南大洋和南极考察航线海表超微型浮游生物分布及影响因子
2023-01-12罗光富何剑锋邵和宾曹叔楠蓝木盛张迪
罗光富 ,何剑锋 , ,邵和宾,曹叔楠,蓝木盛,张迪
( 1. 中国极地研究中心 南极长城极地生态国家野外科学观测研究站,上海 200136;2. 中国极地研究中心 自然资源部极地科学重点实验室,上海 200136;3. 同济大学 环境科学与工程学院,上海 200129)
1 引言
海洋超微型浮游生物(细胞粒径<3 μm)是海洋初级生产力的主要贡献者,广泛分布于近海和大洋水体中[1-2]。其主要类群包括:原绿球藻(Prochlorococcus,Pro)、聚球藻(Synechococcus,Syn)、超微型真核藻类(Pico-Eukaryote,PEuk)和异养浮游细菌(Heterotrophic Bacteria,HBac)。其中原绿球藻是迄今发现的地球上最小的放氧型光合自养原核生物,广泛分布在南北纬40°之间的大洋真光层中[3-4];聚球藻粒径略大于原绿球藻,呈球形或短棒状,其主要特征色素为藻胆蛋白,在全球大洋中均有发现[5-6];超微型真核藻类种类多样,主要色素为叶绿素a,广布于全球各大洋;海洋异养细菌是海洋中有机物的分解者,也是微食物环的重要组分,可利用海水中未被摄食消耗的有机碳进行生长和代谢[7]。海洋超微型浮游生物尽管个体微小,但数量巨大,对海洋生态系统的物质循环和能量流动有重要作用。
我国南极考察航线途经西太平洋、东印度洋,最后进入南大洋,形成纬向大断面,包含了从赤道往返极地、环绕南极洲和穿越西风带等重要断面。中国第6次南极考察期间,宁修仁等[8]应用落射荧光显微镜开展了我国首次环南极海域蓝细菌和微微型光合真核生物观测,证明了蓝细菌在极地海洋的存在。Wright等[9]利用高效液相色谱方法,也证实蓝细菌存在于南大洋。相较于传统的荧光显微检测等方法,流式细胞技术[10-12]可以快速有效地对海洋超微型浮游生物的几个类群进行同时检测,获取的数据稳定可靠,目前已在全球大洋(太平洋、大西洋、印度洋等)[3,5-6,13]以及近海(黄海、东海、南海等)[14-17]广泛使用。Lin等[18]和白晓歌[5]分别于中国第24次和第26次南极考察期间,使用流式细胞仪开展了环南大洋、西太平洋、印度洋等海域的超微型浮游生物丰度分布及其与环境因子相互关系研究。整体而言,目前关于超微型浮游生物在南极海洋中分布的相关资料和研究较少,由于海冰和西风带阻隔,只有极地科考破冰船和少量科考船有条件前往南大洋(尤其是60°S以南海域)采样,国内外现有数据主要集中于30°S以北海域,而30°S以南的环南极海域数据量非常少[19-20]。因此,通过每年南极考察逐年积累航线表层水海洋环境数据,对全面认识超微型浮游生物在全球不同纬度大洋中的分布和变化趋势具有重要意义。
本文利用中国第33次南极考察航次获取的去程、回程和环南极表层海水中超微型浮游生物及环境因子相关数据,分析航线上不同海域表层各类超微型浮游生物的丰度分布特征,阐述其丰度分布与环境因子的相关性,探讨其在南半球的分布界限,以期加深对超微型浮游生物全球分布现状的认识。
