锌和铜在生猪养殖中的作用及对环境的影响:从饲料到粪污排放到土壤
2023-01-17作者李兰
◆作者:李兰 译
◆文章来源:INRAE动物生产期刊(INRAE:Institut national de recherche pour l’agriculture,l’alimentation et l’environnement,法国国立农业食品环境研究所)。原文链接:https://doi.org/10.20870/productions-animales.2022.35.2.7073
每年,法国的农业和食品工业会产生3亿吨有机排放物,其中有2600万吨粪便和80万吨厩肥来自生猪养殖(Loyon,2017)。这些排放物有很多的营养和能量。然而它们也会对当地和全球的空气、水或者土壤造成污染,对环境、动物和人类产生危害(“同一健康”概念)。因此,我们的挑战在于更好地利用废液中所含的资源(营养物质和能量),以减少其对环境的影响。并在循环生物经济的框架内将它们的农业、能源和经济潜力最大化。
由于锌和铜的较低的吸收率,以及其对健康和生长性能的作用,往往在日粮中会超过动物需求量添加。使用有机排放物时会根据氮磷的含量来推算需要施用的厩肥数量,然而其中锌和铜的含量可能比植物需要量高出4-10倍(Jondreville等,2002;Revy,2003)。由于直接撒播是利用废水的主要方式,因此锌铜等元素会过量被洒播并积聚在土壤中(Coppenet等,1993;L'Herroux等,1997;Jensen等,2018)。这增加了土壤微生物中毒的风险(McGrath等,1995),也增加了对植物的风险(McGrath,1981;Jondreville等,2002;Revy,2003)。此外,在仔猪阶段使用药理学剂量的氧化锌会增加细菌对抗生素的耐药性(Jensen等,2018),这也使得减少药理学剂量氧化锌的使用成了一个重要议题。
为了减少对环境、动物和人类健康的各种风险,欧盟法规对动物饲料中锌和铜的限量越来 越严格(EU-2016/1095;EU-2018/1039)。欧盟同时也存在撒播这些矿物质的有关规定,以及关于肥料和有机土壤改良剂中锌和铜含量的规范或者标准。另外,正在开发的各种有机肥处理技术(堆肥、好氧、厌氧消化等)可能导致这些元素的含量增加,尤其是与干物质相关的浓度(Hsu和Lo,2001;Legros等,2017)。有必要需要了解锌和铜的流向,以便于对其管理。
本文会对锌、铜的流向及其发展变化,从饲料-动物-粪便-土壤进行描述。第一部分回顾了猪料中锌和铜的使用以及猪的需求。第二部分描述了污水中这些元素的排放及其在各种猪污水的管理方式。最后,讨论了生猪养殖中产生含有锌和铜的废水粪便回田对环境的影响。鉴于有关母猪的可用数据数量有限,本摘要主要关注仔猪和育肥猪。
1 猪料中的锌和铜
锌和铜被认为是猪营养中的必要矿物元素。其在饲料原料中的生物可利用量不足以满足生理需要(Manner,2008)。因此在饲料中会额外添加甚至超剂量添加以满足营养学作用或营养学以外的作用,如促生长或者肠道健康。然而动物对其吸收利用率很低,大部分通过粪便排出。因此,减少在饲料中锌铜的添加量,是减少锌铜在粪便、污水中的排放量以及减少对环境的潜在影响的主要手段。与此同时不能影响动物的生长性能或健康。为此,必须更好地了解它们的功能和它们在体内的代谢,这会影响它们的生物利用度,从而影响它们的吸收和排泄。
