北极楚科奇海北部特征水团对浮游植物空间分布的调控
2023-01-12费淑怡李纪高源葛跃浩郭正东高咏卉
费淑怡,李纪, ,高源,葛跃浩,郭正东,高咏卉, *
( 1. 上海交通大学 海洋学院,上海 200030;2. 中国极地研究中心 自然资源部极地科学重点实验室,上海 200136;3. 上海交通大学 海洋学院 上海市极地生命过程与环境重点实验室,上海 200030;4. 国家海洋局南通海洋环境监测中心站,江苏南通 226002;5. 中国人民解放军海军潜艇学院,山东 青岛 266199)
1 引言
北极的气温增速约为全球平均值的两倍,这造成海冰面积减小、多年冰减退、无冰区增加,对脆弱的极地生态环境产生深远影响[1]。自1978年以来,北极夏季冰盖面积大幅下降,2021年夏季海冰面积仅为1981-2010年平均面积的77%左右[2]。而且多年冰占总海冰面积的比例从40年前的70%降低至近年来的30%[3],导致冰层逐渐变薄,透光性增强,利于浮游植物生长。此外,海冰开始融化的时间提前,冻结得更晚,无冰期随之延长[1]。
随着北极海冰面积减小、透光增强、开放时间延长等变化,北极的水文特性和环流结构也发生变化,对营养物质的输运、物种的分布、群落结构和生态环境等产生重要影响。北极浮游植物的生物量总体呈增加趋势[4-5],1998-2012年,海冰面积减小是驱动生物量和生产力增加的主要原因。此后,海冰损失明显减缓,营养供应增加可能是近10年来的浮游植物生物量增加的主要原因[1]。同时,藻华出现时间更早,秋季二次藻华的报道也有所增加[6-7]。浮游植物通过光合作用吸收CO2,对调节CO2的海气交换、收支和存储起着重要作用[2-3,7];作为初级生产者,它们也为鱼类、浮游动物和底栖动物提供食物,对生态系统起到关键的支持作用[8]。
北极冰下藻华可以从冰边缘一直延伸至海冰深处[4]。海冰中普遍存在藻类(如海链藻Thalassiosira,拟脆杆藻Fragilariopsis, 齿状藻Odontella等)的休眠孢子,入水萌发,为夏季藻华的形成提供种源。也有少量报道表明,北极水中发现的硅藻能长时间在黑暗中存活[5]。当冰雪融化或者融池形成后,为冰下的浮游植物带来适宜的光照、削弱紫外线辐射,楚科奇海冰层下的浮游植物就开始生长。冰下寒冷的水温也造成浮游动物的摄食速率降低[9],这使得冰下区域存在适宜浮游植物生长栖息的区域。浮游植物对环境的适应性有利于它们在高纬度地区的生存,并影响春、夏季藻华中浮游植物的群落结构。
在北极,冰下浮游植物对海洋碳汇的贡献量约占80%,它们的丰度和分布也造成光合效率、初级生产力和碳通量的区域性差异[3,7]。楚科奇海作为北极气温增速最快的海域之一,扩大的无冰区面积和延长的开放时间均居北极的前列,近20年叶绿素浓度和生产力也显著增加[7]。浮游植物以硅藻为主,叶绿素a浓度(3~800 mg/m3)和初级生产力(以碳计,0.2~463 mg/(m2·d))均呈现明显的时空变化[5]。然而,过去的研究多集中在楚科奇海南部无冰或少冰区域[10-12],北部边缘海区的数据不足,缺少对冰下浮游植物及其生存环境的系统性分析。
