水稻粒型与粒质量的QTL分析
2016-07-25谢婷婷张令瑄郭伟伟赵金凤李学勇
谢婷婷+张令瑄+郭伟伟+赵金凤+李学勇+张文会


摘要:以元江普通野生稻与优良栽培稻亲本特青配制的野生稻染色体片段代换系为材料,对水稻粒长、粒宽、粒厚、容积、密度、粒质量进行数量性状位点(QTL)定位。结果分别检测到12个与粒长相关的QTLs,16个与粒宽相关的QTLs,9个与粒厚相关的QTLs,3个与密度有关的QTLs,14个与籽粒容积有关的QTLs,10个与粒质量相关的QTLs,其中17个QTLs位点被多次重复检测到。相关系数分析及QTLs位点分析证明,容积、粒厚对粒质量的贡献率最大,且粒型间也有一定的相关性,尤其是粒厚与容积的相关性最大,同时也说明粒厚与容积可能有相同的遗传基础。此外,有研究表明,野生稻逐渐进化成的栽培稻更有利于提高产量。
关键词:水稻;粒型;粒质量;数量性状位点(QTL)
中图分类号: S511.032文献标志码: A文章编号:1002-1302(2016)06-0099-05
收稿日期:2015-05-17
基金项目:山东省自然科学基金(编号:ZR2014CL009)。
作者简介:谢婷婷(1989—),女,山东聊城人,硕士研究生,主要从事分子复制遗传育种研究。Tel:(0635)8230714;E-mail:xietingting.aa@163.com。
通信作者:张文会,博士,教授,从事植物生理胁迫的研究。Tel:(0635)8230714;E-mail:whzhang@lcu.edu.cn。据报道,2030年世界水稻总产量必须是当前产量的140%才能满足人口对粮食中稻谷的需求[1]。粒质量是构成产量的3个要素之一,对提高水稻产量具有重要意义[2-4]。而粒质量是一个与籽粒长度、宽度、厚度有关的综合指标,因而改良粒长、粒宽、粒厚等粒型对提高水稻产量也具有十分重要的意义。在2000年的国际水稻遗传大会上,学者提出了较为权威的稻属分类系统,将水稻分为23个种,包括2个栽培种、21个野生稻种[5]。目前公认的亚洲栽培稻起源于普通野生稻,中国栽培稻通常被认为起源于中国普通野生稻[6]。
至2013年12月,在水稻中共定位到102个影响粒长的数量性状位点(QTL)、73个影响粒宽的QTLs(http://www.Gramene.org),遍布于水稻的12条染色体上[7]。随着水稻高通量遗传图谱的构建和全基因组序列的公布,一些水稻粒型基因相继被精细定位和克隆。已经克隆到的与粒宽相关的基因包括:Song等发现的GW2[8],Shomura等克隆的5号染色体上的控制粒宽的主效基因qSW5[9],Weng等克隆的GW5[10],Li等克隆的位于GW5附近的控制粒宽的微效QTL GS5[11],Wang等克隆的位于8号染色体上的控制粒宽的主效QTL GW8[12],以及Fan等克隆的GS3[3],Qi等克隆的控制粒长、粒质量的GL3.1[13],Ishimaru等克隆的控制水稻粒质量的基因TGW6[14]。
本研究从粒型的6个指标(粒长、粒宽、粒厚、籽粒容积、籽粒密度、粒质量)入手,分析其各自的遗传规律,检测各自的QTL位点,并对彼此之间进行相关系数及直接通径系数分析,重点分析粒长、粒宽、粒厚、籽粒容积、籽粒密度分别对粒质量的贡献率,以期能为水稻粒型QTL/基因克隆及水稻的高产育种提供理论依据。
1材料与方法
1.1试验材料
供试材料为云南元江普通野生稻与优良籼稻品种特青(TQ)配制的高代回交渗入系群体,由中国农业大学孙传清博士提供。