1.1 锌和铜对猪的作用
1.1.1 体内含量
锌大量存在于猪体内,100kg活体重的动物,其含量大约为1.5~2.5g,主要存在于猪的肌肉组织(占60%)和骨骼(占30%)中。骨骼是主要的储存器官,也就是说当锌供应过量或缺乏时,能储存或动员锌。头发(200mg/kg干 物 质)和 肝 脏(150mg/kg干物质)中的锌浓度最高。与大多数哺乳动物一样,猪的血浆中锌含量约为1mg/L,相当于全身锌的0.1%含量(Swinkels等,1994)。
猪体内铜含量较低,100kg活体重的动物,其含量大约为200mg。铜主要存在于骨骼(40%~46%)、肌肉(23%~26%)和肝脏中(8%~10%)(Cromwell,1997)。当铜超过需求时主要储存在肝脏中。屠宰后测定肝脏铜含量可以作为反应动物体内铜含量及饲料铜含量的指标。在生产中,Hodges和Fraser(1983)观察到不同来源和年龄的猪肝脏样本中铜含量存在巨大差异。9%的情况下,他们观察到肝脏内铜低于12mg/kg干物质时,表明有缺铜的风险。但对于大部分动物来说,其肝脏的铜含量超过20mg/kg干物质,这表明铜被超营养剂量添加。肝脏铜含量不仅仅取决于饲料铜含量,也与铜源的生物学利用率有关(Roméo等,2018)。
1.1.2 生理和催化功能
锌是近200种转录因子的结构和功能完整性的基础。锌是300多种金属酶的辅助因子,大多数的代谢途径依赖于与锌有关的一种或多种蛋白质(Suttle,2010)。它是DNA和RNA合成酶和转运酶及许多消化酶的组成成分(National Research Council,2012)。
铜是参与许多生物功能的必需元素,例如细胞呼吸、防止氧化应激和铁的运输(Suttle,2010),因为其存在于大量的酶、辅因子和蛋白质中(Espinosa和Stein,2021年)。
锌和铜在防止氧化应激方面发挥作用,这是因为它们作为蛋白质和酶的辅助因子,可防止氧化应激(超氧化物歧化酶、铜蓝蛋白氧化亚铁酶)。缺锌会增加内皮细胞对氧化应激的敏感性。锌是金属硫蛋白的激活剂,后者会参与细胞解毒,对锌和铜有很强的亲和力,此外,金属硫蛋白还表现出抗氧化特性(Suttle,2010)。
在药理学剂量下,锌和铜可以对猪起到促生长作用。这可能与改善动物的消化吸收率和增加食欲有关(Suttle,2010),以及与减少如腹泻等消化道问题有关(Bikker等,2016),还有与其抑菌、改善肠道微生物群落和肠道完整性有关系(Villagómez-Estrada等,2020a)。
1.2 猪的需求
动物对某种元素的需求对应于为使其有机体完成其所有生理功能而需提供的数量(Revy,2003;Schlegel,2010)。这个量会因不同的相应标准的定义而有所不同。动物生理需求的满足取决于日粮中微量元素的生物学利用率(National Research Council,2012)。定义需求量时还需要考虑提供的形式。在生产中,需要了解动物对锌铜需求随着其年龄的变化,以便随时调整日粮,从而降低提供不足时对生产性能的影响,或者过量时对环境的影响。