本文依托中国第12次北极科学考察,对楚科奇海北部边缘海与加拿大海盆西侧(74°~78°N,150°~170°W)进行了生态调查(图1)。结合水团等环境特征,采样并分析密集冰区中营养盐和叶绿素a浓度的分布特征。通过厘清水团分布,探究环境因子变化与浮游植物分布之间的关系,构建北极在环流作用下的物理-化学-生物间的相互作用,有助于评估北极气候变化对海洋生态系统的潜在影响。

图1 西北冰洋环流示意图(a)和采样区的海冰密集度在调查期间的均值(b)Fig. 1 Schematic diagram of the western Arctic Ocean circulation (a) and the average of sea ice concentration during the sampling time (b)
2 材料与方法
2.1 研究区域
2021年7月25日至8月1日,在北极75°N以北的高纬度冰区的3个断面开展调查(图1)。R断面沿170°W经线,由南向北穿过楚科奇深海高原,其底部为典型的深海海沟和山脊,部分区域呈现平坦的高原状峰顶。P1和P2断面大致平行,跨越北风海脊,向东延展至加拿大海盆,水深从200 m骤增至4 800 m(图1a)。各站位均位于海冰边缘线以北(图1,https://seaice.uni-bremen.de/databrowser)。
研究区域位于加拿大海盆边缘,混合层以下的极地表层水包括夏季、冬季白令海水和阿拉斯加沿岸流(Alaskan Coastal Water,ACW),物理环境复杂[13]。太平洋夏季水(Pacific Summer Water,PSW)流经白令海峡后,携带的陆坡沉积物释放营养盐,形成营养盐丰富的白令海夏季水(summer Bering Sea Water,sBSW),向北流向北极的中央海区[14]。PSW的东侧支流沿着阿拉斯加沿岸向东北流动。在巴罗峡谷附近,受波弗特高压和加拿大海盆地形的共同作用,大部分ACW沿加拿大海盆汇入顺时针运动的波弗特流涡(Beaufort Gyre, BG)[15];其余沿阿拉斯加陆架继续向东输送[16]。由于波弗特高压强度和位置的改变,BG影响的范围和强度往往呈现明显的时空变化[17]。每年的6-7月,阿拉斯加周边的河流径流量最大,海冰融水增加,往往造成BG覆盖区域内的淡水储量增加[18],BG影响范围(70°~80°N ,130°~170°W)在8月达到最大[15],形成营养盐浓度和盐度略低的ACW暖水团向本研究区域输送。
2.2 观测和取样
利用“雪龙2”号的破冰能力实现冰区采样。站位水温和盐度通过温盐深剖面仪(CTD 911,Seabird InC.)测定。水样在表层、10 m、20 m、50 m、75 m、100 m、150 m、200 m以及叶绿素a浓度最大层(ChlorophyllaMaximum Layer,CML)采集,用于营养盐和叶绿素a浓度分析。
2.2.1 营养盐浓度测定