1.2试验方法
1.2.1遗传图谱构建根据Temnykh等发表的水稻SSR序列[15]合成引物。用在亲本间有多态性的112个SSR标记,调查上述群体106个系的基因型。
1.2.2DNA提取及PCR扩增采用CTAB 法(略有改动)提取水稻基因组DNA[16],具体操作步骤如下:取每个株系的适量叶片置于液氮中快速研磨后移入1.5 mL离心管中,加 700 μL 1.5%CTAB,65 ℃温浴30 min后加600 μL三氯甲烷/异戊醇(24 ∶ 1),颠倒混匀,12 000 r/min离心10 min;转移上清液至另一管中,向其中加入2/3体积的预冷异丙醇,置于-20 ℃ 冰箱中30 min以上,12 000 r/min离心10 min;弃上清,用体积分数为70%的乙醇洗涤沉淀2次,吹干,加200 μL TE缓冲液溶解,-40 ℃冻存,在PCR分析时作为模板使用。
PCR扩增采用天根生化科技(北京)有限公司的普通Taq DNA聚合酶,PCR反应体系为:1 μL 10×Taq buffer,0.1 μL dNTP (2.5 mmol/L),0.4 μL Primer (F+R) (10 μmol/L),1 μL DNA,0.1 μL Taq,加ddH2O至10 μL。PCR反应程序为:94 ℃ 5 min;94 ℃ 30 s,53~65 ℃ 30 s,72 ℃ 30 s,35个循环;72 ℃ 5 min,扩增产物经8%非变性聚丙烯酰胺凝胶电泳分离,继而进行银染显影并作统计分析。
1.2.3数据测量测定性状包括稻米的粒长、粒宽、粒厚、容积、密度、粒质量。采用《水稻种质资源描述规范和数据标准》鉴定群体与亲本的粒型性状。随机抽取85株作为分析群体,每株随机抽取成熟稻米5粒,用游标卡尺测量粒长、粒宽、粒厚,并取其平均值。按如下公式计算每粒稻米的容积并取其平均值:
稻米容积=粒长×粒宽×粒厚。
粒质量是利用电子天平逐个称量,并取其平均值。按如下公式计算每粒稻米的密度并取其平均值:
稻米密度=粒质量/每粒米容积。
用软件DPS7.0计算粒长、粒宽、粒厚、容积、密度、粒质量的相关系数与直接通径系数。
1.2.4QTL分析采用Map Manager QTX b17软件,以概率值小于0.05作为判断QTL存在的阈值,采用单标记分析法进行定位分析。参考Temnykh等报道的水稻SSR连锁图谱为框架[15],构建连锁图谱,112个SSR标记分布于12条染色体上,各染色体上最多的有13个标记,最少的有7个标记,平均每条染色体上有9.3个标记,标记平均间距17.8 cM,基本能满足QTL定位的需要。
2结果与分析
2.1性状分析
粒长、粒宽、粒厚、容积、密度、粒质量的数据分布情况见图1,可见6组数据均呈正态分布,说明所得数据适于进行QTL分析。
2.2相关性分析
粒长、粒宽、粒厚、容积与粒质量的相关性均达到显著水平,由大到小依次为籽粒容积、粒厚、粒宽、粒长,分别为0.919、0.661、0.530、0.302。粒长、粒宽、粒厚、容积与密度之间也有一定的相关性,除粒长与容积、密度与容积之间的相关性不显著外,其他粒型之间均极显著相关(表1)。此外,各粒型性状对粒质量的直接通径系数显示,粒容积对粒质量的直接通径系数最大,为0.413 9;粒厚次之,为0133 3(表1)。可以看出,容积与粒质量的相关系数以及直接通径系数均是最大的,其次为粒厚,说明籽粒容积对粒质量的贡献率最大,粒厚次之。
2.3QTL分析
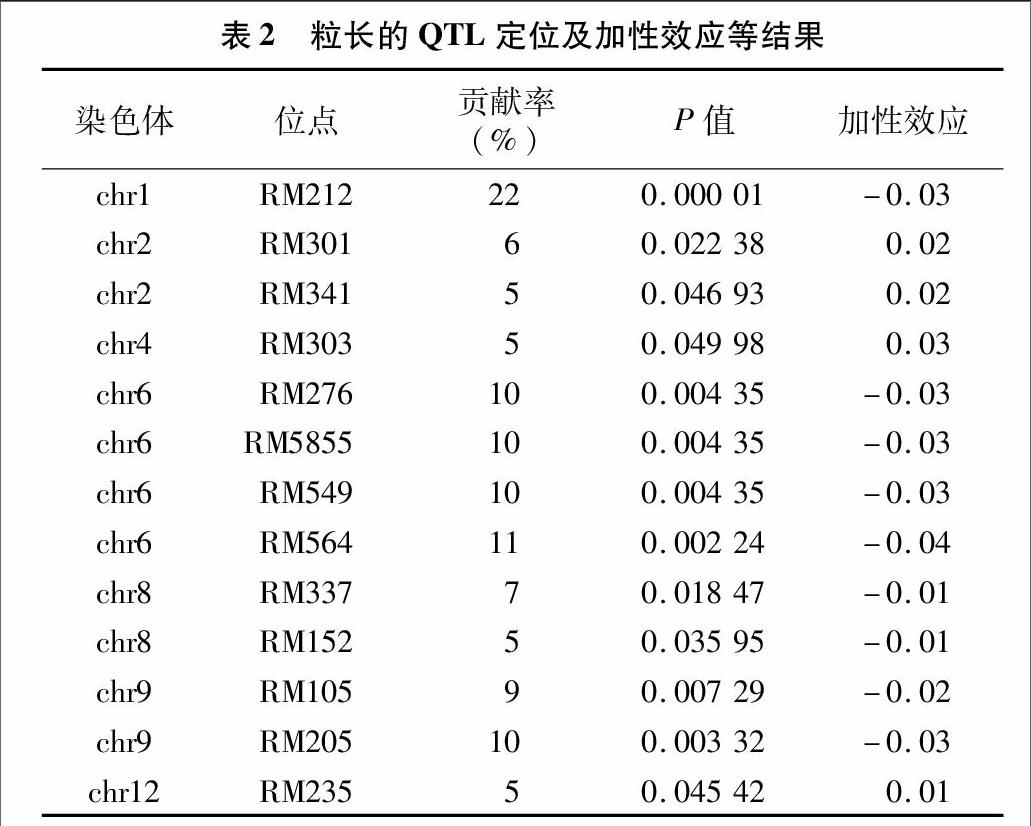