1.2.1 对仔猪和生长猪需求量的评估
有两种评估矿物质需求的方法,一种是经验法,一种是阶乘法(Schlegel,2010)。前者是对于金属元素最常见的方法。这包括通过试验测试逐渐增加日粮中锌和铜含量的影响,以评估动物的不同现场和生理参数的反应。然后通常使用线性模型来描述响应,并可以在拐点处预测动物的需求(Kirchgessner,1993)。
必须观察动物对锌或铜的功能之一的特定和代表性反应,以确定作为需求特征的标准(Schlegel,2010)。对于 锌,通常采用的最具代表性的标准是其在骨骼或血浆中的浓度;对于铜,这个标准是其在肝脏中的浓度(Jongbloed,2010)。
断奶后仔猪的锌含量如图1所示,数据是基于各种参考文献。因此根据摄入量,我们可以定义①缺乏区(红色区),其中摄入量不足以维持足够的血浆水平;②由于建立了不同的稳态机制(储存、排泄等),血浆水平保持恒定的稳态区(绿色区);③过量区域(蓝色区),其中血浆浓度随着食物摄入而增加,体内稳态平衡机制可能已饱和。红色和绿色区域之间的拐点可以定义营养需求,而绿色和蓝色区域之间的拐点表明出现与采食量下降相关的过量风险。
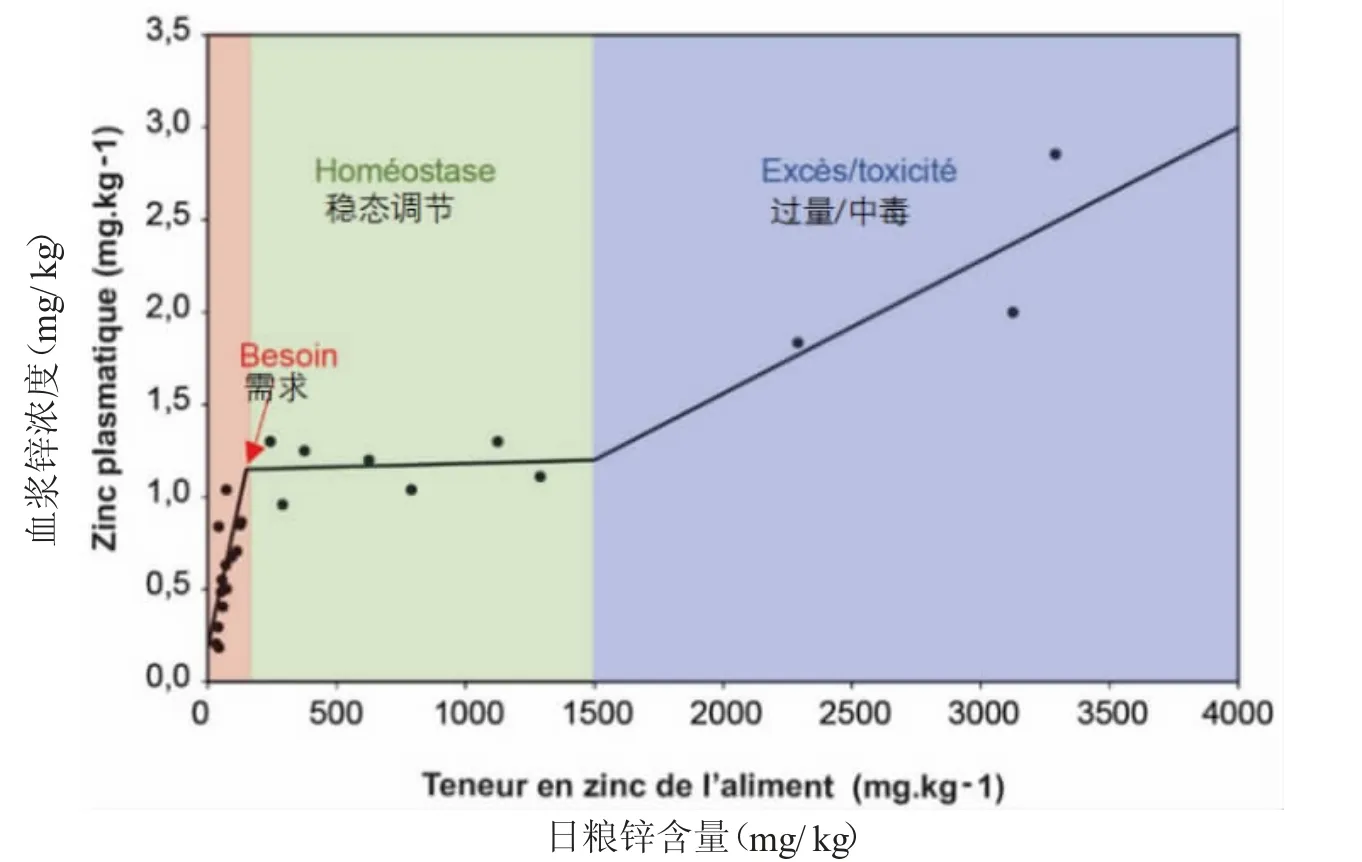
图1 断奶后仔猪血浆锌浓度随饲料中锌含量的变化(根据Hahn和Baker,1993;Hill等,2001;Revy,2003)
2012年NRC根据动物的生长阶段或生理阶段对猪的锌需求量给出不同的参考。断奶仔猪的锌需求量约为80mg/kg。NRC(2012)给出育肥猪基础日粮中平均需求量约为50 mg/kg干物质饲料。这个需求可能会根据锌来源生物利用度的不同而有所不同:日粮的性质、钙和植酸盐含量以及植酸酶的添加等(Revy,2003;Spears和Hansen,2008)。
仔猪对铜的严格的需求约为5~6 mg/kg干物质饲料,并且对于更高级的生理阶段也不超过该值(NRC,2012)。然而,这个需要量并未考虑在动物的某些关键阶段(如断奶时或繁殖母猪)高铜对生长性能或消化健康的有益影响。对铜的需求可能会根据锌的生物利用度的不同而变化,例如在食物中添加更多的锌。
1.2.2 缺乏效应
缺锌的主要症状,一开始是缺乏食欲、出现腹泻和生长迟缓。厌食是最早观察到的迹象,接着出现缺锌的仔猪股骨大小和坚固性的降低(Suttle,2010)。出现腹泻是由于肠细胞更新减少(Revy,2003)。此外,可以观察到血浆或血清中锌水平以及碱性磷酸酶和白蛋白水平降低(Suttle,2010;NRC,2012)。然而,动物通过调节各种肠道锌转运蛋白来应对锌缺乏症,但这不足以弥补缺锌仔猪血浆和血清中锌浓度的过早下降(Suttle,2010)。缺锌最突出的症状和最后出现的症状是称为角化不全的皮 肤 过 度 角 化(Suttle,2010;NRC,2012)。
很少观察到猪缺乏铜的情况,可能出现在断奶仔猪饲喂大约100mg/kg干物质的Fe、130mg/kg干物质的锌和2 mg/kg干物质的铜时会发生缺铜(Jondreville等,2002)。缺铜的明显迹象是贫血、四肢弯曲、自发性骨折、心脏和血管问题以及脱色(NRC,2012)。然而,最近的结果(Dalto等,2021)表明,在以药理剂量(2500~3000 mg/kg干物质)摄入锌的情况下,存在铜缺乏的风险,从而证实了Hill等(2001)观察到的血浆铜减少等(见1.3.4)。