2 研究区域和方法
2.1 研究区域
2016年11月8日至2017年4月7日,中国第33次南极考察“雪龙”号航渡期间,利用船载表层海水采集系统获取了105个站位的表层海水样品,其中上海至南极中山站去程航段22站、环南极航段50站、自中山站返程航段33站。航线往返途经海域为东海、菲律宾以东海域、澳大利亚以西海域和南大洋(图1)。
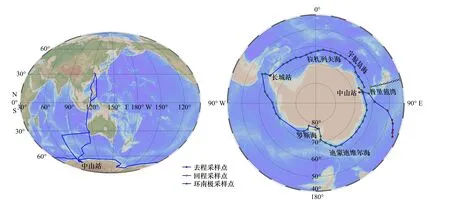
图1 中国第33次南极考察航迹及南大洋表层水采样站位(显示部分站位)Fig. 1 The 33rd Chinese National Antarctic Research Expedition route and sampling stations in the surface water of the Southern Ocean(show a part of the sites)
2.2 超微型浮游生物采样与分析
现场每个站位用棕色PEB瓶取海水100 mL,经50 μm孔径的筛绢预过滤后,取1 mL滤液用BD Accuri C6型流式细胞仪测定海洋超微型光合浮游生物丰度[5,18,21]。采集前向散射光(FSC)、侧向散射光(SSC)、橙色荧光(FL2,波长为(585±40)nm)和红色荧光(FL3,波长大于670 nm)信号强度。其中前向散射光信号表征细胞的大小和形状,侧向散射光信号表征细胞质的颗粒性,橙色荧光信号表征细胞中的藻胆蛋白含量,红色荧光信号表征细胞中的叶绿素含量,根据原绿球藻、聚球藻和超微型真核藻类上述信号的不同进行区分和计数。前向散射光、侧向散射光和红色荧光信号强度由大到小依次为超微型真核藻类、聚球藻、原绿球藻;橙色荧光信号强度聚球藻最大,原绿球藻和超微型真核藻类大约相等。
浮游细菌不含自发荧光的蛋白,需先染色后再上机检测。另取1 mL滤液染色,上机测定异养细菌丰度[5,11,22]。滤液加入染色剂SYBR Green-I 至终浓度为万分之一,混合均匀后在室温条件下避光染色 15 min,用上述流式细胞仪测定,采集绿色荧光(FL1,波长为(533±30)nm)和侧向散射光(SSC)信号强度,根据颗粒大小和绿色荧光信号强度对浮游细菌进行计数。
2.3 基础环境参数获取
温度和盐度数据由“雪龙”号表层海水采集系统配备的SBE 21 CTD 传感器直接测得。
采用萃取荧光法测定叶绿素a浓度[23-25]。每站采集1 000 mL水样,经20 μm孔径的筛绢、3 μm孔径的Nuclepore滤 膜 和0.7 μm孔 径 的Whatman GF/F玻 璃纤维滤膜逐级过滤。筛绢和滤膜在10 mL的90%丙酮中低温、避光萃取24 h,用Turner Designs Trilogy 实验室荧光仪测定,分别获得小型、微型和超微型光合浮游生物叶绿素a浓度。由于GF/F玻璃纤维滤膜可允许约15%的原绿球藻通过[11],因此原绿球藻数量较多的站位,其叶绿素a浓度可能比真实值偏低。