2.3.1粒长以粒长作为检测标准,共检测到13个与粒长相关的QTLs(表2),分别位于第1、2、4、6、8、9、12号染色体上。其中,2号染色体上的RM301、RM341,4号染色体上的RM303,12号染色体上的RM235附近的QTL位点贡献率分别为6%、5%、5%、5%,加性效应分别为0.02、0.02、0.03、001,显示来源于TQ的等位基因QTL位点促进粒长变长。位于1号染色体上的RM212,6号染色体上的RM276、RM5855、RM549、RM564,8号染色体上的RM337、RM152,9号染色体上的RM105、RM205附近QTL位点的贡献率分别为22%、10%、10%、10%、11%、7%、5%、9%、10%,加性效应分别为-0.03、-0.03、-0.03、-0.03、-0.04、-0.01、-001、-0.02、-0.03,表明源于野生稻的等位基因QTL位点促进粒长变长,尤其是1号染色体的RM212的贡献率、加性效应均较大,分析其为主效QTL。
2.3.2粒宽以粒宽作为检测标准,检测到16个与粒宽相关的QTLs(表3),分别位于第1、5、7、8、9、12号染色体上。其中,1号染色体上的RM1201、RM1183、RM212、RM6489,5号染色体上的RM4777、RM1089、RM7118,7号染色体上的RM481、RM82、RM82,8号染色体上的RM152、RM339,9号染色体上RM201、RM3787、RM1099的贡献率分别为22%、7%、17%、18%、7%、17%、18%、5%、11%、8%、7%、6%、22%、29%、14%,加性效应分别为0.02、0.01、0.01、0.02、0.01、001、0.01、0.01、0.01、0.01、0.01、0.01、0.01、0.02、0.01,表明来源于TQ的等位基因QTL位点促进粒变宽。12号染色体上的RM235的贡献率为8%,加性效应为-0.01,表明来源于野生稻的等位基因QTL位点促进粒变宽。其中1号染色体的RM1201、9号染色体的RM3787附近QTL位点的贡献率、加性效应均较大,分析其为主效QTL。
2.3.3粒厚以粒厚为检测标准,共检测到9个与粒厚相关的QTL位点(表4),分别位于第2、3、5、7、9号染色体上。其中,3号染色体上的RM6676,5号染色体上的RM1089、RM7118,7号染色体上的ID41、RM5711、RM82、RM125,9号染色体上的RM3787的贡献率分别为10%、14%、14%、7%、11%、18%、16%、7%,加性效应分别为0.01、0.01、0.01、001、0.01、0.01、0.01、0.01,表明来源于TQ的等位基因QTL位点促进粒厚增加。2号染色体上的RM301附近的QTL位点的贡献率为5%,加性效应为-0.01,表明来源于野生稻的等位基因的QTL位点表现为促进粒变厚。可以看出,加性效应大小均为0.01,然而贡献率最大的为7号染色体的RM82,分析其为主效QTL。
2.3.4籽粒密度以粒密度为检测标准,检测到3个与密度有关的QTLs(表5),分别位于第7、9号染色体上。7号染色体上的RM5344,9号染色体上的RM3787、RM1099附近的QTL位点的贡献率分别为7%、6%、5%,加性效应分别为002、001、0.01,表明来自TQ的等位基因的QTL表现为促进籽粒密度变大。其中7号染色体上的RM5344附近QTL位点的贡献率和加性效应均为最大,分析其为主效QTL。