1.2.3 过量效应
锌对于大部分哺乳动物毒性较小。然而,日粮中过量的锌会导致其吸收利用率降低,与金属硫蛋白结合后,其在骨骼和肠细胞中的储存增加,因此会增加锌在组织和内源性分泌的更新。在药理学剂量下,过量的锌会导致采食量降低,因此生长速度也会降低(Hahn和Baker,1983)。这会导致铜和铁代谢的改变,如果饲喂过量锌较长一段时间,则有缺铜和缺铁的风险(Dalto等,2021)。此外,动物对过量锌的敏感性还取决于饲料中的钙、铁、铜和镉的水平(Revy,2003)。
猪的铜中毒案例比较少见(Jondreville等,2002)。然而,长期饲喂超过250 mg/kg干物质的铜会导致中毒,观察到的迹象是血红蛋白水平降低和黄疸(NRC,2012)。这会导致肝脏中铁的储存减少,从而导致贫血,这是由于铜对铁吸收的负面作用,铁的吸收受到限制。猪对高剂量铜的耐受性取决于饲料中锌的供应以及锌和铜与金属硫蛋白的相互作用(Jondreville等,2002)。
最后,饲料锌铜的含量能否满足猪对其的需求以及这些元素潜在的毒性作用,取决于它们的含量,也取决于其生物利用率(Hahn和Baker,1983;Suttle,2010)。吸收率和效率会影响动物对它们的总需求量。
1.3 饲料中的锌和铜
1.3.1 法规
2003年以来,欧盟修改了关于猪饲料中锌和铜的最高限量(见图2)。
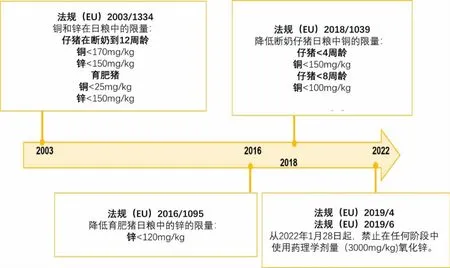
图2 欧盟关于生长猪料中锌和铜最高限量的法规的变化
从2022年1月28日起,欧洲将禁止在兽药处方中使用氧化锌。这些法规旨在减少粪便中锌的排泄(Revy,2003)及其在土壤中的蓄积,并降低动物和人类抗生素耐药性风险,而高锌会增加这种现象的风险(Ciesinski等,2018)。
1.3.2原料中的组成和含量
猪的日粮主要由谷物(主要是玉米和小麦,还有大麦、黑小麦等)、饼粕(主要是大豆,还有油菜籽和向日葵)、油料和蛋白质作物(豌豆……)、谷物副产品(麸皮、粗粮、谷物……)和维生素矿物质组成(额外添加)。
谷物和蛋白原料平均含20~30 mg/kg干物质的锌,谷物副产品的锌含量较高(70~90 mg/kg干物质,见图3)。油籽和饼粕类含30~90 mg/kg干物质的锌。谷物、其副产品、油籽和油饼含5~15mg/kg干物质的铜。

图3 猪料中锌和铜的含量(根据INRA-AFZ,2004的数据库)
1.3.3 不同形式的补充剂
为满足动物需要,猪饲料中需要额外添加锌,因为仅从原料中摄入是不够的。此外,还必须考虑到日粮中不同成分导致的日粮锌生物利用率的变化(Revy,2003)。
与锌不同,理论上饲料中不额外添加铜是可以满足生长猪对铜的需求(约6 mg/kg)。然而在实践中,通常会在饲料中额外添加铜,以考虑到总需求量估计的不精确性,并抵消日粮中某些元素的拮抗作用,影响铜的生物利用率,从而影响对铜的需求。猪料中的额外加铜是作为安全边际的作用(Jondreville等,2002)。
