTGAA-3′,反向引物为5′-GGAAACCTTGTTACG ACTT-3′。荧光定量PCR 反应程序:95 ℃预变性5 min;95 ℃变性10 s,60 ℃延伸30 s,40 个循环。每个基因进行3次生物学重复和3次技术重复。利用相对定量法2-ΔΔCT[8]计算各个基因的相对表达水平。
2 结果与分析
2.1 神经肽及其受体基因的功能筛查
通过PCR 扩增并验证得到了41 个神经肽基因(含1个可变剪接本)和44个受体基因。以褐飞虱3龄若虫代表低龄若虫,5龄代表高龄若虫,利用RNA干扰法沉默目标神经肽基因,随后观察褐飞虱表型。将阴性对照dsGFP注射入3龄若虫后,褐飞虱培养至成虫的存活率在0.6左右,而在注射5龄若虫后,培养至成虫的存活率在0.9左右。结果(表1)显示:在褐飞虱3 龄时进行注射,RNA 干扰后成虫相对存活率<0.2的有4个神经肽基因,即NlCCAP[编码甲壳类动心肽(crustacean cardioactive peptide,CCAP)]、NlETH[编码蜕皮诱导激素(ecdysis triggering hormone,ETH)]、NlOKA[编码螯虾激肽(orcokinin,OK)]和NlPK[编码焦谷氨酸激肽(pyrokinin,PK)]。其余基因沉默后,成虫相对存活率都在0.6以上。在褐飞虱5 龄时进行注射,对所有基因进行RNA 干扰后成虫相对存活率>0.5。NlCCAP和NlETH基因的沉默对低龄若虫具有剧烈的致死效果,但对高龄若虫的致死效果不明显。在低龄若虫中进行RNA 干扰造成高死亡率的基因有NlPK和NlOKA,而在高龄若虫中RNA 干扰此类基因却不具有致死效果,成虫相对存活率>0.9。

表1 褐飞虱神经肽基因的RNA干扰效果Table 1 RNAi effects of neuropeptide genes in N.lugens
通过对神经肽基因功能的筛查验证,我们发现4 种神经肽基因NlETH、NlCCAP、NlPK、NlOKA经RNA干扰后可造成蜕皮阶段致死的表型。Nlburs-a和Nlburs-b(分别编码鞣化激素的2个亚基)受到干扰后,褐飞虱成虫无法展翅。神经肽基因Nlelevenin受到干扰后,褐飞虱体壁颜色黑化。在3 龄若虫阶段,上述这些神经肽基因沉默后产生明显的表型变化,而在5 龄若虫阶段受到干扰后表型变化不显著。