取100 mL水样,经GF/F玻璃纤维滤膜过滤,保存于经10%稀盐酸浸泡并用超纯水清洗过的白色PE瓶中,加入占水样体积2‰的三氯甲烷,置于4℃冰箱中低温保存。在国内实验室用营养盐自动分析仪(SKALAR SAN++)测定硝酸盐和亚硝酸盐、磷酸盐、硅酸盐浓度。
2.4 数据分析
使用Ocean Data View 软件绘制采样站位图,用Origin和ArcGIS软件绘制超微型浮游生物和环境参数分布图,用SPSS软件对数据进行统计分析和相关性分析。纬向上,由于去程和回程所走的航线并不完全一致,采样时间上也大致相差了4个月,因此本文将去程与回程的数据分开统计,可以体现数据的空间和季节性差异。在南大洋,由于航线经过的南极海洋环境多样,不同海域采样时间差异较大,所有环南极数据一起分析将会弱化特征数值,本文尝试将南大洋分3个扇面分析,以期减少采样时间段和空间位置对相关性分析的干扰。
3 结果
3.1 基础环境参数分布特征
纬向上,赤道南北两侧的表层海水温度基本呈对称分布(图2A),在赤道海域可超30℃,温度随纬度的增加而下降,南极海域温度降至接近冰点。盐度高值则出现在20°N附近和20°~40°S,最高值(35.76)出现在32°S附近,而回程赤道附近海域盐度最低只有30.46。纬向硝酸盐和亚硝酸盐浓度范围为0.02~32.75 μmol/L,平均值为(4.95±10.39)μmol/L;硅酸盐浓度范围为0.80~62.25 μmol/L,平均值为(8.28±13.73)μmol/L;磷酸盐浓度范围为0.01~1.80 μmol/L,平均值为(0.30±0.50)μmol/L。营养盐在40°S以南海域浓度较高,去程途中正值南半球的初夏(2016年11月底),光照加强,南大洋表层水中浮游植物大量生长,维持着较高的叶绿素浓度(0.13~1.57 mg/m3),表层水中硝酸盐和磷酸盐几乎被完全耗尽(59°S站位除外),硅酸盐浓度在47°S以北站位趋于耗尽,而在47°S以南迅速升高(最高值为62.25 μmol/L)。
环南极海域(图2a至图2e)的表层海水温度范围为-1.06~5.79℃,平均为(0.75±1.45)℃,高值主要出现在60°E附近的南极半岛海域。盐度范围为32.5~35.0,平均为33.65±0.52,高值出现在180°附近的罗斯海海域。环南极海域硝酸盐和亚硝酸盐浓度范围为0.26~35.43 μmol/L,平均值为(27.63±8.94)μmol/L;硅酸盐浓度范围为4.35~80.55 μmol/L,平均值为(54.85±17.64)μmol/L;磷酸盐浓度范围为0.09~2.07 μmol/L,平均值为(1.59±0.50)μmol/L。各项营养盐在南大洋海域的平均浓度均高于纬向的平均浓度,在罗斯海(170°E~160°W)出现了叶绿素a浓度次高值(0.33~1.27 mg/m3),此时正值南半球夏季(2017年2月),浮游植物大量繁殖,消耗了水体中的硝酸盐与磷酸盐,而硅酸盐浓度处于环南极海域较高水平。在40°~60°E有4个站位的硝酸盐与磷酸盐几乎被完全耗尽,对应的叶绿素a浓度约为0.50 mg/m3。