各层采集的水样经孔径0.45 μm的醋酸纤维滤膜过滤,冷冻存储,用于生物可利用溶解态营养盐的测定。其中,磷酸盐(SRP)、硅酸盐(Si (OH)4)、硝酸盐( NO-3-N)和亚硝酸盐( NO-2-N)浓度依据《海洋监测技术规程 第1部分:海水》(HY/T 147.1-2013)[19]流动分析法,由Skalar San++营养盐自动分析仪(荷兰,SKALAR)测定。氨氮( N H+4-N)浓度依据《海洋监测技术规程 第1部分:海水》(HY/T 147.1-2013)[19]次溴酸盐氧化法测定。各溶解态营养盐的检出限分别为: NO-3-N 0.04 μmol/L、 NO-2-N 0.025 μmol/L、 NH+4-N 0.015 μmol/L、SRP 0.02 μmol/L、Si (OH)40.04 μmol/L。溶解无机氮(DIN)浓度为 NO-3- N、 N O-2- N和 N H+4-N浓度的总和。
2.2.2 叶绿素a浓度测定
依据《海洋监测技术规程 第1部分:海水》(HY/T 147.1-2013)[19]的荧光法测定叶绿素a浓度。先对各站位200 m以上水层的样品进行粒度分级:量取250 cm3的水样先经孔径20 μm的筛绢进行过滤后(分离获取小型浮游植物细胞,Netphytoplankton),再经孔径为2 μm的 玻 璃 纤 维 素 酯 滤 膜 过 滤(分 离2~20 μm的微型浮游植物细胞,Nanophytoplankton),最后用Whatman GF/F玻璃纤维滤膜过滤(截留微微型浮游植物细胞,Picophytoplankton)。滤膜冷冻避光保存。以90%丙酮对滤膜上的浮游植物细胞萃取24 h后,在10AU型唐纳荧光仪上测定并计算,得到Chla浓度[20]。
2.3 数据分析
运用SPSS 26.0软件对数据进行统计学分析。R1、R3站位和其他站位之间的环境因子差异采用一元方差分析(One-way ANOVA)进行分析。环境因子和分级叶绿素浓度之间的相互关系用Spearman相关系数来表示。
3 结果
3.1 海区冰情、水文分布及水团特征
自1978年以来,虽然北极夏季冰盖面积的总体变化趋势大幅下降,但2021年夏季海冰覆盖面积略高于2015-2020年[2]。调查期间,高纬度冰区刚开始融化。受西北风影响,海冰向研究区域移动和堆积。海冰密集度的均值约为75%,冰厚往往高达1 m左右。除了沿P2断面零星出现海冰密集度约50%的区域,绝大多数区域由冰覆盖(海冰密集度为90%~100%,图1b)。
冰下0~150 m,水温介于-1.75~0.8℃之间,盐度为27~35,呈现复杂的层化现象(图2)。海冰融化、混合形成低盐的融冰水(Melting Water,MW)。融冰水以下,根据Steele等[14]和Wang等[21]对楚科奇海和加拿大海盆地区中夏季盐跃层的划分,可分为ACW和sBSW两个暖水团,ACW水温往往略高于sBSW。向下依次为低温的白令海冬季水(winter Bering Sea Water,wBSW)和高温高盐的大西洋水(Atlantic Water,AW)。