对受体基因进行RNA干扰发现,当干扰3龄若虫时,仅NlA36(PK 受体基因)和NlA46(鞣化激素受体基因)经干扰后成虫相对存活率<0.2,其余均大于0.5。5 龄若虫中NlA46经干扰后成虫仍有较高的死亡率,其相对存活率约为0.3(表2)。对NlA42与Nlelevenin进行干扰后产生的褐飞虱表型变化一致,均为体壁黑化[9]。

表2 褐飞虱神经肽受体基因的RNA干扰效果Table 2 RNAi effects of neuropeptide receptor genes in N.lugens

表2(续) Continuation of Table 2
针对受RNA 干扰后产生明显外观变化的神经肽 及 其 受 体 基 因Nlburs-a、Nlburs-b、NlCCAP、NlETH、Nlelevenin、NlA36、NlA42和NlA46,我们分别设计了与原干扰片段不重叠的另一段干扰片段,通过观察干扰后产生的表型是否一致来验证是否存在脱靶效应。结果显示,所有基因在新双链RNA的干扰下产生的表型与原双链RNA的一致,且干扰效率均达到65%以上。
2.2 经典蜕皮相关神经肽在褐飞虱中的表现
2.2.1 蜕皮诱导激素(ETH)
对褐飞虱3龄若虫进行RNA干扰,发现NlETH沉默后,若虫主要在4 龄末和5 龄末蜕皮前死亡。对4 龄若虫注射dsNlETH,蜕皮前死亡的现象主要出现在5龄末;对5龄若虫进行干扰,褐飞虱成虫只有约一半表现为蜕皮前死亡(图1)。

图1 褐飞虱在NlETH基因经RNA干扰后的存活率和表型Fig.1 Survival rates and phenotypes of N.lugens with NlETH knockdown after RNAi
2.2.2 甲壳类动心肽(CCAP)
本研究对褐飞虱3龄若虫注射dsNlCCAP后,发现其在4 龄至5 龄蜕皮以及5 龄至成虫蜕皮期间死亡,蜕裂线没有完全打开,整个虫体无法从旧皮中脱出(图2)。与NlETH相似,NlCCAP在褐飞虱中的沉默表型也表现为跨龄期的模式。对4龄若虫进行干扰,蜕皮受阻的表型主要出现在5 龄蜕皮至成虫阶段;对5龄若虫进行干扰,则大部分褐飞虱能正常蜕皮羽化为成虫。