图2 不同海域表层温度、盐度、营养盐浓度纬向(A-E)和南大洋(a-e)分布Fig. 2 Distribution of surface temperature, salinity and nutrients concentration in different oceans along the latitudinal (A-E) and Southern Ocean (a-e)
3.2 超微型浮游生物的纬向分布特征
纬向上,超微型浮游生物的叶绿素a浓度(图3)范围在0.01~0.86 mg/m3,平均为(0.09±0.12)mg/m3。在43°S以北的热带和温带海域,超微型浮游生物可占总叶绿素a浓度的60%以上;而43°S以南的南大洋则降至占20%左右,但其叶绿素a浓度平均值仍高于热带和温带海域平均值。

图3 超微型、微型和小型浮游植物叶绿素a浓度纬向分布及分别占总叶绿素a浓度百分比Fig. 3 Distribution of chlorophyll a concentration of picoplankton, nanoplankton and microplankton along the latitudinal and the percentage of total chlorophyll a concentration respectively
超微型浮游生物各类群丰度纬向分布见图4。原绿球藻主要分布在40°S以北海域,丰度为0.12×103~48.64×103cells/mL,平均为(5.50±9.09)×103cells/mL。高值主要分布在10°N~10°S及20°~40°S之间,最高值出现在回程25°S附近。
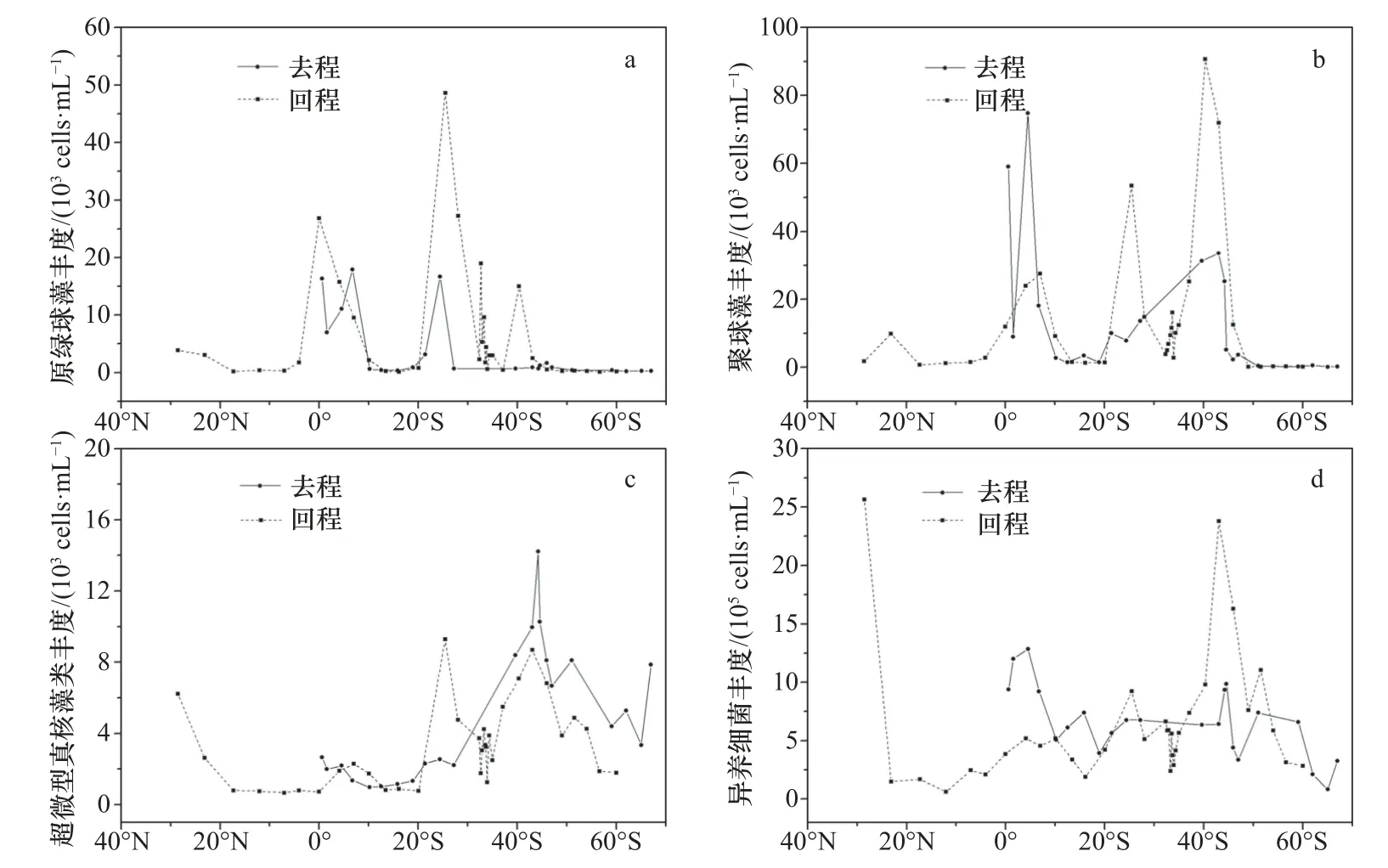
图4 超微型浮游生物丰度纬向表层分布Fig. 4 Latitudinal distribution of abundance of ultraplankton in the surface water
聚球藻主要分布在50°S以北海域,丰度范围在0.08×103~90.69×103cells/mL,平均为(13.56±20.33)×103cells/mL。高值主要集中在5°N~10°S及20°~50°S之间,最高值出现在回程途中的40°S附近。去程两个峰值分别出现在5°S和40°S附近,为74.73×103cells/mL
和33.56×103cells/mL。回程途中,分别在40°S、25°S和4°S出现明显峰值,并呈逐步降低趋势。
超微型真核藻类在各纬度的海域广泛存在,丰度范围为0.67×103~14.21×103cells/mL,平均为(3.87±3.08)×103cells/mL。去程与回程丰度的纬向分布曲线较为相似,由热带至温带逐步升高,在30°~65°S之间存在一个高值区,最高值(14.21×103cells/mL)出现在去程40°S附近,而后至南极近岸海域呈逐步降低趋势。回程的最高值出现在25°S附近,为9.28×103cells/mL,并在43°S附近出现次高值(8.68×103cells/mL)。