本次研究区域存在相似的垂向和水平结构(图2)。融冰水使表层形成水温接近于海水冰点(-1.7℃)、盐度小于30的低密度水层,从表层延伸至20 m上下。融冰水以下,盐度随深度增加而逐渐增大,温度垂向分布存在明显差异。在次表层(30~100 m),3个断面均形成狭长的温度略高的水舌。在R断面北部、P1断面、P2断面(8-10站位)尤为明显,水温向加拿大海盆边缘方向逐渐降低,该暖水团(温度:-1.25~0.75℃,盐度:30~32)符合受BG驱动的ACW特征。在R断面南部的次表层(50~100 m)为sBSW暖水团(中心温度约-1.5℃,盐度为31~32),核心温度呈随纬度升高而降低的趋势。在较冷的wBSW(70~125 m)下方,出现高温高盐的AW水团(图2,表1)。
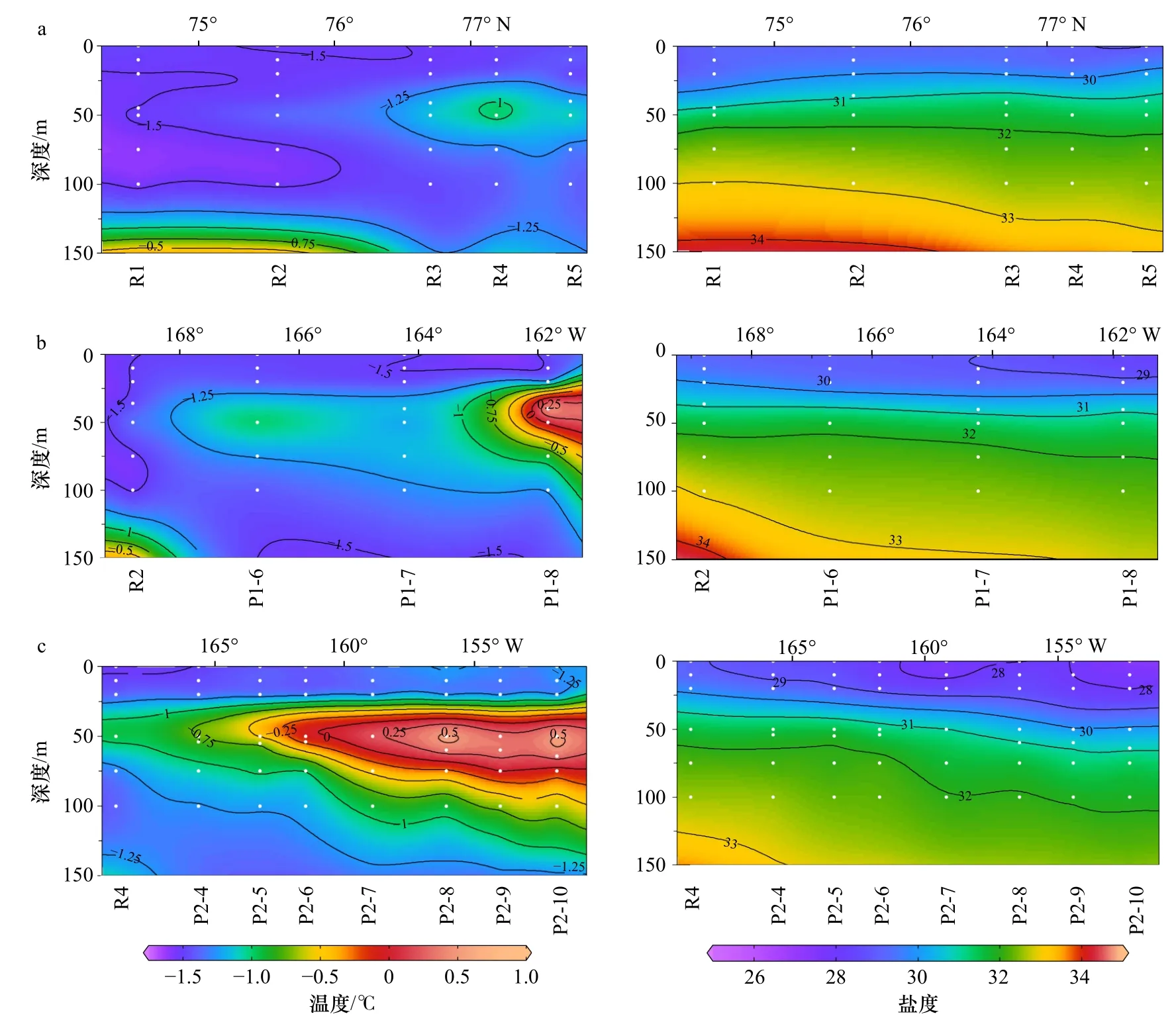
图2 在 R(a)、P1(b)和 P2(c)断面水温和盐度的垂向分布Fig. 2 Vertical distribution of temperature and salinity in the R (a), P1 (b), and P2 (c) transections

表1 不同水团温度、盐度、密度的分布Table 1 Distribution of temperature, salinity and density in different water mass
3.2 营养盐的分布
生物可利用溶解态营养盐的浓度变化范围分别为DIN 0.47~18.19 μmol/L,SRP 0.23~2.65 μmol/L和Si(OH)41.08~39.06 μmol/L(图3)。总体而言,营养盐浓度随着深度增加而逐渐升高。在0~20 m的MW中生物可利用的营养盐浓度最低(DIN浓度<2.5 μmol/L、SRP浓度<1.5 μmol/L、Si(OH)4浓度<5 μmol/L)。50~100 m为营养盐浓度的跃层,营养盐的浓度均随深度迅速增加。受BG影响,在P1、P2断面呈现DIN等营养盐跃层从深海向加拿大海盆边缘地带(从东向西)逐渐变浅的现象。R断面(1~3站)在sBSW影响下,自南向北营养盐浓度逐渐降低。100 m以下营养盐浓度普遍较高,在靠近AW的125~150 m出现营养盐浓度的最高值。DIN浓度与SRP浓度之比多低于16,存在氮的相对限制。
3.3 叶绿素分布
Chla的浓度为0~0.69 mg/m3,垂向变化明显(图4)。分布规律与水团的密度层化一致性良好。低营养盐的表层MW中的Chla浓度低,部分区域几乎为0(图3,图4)。在40~60 m的次表层暖水团区域内,尤其是ACW和sBSW的上边缘,往往出现叶绿素a浓度最大值(平均值±标准差=(0.327±0.163)mg/m3)。Chla浓度在R断面普遍高于P1和P2断面(图4)。在P1断面,Chla浓度从西向东向加拿大海盆边缘逐渐增加。在60 m以下,光强随着深度增加而减弱,Chla浓度也逐渐降低,100 m以下Chla浓度几乎为0。

图3 溶解态无机氮(DIN)、磷酸盐(SRP)和硅酸盐(Si(OH)4)浓度在 3 个断面的垂向分布Fig. 3 Vertical distribution of dissolved inorganic nitrates (DIN), soluble reactive phosphates (SRP) and silicates (Si(OH)4) concentration in three transections