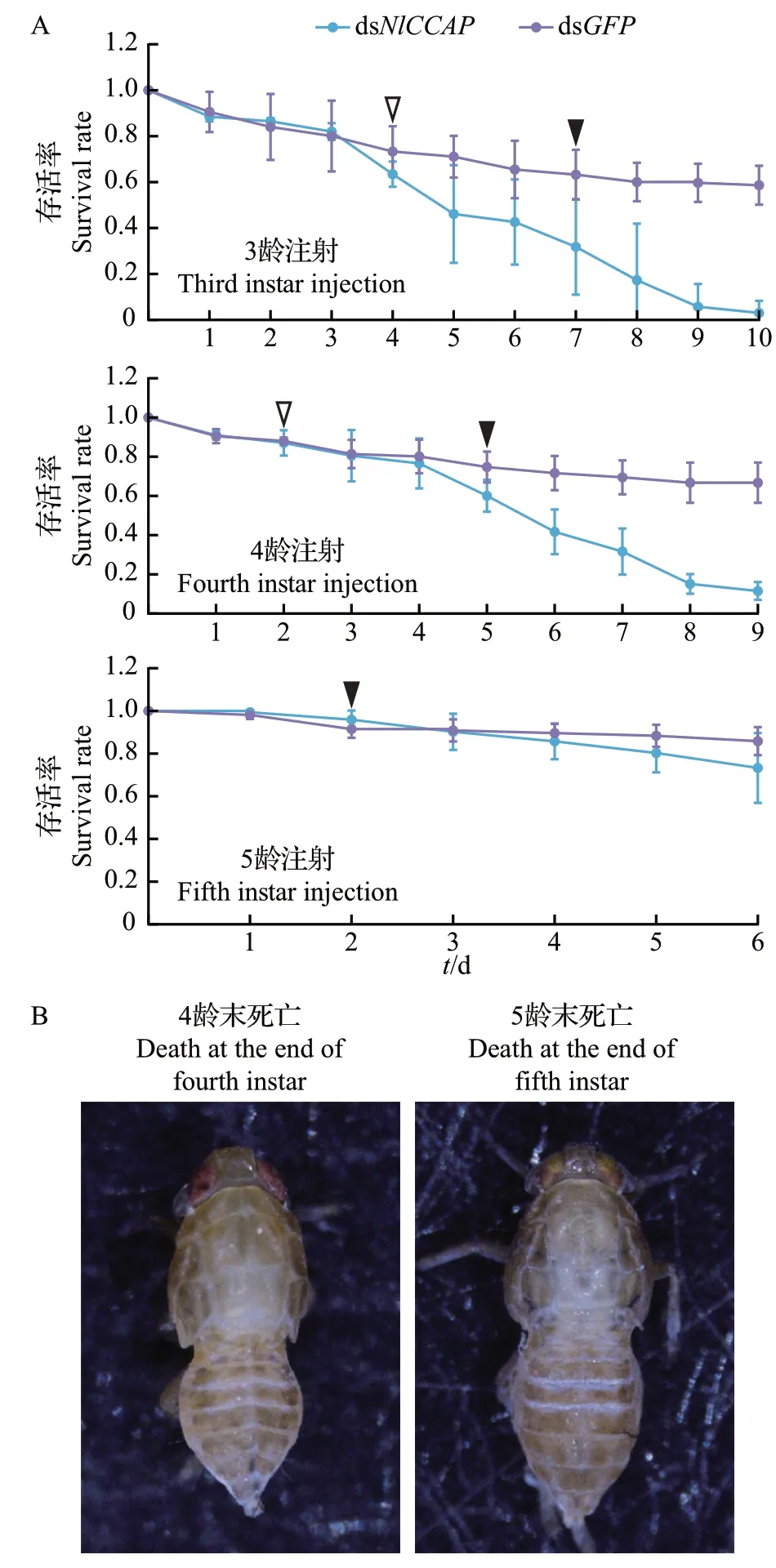
图2 褐飞虱在NlCCAP 基因经RNA 干扰后的存活率和表型Fig.2 Survival rates and phenotypes of N. lugens with NlCCAP knockdown after RNAi
2.2.3 鞣化激素
鞣化激素是由2个不同基因编码的亚基组成的神经肽类激素。在褐飞虱中,编码鞣化激素2 个亚基的基因分别为Nlburs-a和Nlburs-b。
在针对编码鞣化激素基因的RNA 干扰实验中,对Nlburs-a及Nlburs-b进行独立干扰时不影响另一个亚基的表达量(图3)。注射dsNlburs-a和dsNlburs-b产生的表型变化一致,均是造成成虫展翅不正常(图4)。在不同龄期干扰鞣化激素基因,均对褐飞虱的存活率没有显著影响,但产生的表型有所不同。在3龄若虫时期进行RNA干扰,褐飞虱发育至成虫时翅发育严重畸形,表现为无法正常伸展;在4龄若虫时期进行干扰,成虫的翅仍无法正常伸展,但翅展面积比3龄若虫时期干扰时稍大;而在5 龄若虫时期进行干扰,成虫翅面能伸展至透明状态,但仍有部分弯曲。随着注射的虫龄增加,干扰后产生的卷翅成虫的比例也越少(图5)。编码鞣化激素的基因经干扰后,褐飞虱表型呈现越低龄期进行干扰,受到的影响越严重。对鞣化激素的受体基因NlA46进行干扰,结果(图6)显示产生的外观表型与Nlburs-a和Nlburs-b沉默的效应一致,但不同的是成虫的死亡率较高,此时大部分成虫蜕皮后表皮柔软,鞣化过程明显受阻,最终死亡。该结果揭示了神经肽受体基因经RNA 干扰后产生的效应比配体基因受干扰后更剧烈。
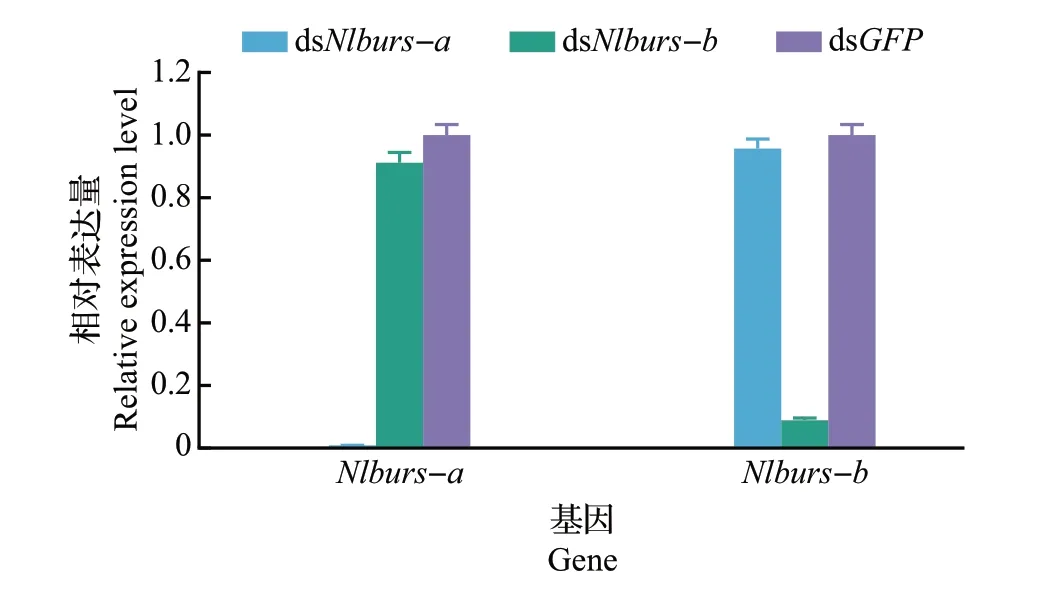
图3 注射不同的dsRNA 后编码鞣化激素的2 个亚基的基因表达量变化Fig.3 Expression level changes of genes encoding two subunits of bursicon after injected with different dsRNAs