作为添加剂添加的锌和铜可以来自不同的来源。比较生物利用率的研究中通常用硫酸盐(ZnSO4和CuSO4)作为参考来源。然而这些硫酸盐溶解速度太快,这意味着它们也更容易与日粮的其他元素结合,特别是许多原料中所含的植酸盐,从而降低其生物利用率(Revy,2003)。铜和锌还存在其他补充来源(氧化物、氯化物、螯合物),其物理化学特性和生物利用率差异很大。在实践中,ZnO和CuSO4是最常用的动物饲料来源(Revy,2003)。(译者注:氧化锌在欧洲是最常用的锌源,在中国和欧洲有所区别,因本文是以欧洲为背景,特此解释)
1.3.4 饲料中铜和锌的生物利用率和变异因素
存在两种实验方法来测量矿物质的生物利用率。第一个方法是测量在低于动物需求的剂量下,不同生理指标对逐渐增加矿物质摄入量的反应。第二个方法是在摄入量远高于需求的剂量下,测量这些生理指标,并测量元素在血液或不同组织(如骨骼或肝脏)中的积累(Spears和Hansen,2008)。这两种方法,均以硫酸盐形式的矿物质为参考(CuSO4/ZnSO4),其吸收利用率定义为100%。然后通过计算不同试验下的响应斜率来计算生物利用率。一般来说,微量元素的生物利用率受微量元素的来源、动物生理状态或其矿物质状态等因素的影响,以及与日粮、促进剂或拮抗剂等其他元素相互作用的影响(O′Dell,1989;Manner,2008)。不同类型的相互作用是由于在肠道中形成不可吸收的复合物、阳离子之间竞争非特异性二价阳离子的转运或非特异性金属对蛋白质的诱导(Suttle,2010)。可以用这些影响其吸收的因素以及影响其溶解度的不同物理化学性质,来解释不同来源的锌和铜生物利用率的巨大差异。
植酸盐与锌形成不可吸收的复合物,从而降低锌的生物利用率(Suttle,2010)。植酸盐水解后能释放出锌,但单胃动物不会自然产生植酸酶,或者数量非常有限。因此,在日粮中添加微生物植酸酶可以提高锌的利用率Revy等(2004)揭示了700个单位的植酸酶可以取代32~43 mg硫酸盐形式的Zn。同样,Bikker等(2012)估算500单位的植酸酶可以替代27 mg硫酸盐形式的锌。根据Schlegel和Jondreville(2011)进行的荟萃分析,只有食物中天然存在的Zn会受到植酸盐和植酸酶的影响,而不是以矿物质或有机补充剂的形式添加到日粮中的锌。在日粮中添加钙后,植酸盐的这种拮抗作用会增强(Suttle,2010)。植酸盐-钙-锌复合物在消化道中形成(Revy,2003)。植酸盐对锌的拮抗作用在幼畜中更为重要,因为其日粮中钙含量往往超过了它们的需要(Suttle,2010)。
铜受日粮中植酸盐和钙的影响较小,因为它对游离氨基酸具有更高的亲和力,与游离氨基酸形成螯合物,从而使铜保持其溶解度(Jondreville等,2002)。然而,可溶性铜源也可在消化道中与植酸盐相互作用并形成锌-钙-铜-植酸盐或铜-钙-植酸盐络合物(Oberleas,1973),影响植酸酶的作用。
锌对铜有拮抗作用,即会抑制其吸收。这种作用是由于锌对金属硫蛋白合成的诱导作用,金属硫蛋白与铜有很强的亲和力,会阻止其转移到肠上皮(Revy,2003)。