结合垂向分布(图2至图4)和温-盐-深的点聚图(图5a,表1),可以看出该区域的层化现象除了长期存在的白令海冬季水(wBSW)和深层的大西洋水(AW)外,主要受融冰水(MW)、阿拉斯加沿岸流(ACW)和白令海夏季水(sBSW)入侵水团的影响。以对浮游植物生长有潜在限制的DIN为例,结合温-盐-叶绿素a浓度(图5b,图5c)进行点聚分析:DIN浓度在MW中最低(0~2.5 μmol/L之间)。在次表层暖水团中,DIN在ACW水团中浓度近似于sBSW水团,与叶绿素a浓度高值区相吻合。在125 m以下的AW水团中则呈现高DIN浓度和低Chla浓度的特征。
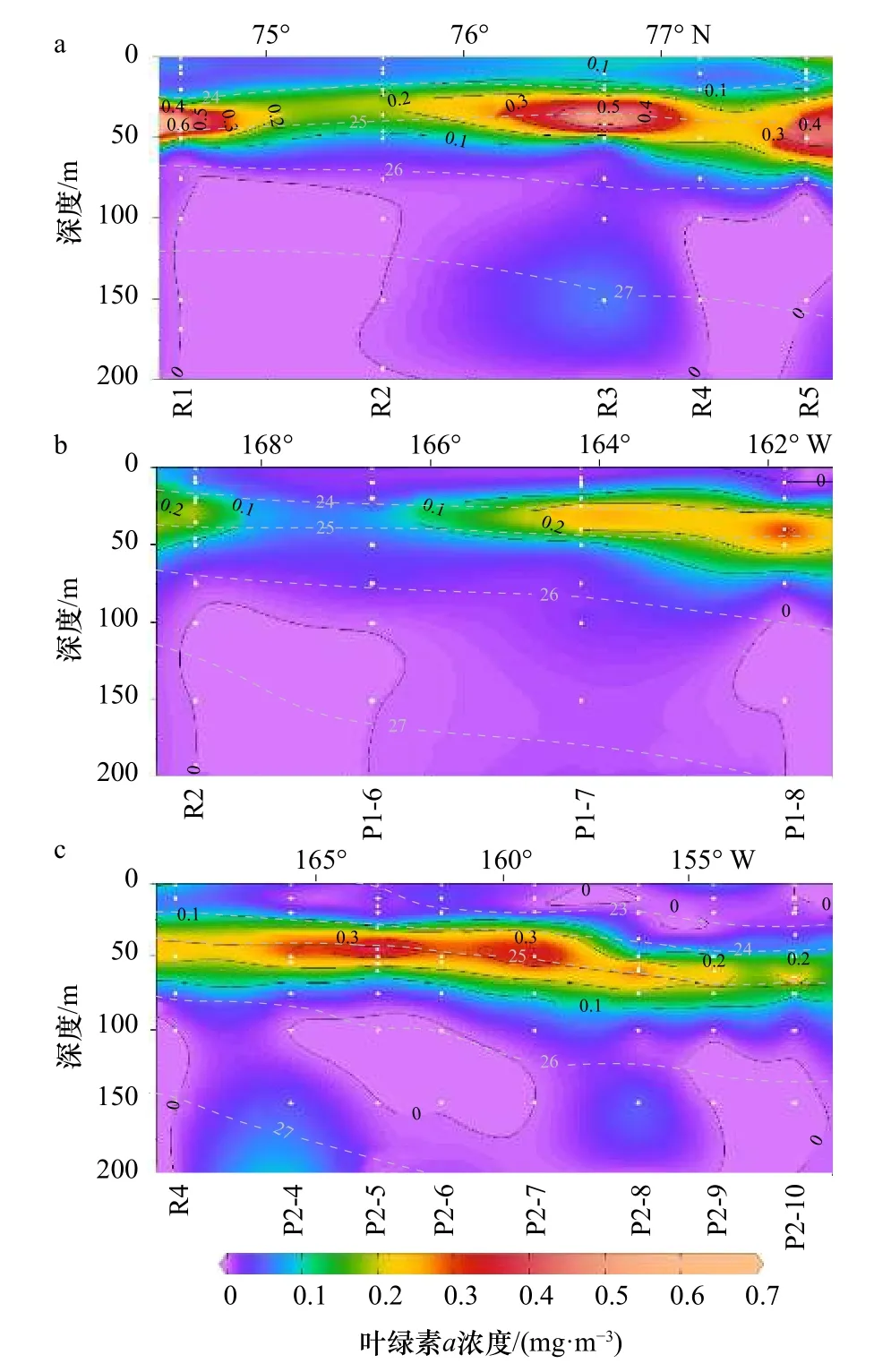
图4 在 R(a)、P1(b)和 P2(c)断面叶绿素a浓度的垂向分布(灰色虚线为等密度线)Fig. 4 Vertical distribution of chlorophyll a concentration in the R (a), P1 (b), and P2 (c) transections (gray dotted lines are equal density lines)
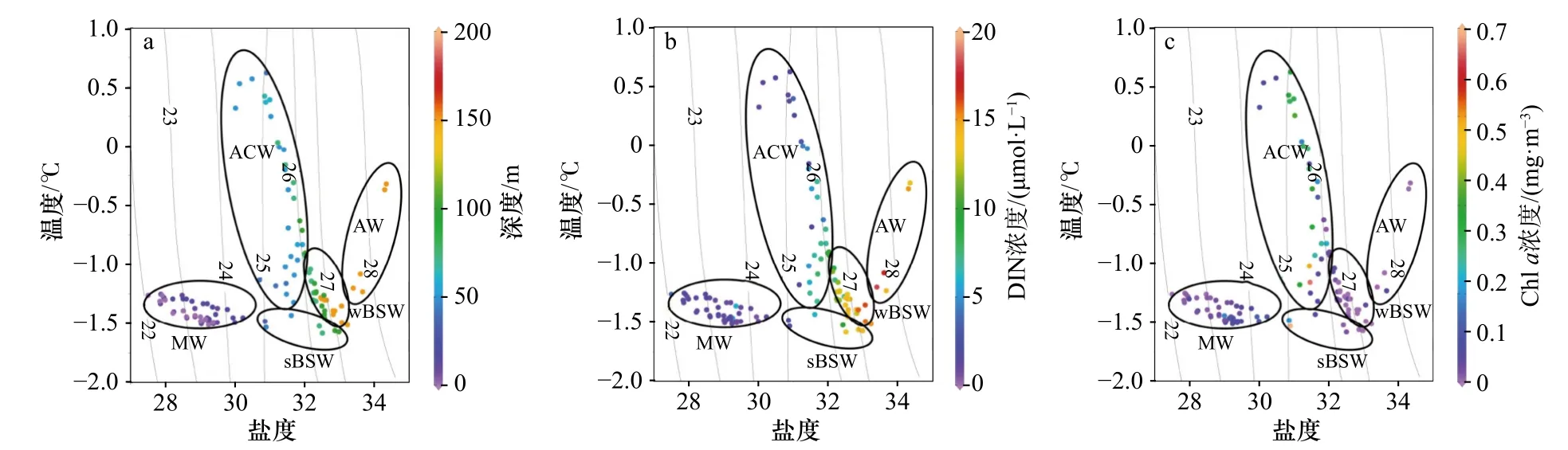
图5 150 m 以上水体的温-盐-深度点聚图(a)、温-盐-DIN 浓度点聚图(b)和温-盐-叶绿素 a浓度点聚图(c)Fig. 5 Depth distribution in temprature-salinity-depth scatter diagram (a), temperature-salinity-DIN concentration scatter diagram (b)and temperature-salinity-Chl a concentration scatter diagram (c) above 150 m
3.4 不同粒径叶绿素a浓度的分布及其与环境因子的关系
各站位叶绿素a浓度最大层处的分级Chla浓度分布如图6所示。sBSW影响下的R1、R3站位小型浮游植物叶绿素a浓度占比70%以上,这两个站位的温度明显低于其他站位(One-way ANOVA,p=0.053<0.1)。在其他站位,浮游植物以微微型为主,微微型浮游植物Chla浓度范围为0.03~0.28 mg/m3,占总叶绿素a浓度的40%~80%(均值约为65%)。微型浮游植物叶绿素a浓度在所有站位中比例均低于30%。
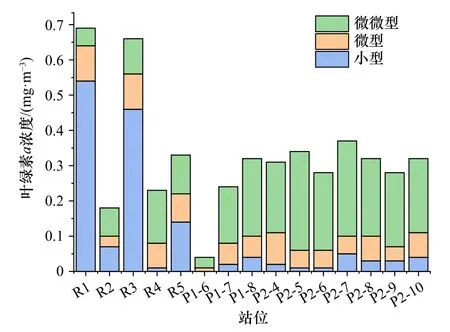
图6 叶绿素a浓度最大层的浮游植物叶绿素a浓度的分级分布Fig. 6 Size-fraction distribution in Chl a concentration of phytoplankton in Chl a maximum concentration layer
各粒级浮游植物叶绿素a浓度与环境因子的Spearman相关性分析(表2)表明:小型浮游植物Chla浓度受多种环境因子的影响,与盐度、DIN浓度、SRP浓度、Si (OH)4浓度和N/P值都呈显著性负相关;微微型浮游植物Chla浓度与温度呈显著正相关,受盐度和营养盐的影响较弱。