异养细菌丰度范围为0.61×105~25.65×105cells/mL,平均为(6.39±4.78)×105cells/mL。高值主要分布在30°N及30°~50°S之间,最高值出现在回程途中的30°N附近,并在43°S附近出现次高值23.78×105cells/mL。
3.3 环南极超微型浮游生物分布特征
在南大洋海域的表层水体中,超微型浮游生物的叶绿素a浓度(图5)范围为0.01~0.74 mg/m3,平均为(0.11±0.16)mg/m3,在拉扎列夫海、迪蒙迪维尔海、罗斯海西侧和南极半岛海域分别出现峰值。大部分站位超微型浮游生物叶绿素a浓度可占总叶绿素a浓度的15%~40%,在4个峰值附近可达60%。
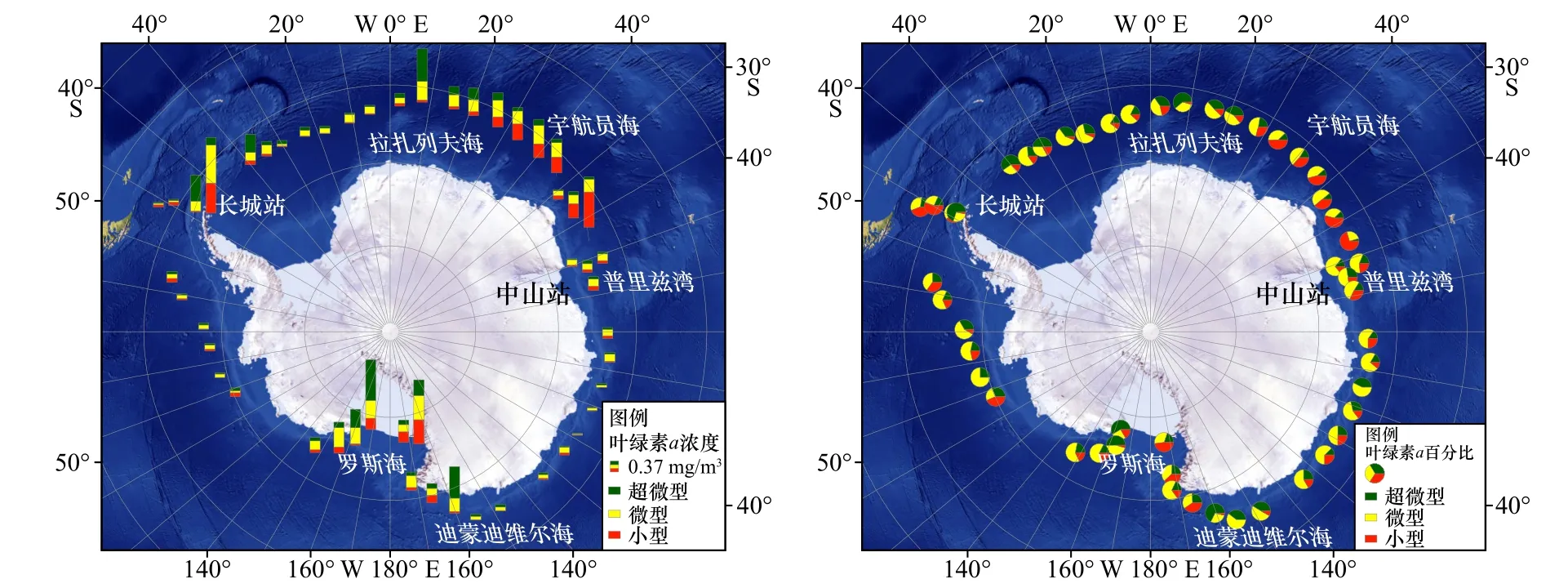
图5 超微型、微型和小型浮游植物叶绿素a浓度南大洋表层分布及占总叶绿素a浓度百分比Fig. 5 Distribution of chlorophyll a concentration of picoplankton, nanoplanktonin and microplankton in the surface water of the Southern Ocean and the percentage of total chlorophyll a concentration respectively
在丰度方面,大部分站位仅检测到超微型真核藻类和异养细菌,在少数站位检测到丰度较低的聚球藻(图6)。其中超微型真核藻类丰度范围在1.66×103~8.07×103cells/mL,平均为(3.31±1.46)×103cells/mL,在拉扎列夫海、迪蒙迪维尔海、罗斯海西侧和南极半岛海域出现峰值(丰度大于5×103cells/mL)。异养细菌的丰度范围为1.01×105~25.82×105cells/mL,平均为(4.68±4.39)×105cells/mL。在普里兹湾北部、罗斯海西侧和南极半岛海域分别出现峰值(丰度大于10×105cells/mL),其中罗斯海西侧和南极半岛海域两个站位的异养细菌丰度显著高于南大洋其他海域的丰度(可达4~5倍)。根据叶绿素a实测数据可推测,采样时段可能处于水华期,浮游植物大量生长繁殖促进异养细菌丰度改变。聚球藻的平均丰度为(0.38±0.39)×103cells/mL,其中南极半岛海域、普里兹湾和宇航员海3个站位的丰度大于1×103cells/mL。
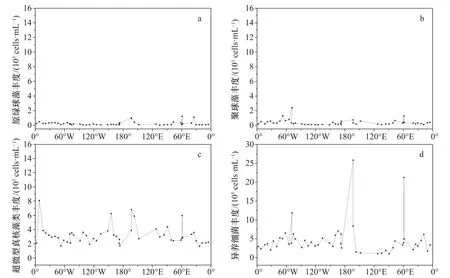
图6 超微型浮游生物丰度南大洋表层分布Fig. 6 Distribution of abundance of ultraplankton in the surface water of the Southern Ocean
3.4 超微型浮游生物丰度与环境因子的相关性