2.3.5籽粒容积以籽粒容积作为检测标准,共检测到14个与籽粒容积有关的QTLs(表6),分别位于第2、3、5、7号染色体上。2号染色体上的RM7451、RM7033、RM5984、RM233、RM6,3号染色体上的RM6676,5号染色体上的RM4777、RM1089、RM7118,7号染色体上的ID41、RM481、RM5711、RM82、RM125附近的QTL位点的贡献率均为0.01,表明来源于TQ的等位基因QTL位点促进籽粒容积变大。其中贡献率最大的为7号染色体上RM82附近的QTL位点,分析其为主效QTL。
2.3.6粒质量以粒质量为检测标准,共检测到10个与粒质量相关的QTLs(表7),分别位于第2、5、7号染色体上。2号染色体上的RM5984、RM6,5号染色体上的RM4777、RM1089、RM7118,7号染色体上的ID41、RM481、RM5711、RM82、RM125附近的QTL位点的贡献率分别为8%、6%、6%、15%、16%、6%、11%、8%、17%、13%,加性效应均为0.01,表明来源于TQ的等位基因QTL位点促进粒质量提高。其中贡献率最大的为7号染色体上的RM82,分析其为主效基因。
2.3.7重复检测到的QTL位点表8结果表明,有多个QTL位点被多次检测到,被重复检测到的QTL位点分布于第1、2、3、5、7、8、9、12号染色体上。5号染色体上的RM1089、RM7118附近的QTL位点,7号染色体上的RM82、RM125附近的QTL位点被重复检测到4次,7号染色体上的ID41、RM481、RM5711,9号染色体上RM3787附近的QTL位点被检测到3次,其余的被检测到2次。初步分析认为,这些QTLs是稳定的QTLs。此外,从表8中也不难看出:籽粒容积与粒质量的QTL位点重叠率最高,即以粒质量为限制因素被重复检测到的QTL位点在籽粒容积下全部被检测到。这一结果与前面提到的相关系数的分析结果相呼应,籽粒容积对粒质量的贡献最大。
3讨论
3.1QTL位点分析
林鸿宣等检测到了5个控制粒长的微效QTLs、9个控制粒宽的QTLs[17]。Redona等定位到7个控制粒长的QTLs、4个控制粒宽的QTLs[18]。邢永忠等定位到8个控制粒长、5个控制粒宽的QTLs[19]。Rabiei等定位到5个影响粒长、7个影响粒宽的QTLs[20]。Govindaraj等定位到1个控制粒长、1个控制粒宽的QTLs[21]。赵芳明等检测到2个粒长QTLs、2个粒宽QTLs[22]。Amarawathi等定位到3个影响粒长的QTLs、2个影响粒宽的QTLs[23]。Hu等定位到7个影响粒长的QTLs、6个影响粒宽的QTLs[24]。
在本研究中,共检测到12个与粒长有关的QTLs,其中1号染色体的RM212贡献率最大,分析其为主效QTL。检测到16个与粒宽相关的QTLs,其中1号染色体的RM1201、9号染色体的RM3787附近QTL位点的贡献率、加性效应均较大,分析其为主效QTL。检测到9个与粒厚相关的QTL位点,7号染色体的RM82贡献率最大,分析其为主效QTL。检测到3个与密度有关的QTLs。检测到14个与籽粒容积有关的QTLs,其中7号染色体上RM82附近的QTL位点贡献率最大,分析其为主效QTL。检测到14个与籽粒容积有关的QTLs,其中7号染色体上RM82附近TQL位点的贡献率较大,分析其为主效QTL。检测到3个与粒质量相关的QTLs,7号染色体上的RM82贡献率最大,分析其为主效QTL。