4 讨论
4.1 环境因子对浮游植物的影响
叶绿素浓度的垂向分布与2003年[20]、2008年[22]我国在北风海脊附近的观测相似。由于太阳辐射受海冰阻挡,海水中光照随深度增加而减弱。各站位的叶绿素a浓度最大层位于50 m上下,光合有效辐射(PAR)范 围 为1~27 E/(m2·d),平 均PAR为(8.60±8.08)E/(m2·d)。虽然叶绿素a浓度最大层光较弱,但是越来越多的观测结果揭示了北极浮游植物在冰下和极夜的低光照环境中,以消耗脂类、碳水化合物和蛋白质来提供能量,降低代谢速率和细胞分裂等机制,维持Chla浓度增长和光合作用,形成了独特、长期的暗适应能力[23-25]。浮游植物在PAR小于10 E/(m2·d)受 到 光 限 制,但 仍 能 在PAR低 至1 E/(m2·d)的环境下存活[6,23-24]。叶绿素a浓度最大层处微弱的光强也能支持浮游植物生长。由此可见(表2),光照对叶绿素a浓度最大层处浮游植物的分级Chla浓度的影响不显著。
浮游植物的叶绿素a浓度最大层与营养盐的分布密切相关。虽然表层光照更适宜浮游植物生长,但寡营养的融冰水在表层融冰层造成营养盐(尤其是氮)的限制。太平洋入流水中经阿拉斯加沿岸进入加拿大海盆的ACW,富含营养盐、相对温暖,在次表层创造了相对适宜浮游植物生长的环境。在叶绿素a浓度最大层处,DIN∶SRP在1.0~4.1之间,而Si(OH)4∶DIN在1.9~3.7之间。与Redfiled Ratio(C∶N∶P∶Si=106∶16∶1∶16)相比,虽然N存在相对限制,但仍高于限制浮游植物生长的阈值(0.1 μmol/L)[26-27],对浮游植物不构成绝对限制。在叶绿素a浓度最大层处,浮游植物中耐受性强的物种会对N产生明显的生物吸收。
从浮游植物粒径分布来看,除了R1和R3站位的小型浮游植物占比较高(约70%)外,微微型浮游植物在冰区占主导地位。这一结果与Joo等[28]在2008年楚科奇海夏季浮游植物中的发现相符。Smith[29]也发现浮游植物的粒径与冰覆盖面积和营养盐浓度有关:在冰覆盖超过50%的海区浮游植物以微微型为主,而在层化强、营养盐充沛的区域则以小型浮游植物为主。由Spearman分析(表2)可以看到,微微型浮游植物多出现在温度略高、营养盐浓度较低的水层,并成为该区域的优势种。在低温但营养盐浓度较高的R断面,以R1和R3为代表的站位中,小型浮游植物占比较高,形成了以小型浮游植物为主的浮游植物群落。这些粒径不同的浮游植物群落在北极生态系统的群落结构和营养关系中起着重要作用。微微型浮游植物的粒径更小,具有更大的比表面积,代谢和能量转换效率高,通过冰下和融冰初期生物碳吸收、食物链间的传递利用和生物泵作用,在楚科奇边缘海区的物质循环与能量流动中起着不可替代的作用[20]。
4.2 生物作用对叶绿素a浓度最大层处营养盐去除的影响
本文通过水团分析发现叶绿素a浓度最大层与太平洋入流水的传输和区域性分布密切相关。叶绿素a浓度最大层中,不同粒径的Chla浓度与营养盐浓度呈负相关(表2),浮游植物对营养盐的吸收造成了营养盐浓度降低。
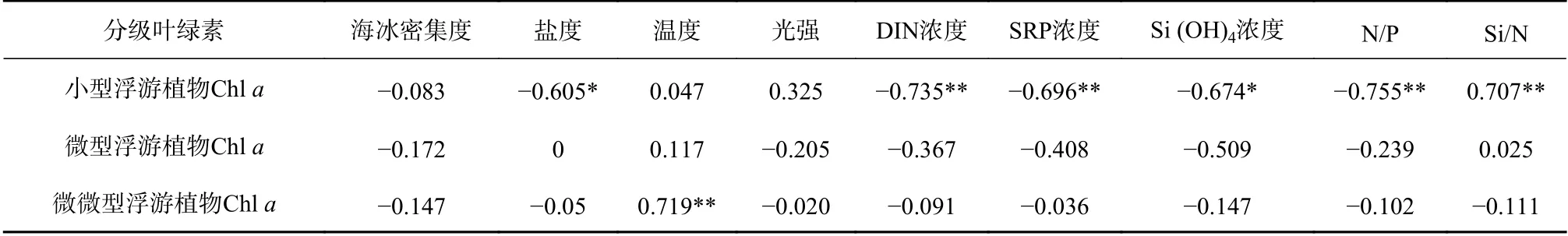