超微型浮游生物与环境因子的Pearson相关性分析结果(表1)显示:(1)纬向上,超微型真核藻类(PEuk)丰度、总叶绿素a浓度与温度呈极显著负相关(p<0.01,n=55);原绿球藻(Pro)丰度与温度呈显著正相关(p<0.05,n=55);4类超微型浮游生物丰度与盐度、营养盐浓度均无显著相关性;(2)南大洋海域,超微型浮游生物与营养盐浓度的相互作用密切。其中0°~120°E扇面(印度洋扇面,包含普里兹湾)总叶绿素a浓度与温度呈显著负相关(p<0.05,n=20);在120°E~120°W扇面(太平洋扇面,包含罗斯海)总叶绿素a浓度、聚球藻丰度和异养浮游细菌丰度与硝酸盐和亚硝酸盐浓度呈显著负相关(p<0.05,n=13),超微型真核藻类丰度与磷酸盐浓度呈负相关,总叶绿素a浓度与硅酸盐浓度呈显著正相关,说明罗斯海等海域的浮游植物主要利用硝酸盐和亚硝酸盐与磷酸盐;在120°W~0°扇面(主要为大西洋扇面,包含南极半岛海域)超微型真核藻类丰度、聚球藻丰度与硝酸盐和亚硝酸盐浓度呈显著负相关(p<0.05,n=17),聚球藻丰度与磷酸盐浓度呈极显著负相关(p<0.01,n=17)。
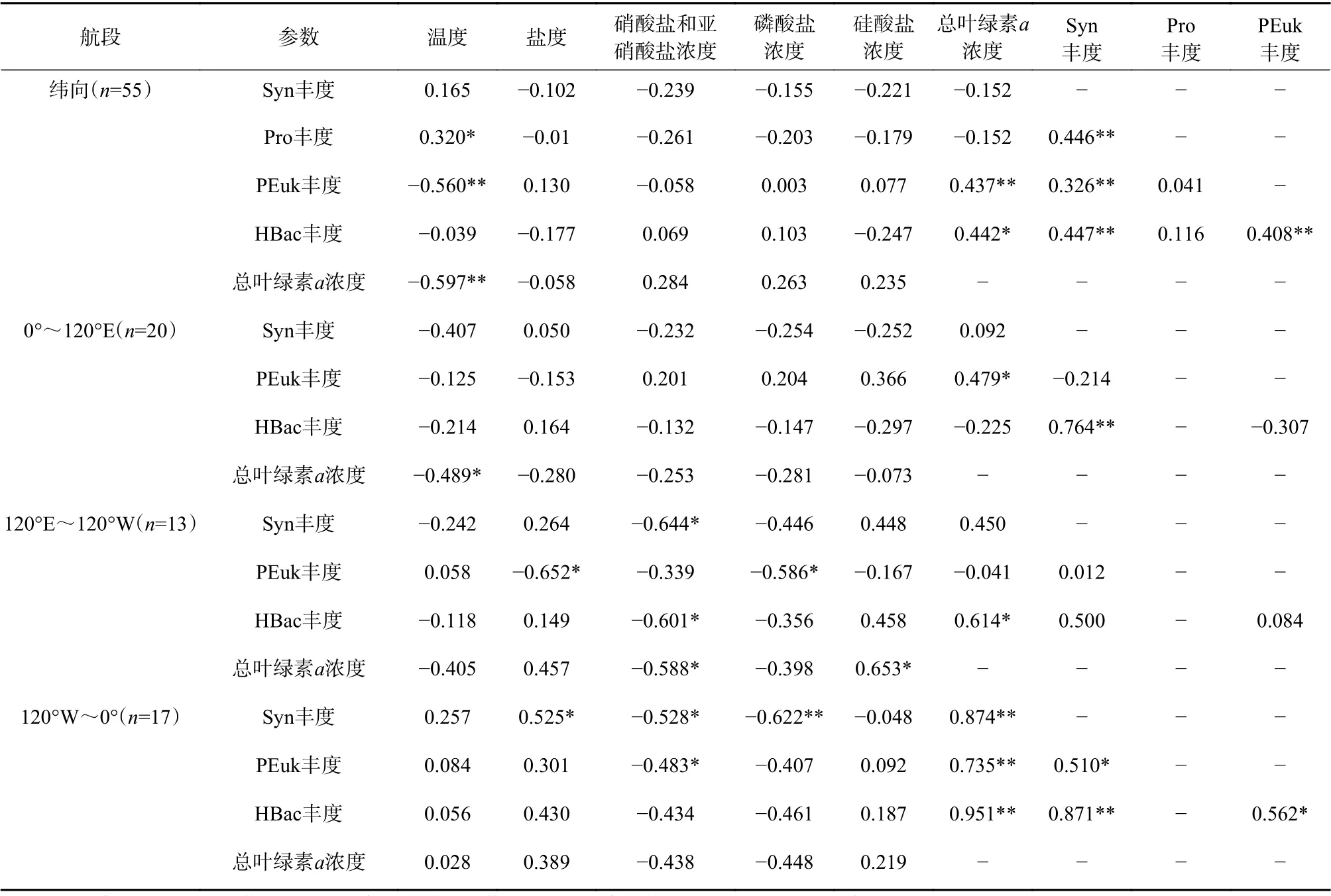
表1 超微型浮游生物丰度与相关环境因子的Pearson相关性分析Table 1 Pearson correlation analysis between ultraplankton abundance and related environmental parameters
4 讨论
4.1 超微型浮游生物的南半球分布界限
已有研究表明,原绿球藻广泛分布于40°N~40°S之间,水体温度小于15℃时会限制原绿球藻的分布[19,26],在北极或亚北极海域(45°N以北)可以被检测到[27],在西太平洋原绿球藻最南分布于50°S[5],Doolittle等[28]发现在水温低于10.65℃或者42°S以南海域不存在原绿球藻。本次考察主要在40°S以北的站位检测到原绿球藻,其中回程途中从43°S站位(水温13.8℃)开始检测到少量原绿球藻,在40°S站位(水温16.0℃)原绿球藻显著增加,在25°S附近(水温24.3℃)达到本航次最高值,继续往北原绿球藻丰度迅速降低并在赤道附近出现本航次的次高值。
聚球藻在极地或亚极地低温环境下也能够被检测到[29-30],河口或海岸型聚球藻在白令海和楚科奇海北部海域微微型浮游生物群落中占主导地位[31],丰度通常小于1×103cells/mL;在南极的近岸海域和湖泊中也能检测到聚球藻[32-33]。本航次在南大洋海域检测到低浓度的聚球藻,说明聚球藻在南半球分布界限可达60°S以南,平均值为(0.38±0.39)×103cells/mL,略高于宁修仁等[8]在1989年和1990年的调查结果,其中南极半岛海域、普里兹湾和宇航员海3个站位的丰度大于1×103cells/mL,总体上南大洋海域的聚球藻丰度平均值小于北极海域(表2)[27,34-35]。Walker和Marchant[33]报道了在南极普里兹湾冬季的沿岸水体和融冰区,尽管水温很低仍发现高丰度的蓝细菌,推测其来源可能是底栖种群随冰晶的再悬浮和夏季海冰消融的释放。本研究中在南大洋少量站位出现的聚球藻也有可能是底栖种群再悬浮和海冰的消融所导致。