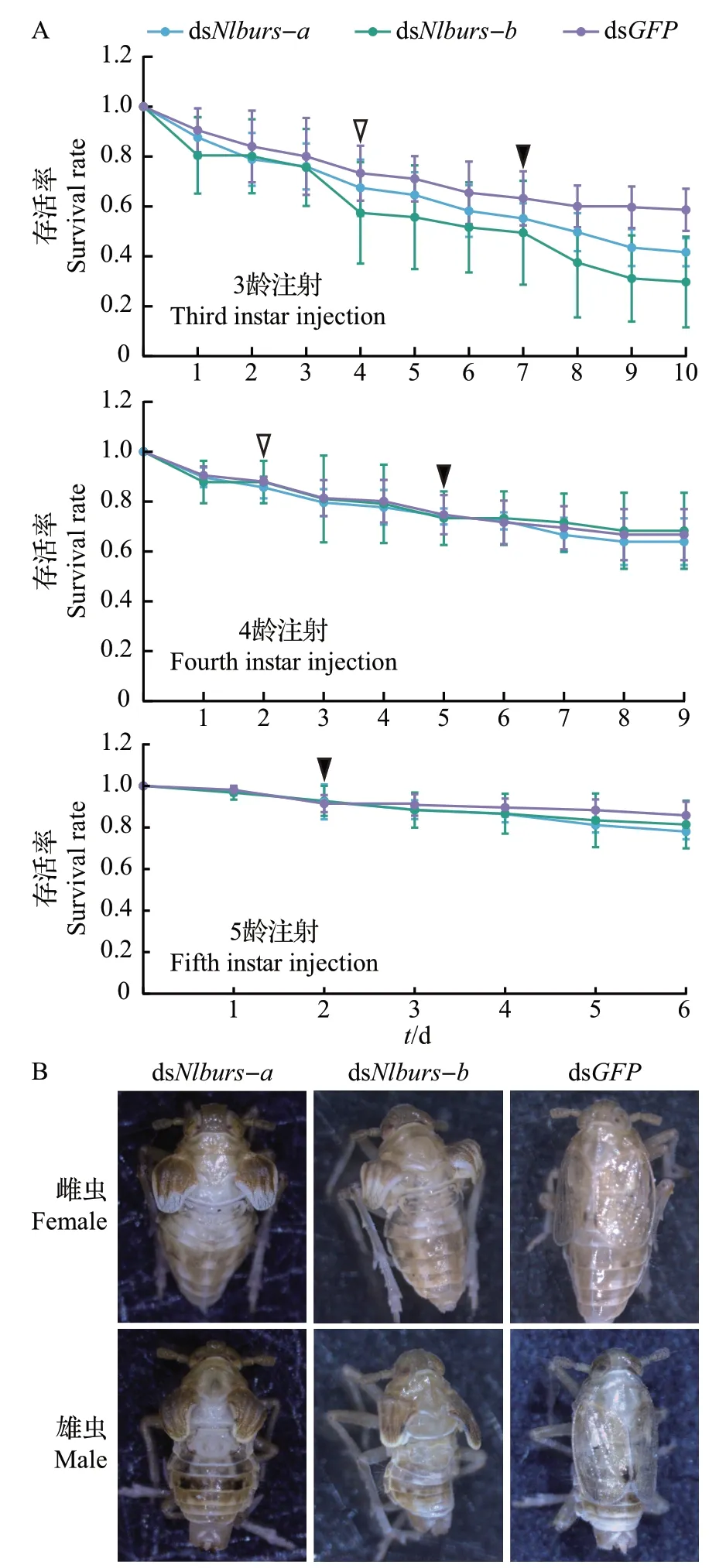
图4 褐飞虱在Nlburs-a和Nlburs-b基因经RNA干扰后的存活率和表型Fig.4 Survival rates and phenotypes of N. lugens with Nlburs-a and Nlburs-b knockdown after RNAi

图5 褐飞虱不同龄期干扰鞣化激素基因对成虫表型(A)及成虫卷翅率(B)的影响Fig.5 Effects of bursicon knockdown on phenotypes(A)and wing-unstretched rates of adults(B)in different instars of N.lugens

图6 褐飞虱在NlA46基因经RNA干扰后的存活率和表型Fig.6 Survival rates and phenotypes of N.lugens with NlA46 knockdown after RNAi
3 讨论
RNA 干扰应用于害虫防治是一种可行的策略[10]。本研究通过RNA 干扰方法对褐飞虱神经肽及其受体基因的生理功能进行筛查,以成虫相对存活率=0.5为界,发现在干扰3龄若虫时,在神经肽基因中,仅NlCCAP、NlETH、NlOKA和NlPK在褐飞虱3 龄阶段沉默后可造成存活率显著降低的现象;受体基因中NlA36(PK 受体基因)和NlA46(鞣化激素受体基因)受干扰后成虫相对存活率<0.5。然而CCAP的受体基因NlA27以及ETH的受体基因NlA6沉默后没有产生明显的表型变化。而这些在低龄若虫中被干扰产生致死现象的基因,在高龄(5 龄)若虫中被干扰时,除了NlA46外其余均没有产生明显的致死效果。
蜕皮是包括昆虫在内的节肢动物的重要生理过程。由于表皮以外骨骼的形式存在,个体的生长会受到阻碍,因此就需要在一定阶段脱去旧的表皮,使旧表皮内已经生长的躯体得到释放并形成新的表皮。前人的研究表明,蜕皮受多种激素、神经肽调控,包括蜕皮诱导激素(ETH)、甲壳类动心肽(CCAP)、促前胸腺激素(prothoracicotropic hormone, PTTH)、羽化激素(eclosion hormone, EH)等[11]。