3.2籽粒容积对粒质量的贡献率最大,粒厚次之
王余龙等研究表明,不同比重谷粒的长、宽差异较小,厚度、容积差异加大[25]。在本研究中,通过对粒长、粒宽、粒厚、籽粒容积、密度、粒质量相关系数的分析表明,除密度与粒质量的相关系数较低外,其余4种粒型与粒质量均呈极显著正相关。其中,籽粒容积与粒质量的相关系数与直接通径系数均为最大,粒厚与粒质量的相关系数与直接通径系数均仅次于籽粒容积,说明籽粒容积对粒质量的贡献率最大,而粒厚次之。通过对重复检测到的QTL位点的分析,发现以籽粒容积、粒质量分别为指标被重复检测到的概率是最大的,分别为RM5984、RM6、RM1089、RM7118、RM4777、ID41、RM481、RM5711、RM82、RM125,其次为粒厚,分别为RM1089、RM7118、ID41、RM5711、RM82、RM125。该结果表明,籽粒容积、粒厚可能与粒质量的遗传基础相近。此外,本研究检测到的粒质量、容积、粒厚的主效QTL均位于7号染色体的RM82附近,分析RM82附近分布着与籽粒性状有关的基因簇。综上所述,籽粒容积对粒质量的贡献率最大,粒厚次之。
3.3除粒质量外其他不同粒型间的相关分析与QTL比较
大部分学者都认为,不同的粒型性状之间存在着一定的相关关系;但也有研究者认为,粒长、粒宽没有相关性[7]。在本研究中,粒长、粒宽、粒厚、容积、密度的主效QTLs分别是1号染色体的RM212、1号染色体的RM1201和9号染色体的RM3787、7号染色体的RM82、7号染色体的RM82、7号染色体的RM5344,而且其中的RM212、RM3787、RM82均被重复检测到。其中RM82在有关粒厚、容积、粒质量的QTLs里均被重复检测到,且贡献率都是最大的,由此说明RM82附近的QTL位点能够稳定遗传,且粒质量、容积、粒厚有较为相近的遗传基础。由本研究可以看出,除容积与密度、粒长与容积之间的相关性不显著之外,其他彼此间都呈极显著相关。其中,粒厚与容积的相关性最大,为极显著相关。其次依次是粒宽与容积的极显著正相关,粒宽与粒厚的极显著正相关,粒长与粒宽的极显著负相关,粒宽与密度的极显著负相关,粒长与密度的极显著负相关,粒长与粒厚的极显著负相关,粒厚与密度的极显著负相关。本研究结果证明,不同粒型之间存在着相关性,这一结论还需要通过构建不同的群体研究,得到大量的证据来进一步验证。
3.4野生稻进化为更有利于提高产量的栽培稻
普通野生稻演化为栽培稻的过程中,经过自然选择和人工选择,由于不断自交,杂合度降低,纯合度提高,在某种程度上满足了现代作物育种提高产量的要求,虽然野生稻还保持着遗传多样性,但就产量而言是极低的[26]。野生稻的粒型呈现的长、细、薄等性状不利于产量提高。
在本研究中,共检测到81个QTLs位点。其中检测到的12个有关粒长的QTLs位点中有9个来源于野生稻等位基因的QTLs位点促进粒变长,检测到的16个有关粒宽的QTLs位点中只有1个来源于野生稻的等位基因的QTL促进粒变宽。其余检测到的有关粒长、粒宽、粒厚、容积、密度、粒质量的71个QTLs位点均来源于栽培稻特青的等位基因的促进效应。这说明在野生稻进化为栽培稻的过程中,一些有利于提高产量的性状受到了人工选择。
参考文献:
[1]Khush G S. What it will take to feed 5.0 billion rice consumers in 2030[J]. Plant Molecular Biology,2005,59(1):1-6.