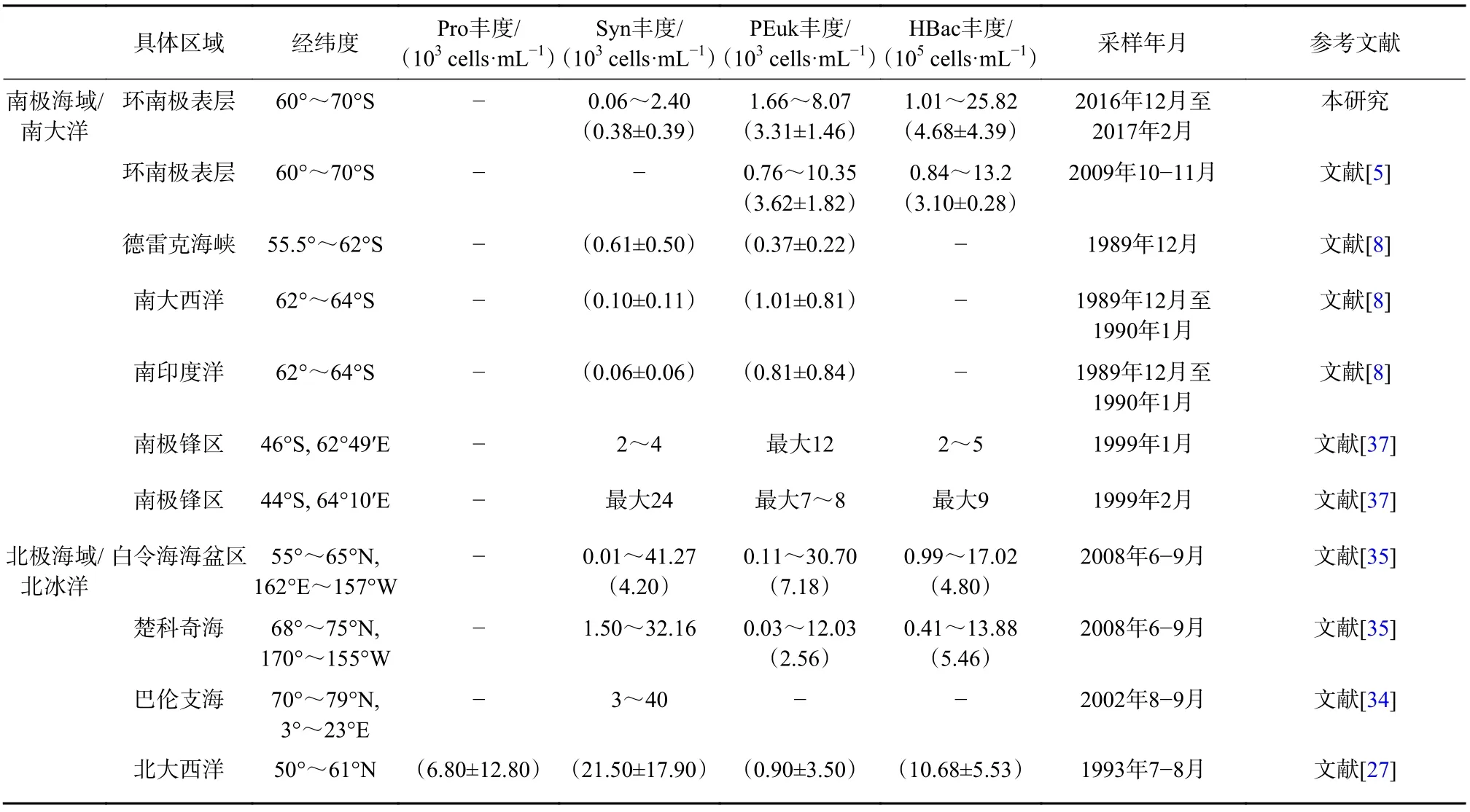
表2 南大洋和北冰洋中超微型浮游生物丰度Table 2 Abundance of ultraplankton in the Southern Ocean and the Arctic Ocean
值得注意的是,Flombaum等[19-20]基于全球上百个航次数据模拟的超微型浮游生物全球分布结果显示,原绿球藻、聚球藻和超微型真核藻类丰度沿赤道向两极随纬度逐步降低。而本航次数据显示,在10°~20°S(印度洋东北侧)表层存在一个超微型浮游生物低值区,纬向分布上呈现非连续和非线性特征,这一现象可能是因利文洋流(Leeuwin Current)引起的下降流,导致大洋洲西部海域大陆架处于寡营养状态[36]。因此,更多的实测数据可以不断提高模型的精度,有助于更全面认识全球不同海域超微型浮游生物的分布特征。
4.2 环境因子对超微型浮游生物的影响
温度是超微型浮游生物分布重要的调控因子[38-39]。在自然界中,温度是影响原绿球藻分布及其群落组成的重要因素[4],本研究中去程与回程航段的原绿球藻丰度与温度呈正相关,符合以上特征。聚球藻同样也受水温的影响显著,本研究中的聚球藻主要分布在50°S以北海域,对应水温为10.6℃,继续向南航线纬度升高水温降低,后续的南大洋表层水样品中仍有少量站位检测到聚球藻。基于全球尺度的生态位模型评估表明,如果未来海水温度上升,会导致原绿球藻和聚球藻丰度增加[19-20]。本研究中超微型真核藻类在纬向上的热带、温带、寒带的海域中均有分布,在纬向上其丰度与表层水温呈现显著负相关,超微型真核藻类的多样性非常高[6,40-41],且具有能够适应极端低温的物种(如球形棕囊藻Phaeocystis globosa Scherffel等),因此它在全球海洋中的分布范围比原绿球藻和聚球藻都要广,在水温8.5℃的海域其丰度达到最大[20]。整体而言,在纬向上温度是表层海水中超微型浮游生物种类和丰度的重要影响因子,不同水温的海洋环境下生活的超微型浮游生物种类和丰度各不相同。而在南大洋海域,表层水温的变化范围小,大部分超微型浮游生物属于适应低温环境的冷水种,温度对总叶绿素a浓度、超微型真核藻类和浮游细菌丰度的影响较小。
超微型浮游植物光合作用需要吸收多种营养物质,大部分营养盐物质在海水中的含量不会构成限制因子,但是其中的无机营养盐类(硝酸盐、磷酸盐、硅酸盐等)的浓度是影响初级生产力的重要因子[38-39]。另外,铁和锰等微量元素在某些海区的含量不足时也可限制初级生产,如南大洋海域受铁元素限制[42-44]。