鞣化激素在昆虫中具有调控蜕皮后表皮鞣化的重要作用[12-13]。在赤拟谷盗(Tribolium castaneum)中,研究人员对ETH、EH、CCAP、鞣化激素相应的基因进行了干扰,发现它们在蜕皮的不同阶段(蜕皮前、蜕皮时、蜕皮后)中都扮演着重要的角色[14]。而在本研究中,NlCCAP和NlETH基因沉默后,褐飞虱表现出蜕皮受阻或蜕皮前死亡的现象,Nlburs-a、Nlburs-b与NlA46基因沉默后,褐飞虱表现出表皮柔软、无法正常展翅的现象。这些基因与在其他昆虫中报道的功能相似。而PTTH、EH等在其他昆虫中表现为对蜕皮有调控作用的神经肽,其在褐飞虱中的相应同源基因受干扰后没有产生明显的表型。另外几种褐飞虱中干扰后造成蜕皮受阻现象的基因为NlOKA(编码OK)、NlPK(编码PK)以及PK 的受体基因NlA36。OK 在家蚕中可以刺激前胸腺的活性,被认为可能是蜕皮激素的上游调控因子[15]。在长红猎蝽(Rhodnius prolixus)中,编码OK 的基因受干扰后也会造成蜕皮受阻[16]。而PK 的功能仍有很多未解之处,且在不同昆虫中功能差异很大,如在马德拉蜚蠊(Leucophaea maderae)中可引起后肠收缩[17],在家蚕(Bombyx mori)中参与飞行肌的运动调控[18],在麻蝇(Neobellieria bullata)中促进化蛹[19],在甲虫中调控脏肌收缩和血糖水平[20]。在本研究中,首次发现PK在褐飞虱中与蜕皮有关,不过有关褐飞虱OK和PK的研究有待深入开展。
4 结语
本研究通过RNA 干扰对褐飞虱神经肽及其受体基因进行了初步功能筛查,发现4 个神经肽基因(NlCCAP、NlETH、NlOKA和NlPK)和2 个受体基因(NlA36和NlA46)对褐飞虱的存活至关重要,具有成为害虫防治靶标的潜力,并初步概览了褐飞虱神经肽的生理功能。对RNA 干扰后未产生明显表型变化的神经肽及其受体基因,后续可以对其进行“组合干扰”[21],观察这些基因之间是否存在功能互补等,以进一步理解褐飞虱神经肽/受体信号的调控网络。