[2]Doganlar S,Frary A,Tanksley S D. The genetic basis of seed-weight variation:tomato as a model system[J]. Theoreitical and Applied Genetics,2000,100(8):1267-1273.
[3]Fan C,Xing Y,Mao H,et al. GS3,a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein[J]. Theoretical and Applied Genetics,2006,112(6):1164-1171.
[4]马丽莲,郭龙彪,钱前. 水稻大粒种质资源和遗传分析[J]. 植物学通报,2006,23(4):395-401.
[5]高志强. 水稻遗传图谱构建及粒形和粒重QTL定位[D]. 北京:中国农业科学院,2011.
[6]Wang X K,Sun C Q,Cai H W,et al. Origin of the Chinese cultivated rice (Oryza sativa L.)[J]. Chinese Science Bulletin,1999,44(4):295-304.
[7]邱先进,袁志华,何文静,等. 水稻粒型性状遗传研究进展[J]. 江西农业大学学报,2014(4):705-709.
[8]Song X J,Huang W,Shi M,et al. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J]. Nature Genetics,2007,39(5):623-630.
[9]Shomura A,Izawa T,Ebana K,et al. Deletion in a gene associated with grain size increased yields during rice domestication[J]. Nature Genetics,2008,40(8):1023-1028.
[10]Weng J,Gu S,Wan X,et al. Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight[J]. Cell Research,2008,18(12):1199-1209.
[11]Li Y,Fan C,Xing Y,et al. Natural variation in GS5 plays an important role in regulating grain size and yield in rice[J]. Nature Genetics,2011,43(12):1266-1269.
[12]Wang S,Wu K,Yuan Q,et al. Control of grain size,shape and quality by OsSPL16 in rice[J]. Nature Genetics,2012,44(8):950-954.
[13]Qi P,Lin Y S,Song X J,et al. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3[J]. Cell Research,2012,22(12):1666-1680.
[14]Ishimaru K,Hirotsu N,Madoka Y,et al. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield[J]. Nature Genetics,2013,45(6):707-711.
[15]Temnykh S P W,Park W D,Ayres N,et al. Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.)[J]. Theoretical & Applied Genetics,2000,100(5):697-712.
[16]Murray M G,Thompson W F. Rapid isolation of high molecular weight plant DNA[J]. Nucleic Acids Research,1980,8(19):4321-4325.
[17]林鸿宣,闵绍楷,熊振民,等. 应用RFLP图谱定位分析籼稻粒形数量性状基因座位[J]. 中国农业科学,1995,28(4):1-7.
[18]Redona E D,Mackill D J. Quantitative trait locus analysis for rice panicle and grain characteristics[J]. Theoretical and Applied Genetics,1998,96(6/7):957-963.
[19]邢永忠,谈移芳,徐才国,等. 利用水稻重组自交系群体定位谷粒外观性状的数量性状基因[J]. 植物学报,2001,43(8):840-845.
[20]Rabiei B,Valizadeh M,Ghareyazie B,et al. Identification of QTLs for rice grain size and shape of Iranian cultivars using SSR markers[J]. Euphytica,2004,137(3):325-332.
[21]Govindaraj P,Arumugachamy S,Maheswaran M. Bulked segregant sanalysis to detect main effect QTL associated with grain quality parameters in Basmati 370/ASD 16 cross in rice (Oryza sativa L.) using SSR markers[J]. Euphytica,2005,144(1/2):61-68.
[22]赵芳明,朱海涛,丁效华,等. 基于SSSL的水稻重要性状QTL的鉴定及稳定性分析[J]. 中国农业科学,2007,40(3):447-456.
[23]Amarawathi Y,Singh R,Singh A K,et al. Mapping of quantitative trait loci for basmati quality traits in rice (Oryza sativa L.)[J]. Molecular Breeding,2008,21(1):49-65.
[24]Hu W,Wen M,Han Z M,et al. Scanning QTLs for grain shape using a whole genome SNP array in rice[J]. Plant Biochem Physiol,2013(1):104-109.
[25]王余龙,姚友礼,李昙云,等. 水稻籽粒有关性状与粒重关系的初步探讨[J]. 作物学报,1995,21(5):573-578.
[26]张晓丽,郭辉,王海岗,等. 中国普通野生稻与栽培稻种SSR多样性的比较分析[J]. 作物学报,2008,34(4):591-597.