表2 分级叶绿素a浓度与环境因子的相关性Table 2 Correlations of size-fraction chlorophyll a concentration and environmental factors
为区分水团混合和生物作用对营养盐浓度的改变,在接近叶绿素a浓度最大层,营养盐吸收较旺盛的50 m处作图(图7)。由于在氮限制的水体中,DIN浓度与盐度的关系优于SRP浓度与盐度的关系。假如在ACW与sBSW水团物理混合的理想状态下,DIN浓度与盐度保守混合线呈线性关系[30]。然而,实测DIN浓度与盐度关系呈凹形,并且DIN浓度低于混合线(虚线)。一般而言,浮游植物对营养盐吸收量越高,DIN缺失量(ΔDIN)越高。这从叶绿素a浓度分布看,P2断面与R断面相差不大,P1断面浓度较小(图4), 而在对氮去除效率上,P1断面与R断面相差不大,但是稍逊P2断面。受ACW暖水团影响的P1和P2断面,Chla浓度与ΔDIN成正比。因而,浮游植物的丰度越高,对营养盐吸收越强。由于ACW平均温度高于sBSW,浮游植物的氮吸收效率往往随温度升高而增强。这导致ΔDIN在Chla浓度近似的P2断面均高于R断面,ΔDIN在低Chla浓度的P1断面近似于R断面(图7)。这也进一步证明了在以小型浮游植物为主的R断面,其浮游植物的营养盐吸收效率低于以微微型浮游植物为主的P断面。
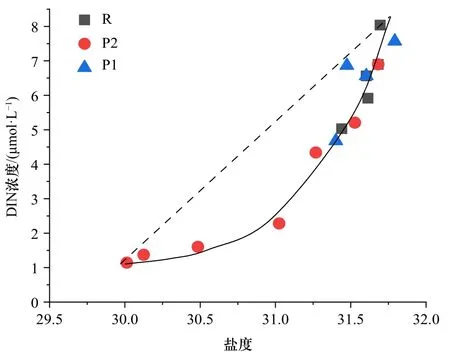
图7 溶解无机氮(DIN)浓度与盐度关系Fig. 7 Relationship between dissolved inorganic nitrate (DIN)concentration and salinity
4.3 浮游植物种群结构
在Ekman输送和地转力的共同作用下,BG上层的密度结构呈现中央低四周高的形态,营养盐浓度从外围向环流中心降低[16]。Zhuang等[31]也报道,从BG边缘到中心区域,以硅藻为主的生物量逐渐减少。这与我们在P断面观测到的营养盐浓度和藻类丰度从BG西边缘区域向中心减低的空间分布趋势相吻合。以P2断面东侧(8~10站位)为例,营养盐浓度的等密度线逐渐向下延伸,暖水ACW和冷水wBSW间的层化加剧,提高了氮限制对藻类生长的胁迫效应。营养盐浓度在真光层的降低,不利于浮游植物生长,导致Chla浓度从西向东逐渐降低(图3)。浮游植物粒径越小,在DIN限制下的适应能力越好[9,32],因而在BG影响范围内,以微微型浮游植物为主。
4.4 浮游植物的发展趋势
在北极变暖和北极放大效应的驱动下,北极生态环境趋向于向利于海洋浮游生物生存与繁衍的方向演变[33-34]。在近10年间,浮游植物的叶绿素a浓度年均增速约为0.013 mg/m3,初级生产力(以碳计)增速约为7 Tg/a[7]。BG海洋热含量在过去的30余年间增加1倍,加速了海冰消融,对生物地球化学过程产生了重要影响[35]。此外,浮游植物增加与营养盐供给增加密切相关。一方面,太平洋入流水持续增加,增加了sBSW携带的营养盐输入[36]。另一方面,融冰促进了阿拉斯加沿岸河流和陆源的有机物输入。BG淡水含量和洋流速度也稳步增加[37],比20世纪70年代升高了40%[38],进而增加了ACW向海盆区的营养盐供给[39-40]。以上原因都将缓解北极营养盐限制,推动浮游植物生物量的进一步提高[28]。
在本次调查的高密度冰区,我们发现叶绿素a浓度高于同地区的历史记录。刘子琳等[22]在2008年中国第三次北极考察时,测得楚科奇海北部区域的Chla浓度在0.004~0.468 mg/m3间变化。其最大值低于本次考察的Chla浓度最大值(0.69 mg/m3)。我们也对研究区域内2003-2016年间国际公开数据统计分析(航 次:32H120030911、32H120040910、49NZ20020-822、49NZ20040901、18SN20110721、18SN20120802、18SN20130724、18SN20140921、18SN20160922,https://explore.webodv.awi.de/ocean/hydrography/glodap/),发现该区域13年间的Chla浓度范围为0.025~0.54 mg/m3,平均浓度为(0.126±0.093)mg/m3。历史数据的Chla浓度最大值低于本次观测,平均值约为本次观测结果((0.327±0.163)mg/m3)的一半。该区域浮游植物的增长与北极圈内的变化趋势相一致[7]。从采样时间看,公开数据和刘梓琳等[22]的数据多在8-9月获取,本次调查采样早,冰覆盖下的藻华仍处于生长初期。预计随气温季节性升高,海冰融化,光限制削弱和无冰区扩大,浮游植物生物量将更高[1,41]。这意味着随着气候的变化,以浮游植物对碳吸收为主的生物碳汇将呈现增加的趋势,有利于北极作为碳汇区域的发展。
5 结论
(1)在楚科奇北部海区-加拿大海盆西侧海区,MW、ACW和sBSW等水团通过调控营养盐浓度和温度等因素,间接影响了浮游植物的分布和群落结构。
(2)叶绿素a浓度在40~50 m处的次表层暖水团(ACW、sBSW)中达到最大值,浓度在0.04~0.69 mg/m3之间,平均为(0.327±0.163)mg/m3。ACW相对于sBSW而言,水温高但营养盐浓度略低。由于浮游植物的环境适应性,在ACW中以粒径小于2 μm的微微型浮游植物为主(约65%),在sBSW影响的站位中,以粒径大于20 μm的小型浮游植物为主(约74%)。
(3)营养盐浓度、温度等因素对浮游植物的分布和群落结构影响较大。受北极极夜和冰下弱光影响,冰下浮游植物在长期演变中的暗适应性较强,光强对浮游植物分布的影响较弱。尽管融冰表层光照强,但寡营养盐和低温的表层冰融水会限制微藻生长。浮游植物在营养盐相对充裕和温度较高的ACW和sBSW暖水团附近(约50 m)聚集。根据保守混合模型和环境因子分析,叶绿素a浓度最大层中浮游植物的丰度和群落结构影响了DIN的吸收量和分布。
(4)浮游植物粒径越小,在DIN限制下的适应能力越好。在BG影响范围内,DIN限制明显,浮游植物多以微微型为主。
(5)在海冰融化初期,叶绿素a浓度高于以往的报道。这与北极变暖、融冰加剧、BG持续增强、太平洋入流水携带的营养盐增加等利于浮游生物生长的因素密切相关。浮游植物的增加有助于碳吸收,有望促进北冰洋碳汇增加。