本航次在纬向40°S以北的中低纬度海域,表层水中的营养盐平均浓度低(其中硝酸盐和亚硝酸盐平均浓度小于1 μmol/L,磷酸盐平均浓度小于0.2 μmol/L,硅酸盐平均浓度小于5 μmol/L),总叶绿素a浓度也低,与印度洋南赤道流区[45]、热带太平洋西部[46]及北太平洋中部[47]的表层水相似,营养盐与总叶绿素a浓度、原绿球藻、聚球藻和超微型真核藻类丰度均没有明显的相关性。表层水营养盐浓度低可能与温跃层限制了高营养盐的底层水体向上补充有关[48],尽管营养盐浓度低,但是超微型浮游生物却可占总叶绿素a浓度的60%以上(图3),原绿球藻和聚球藻的丰度仍处于航次较高水平。原绿球藻和聚球藻尽管粒径小,但具有较大的比表面积,更善于利用海洋中有限的营养盐促进自身生长[6,49]。在南大洋海域,本航次获取的各项表层水营养盐浓度均远远高于中低纬度海域,可能是由于南大洋强烈的西风和湍流作用使其夏季的混合层可达100多米深[44],上层水层化很弱,下层水体中的营养盐可以及时向上补充。尽管南大洋的超微型浮游生物只占总叶绿素a浓度的20%左右,但其叶绿素浓度平均值仍高于热带和温带海域平均值(图3,图5)。在南大洋不同扇面,营养盐浓度与超微型浮游生物丰度的相关性也各不相同。在罗斯海所在的太平洋扇面,聚球藻和异养细菌丰度与硝酸盐和亚硝酸盐浓度呈显著负相关,而超微型真核藻类丰度与磷酸盐浓度呈显著负相关;在大西洋扇面,超微型真核藻类、聚球藻丰度与硝酸盐和亚硝酸盐浓度呈显著负相关,而聚球藻丰度与磷酸盐浓度呈显著负相关;而印度洋扇面,超微型浮游生物丰度与各项营养盐浓度的相关性不明显,这可能与超微型浮游生物占浮游植物总量的比例较低有关(图5)。
总体而言,不同海域的表层海水环境特点会影响该海域超微型浮游生物的丰度、分布、生理特征等,同时该海域的超微型浮游生物及其食物链其他生物的生命活动也会影响和改变水体的环境特征。
5 结论
中国第33次南极考察期间,在“雪龙”号往返南极和环南极航行的途中,对各海域表层海水中的超微型浮游生物丰度和相关环境因子进行了观测和采样,通过对后续数据分析和研究,获得以下结论:
(1)在热带与温带海域,超微型浮游生物可占总叶绿素a浓度的60%以上,在南大洋则集中在15%~40%。原绿球藻主要分布在40°S以北海域,聚球藻主要分布在50°S以北海域,超微型真核藻类和异养细菌在航线经过的各海域均有存在。原绿球藻的平均丰度为(5.50±9.09)×103cells/mL;聚球藻的平均丰度为(13.56±20.33)×103cells/mL;超微型真核藻类的平均丰度为(3.87±3.08)×103cells/mL;异养细菌的平均丰度为(6.39±4.78)×105cells/mL。
(2)在南大洋海域的表层水体中,大部分站位仅检测到超微型真核藻类和异养细菌,超微型真核藻类的平均丰度为(3.31±1.46)×103cells/mL;异养细菌的平均丰度为(4.68±4.39)×105cells/mL;在少数站位检测到较低丰度的聚球藻,平均值为(0.38±0.39)×103cells/mL。
(3)在纬向上,温度对超微型浮游生物的影响显著,超微型真核藻类丰度与温度呈极显著负相关,原绿球藻丰度与温度呈显著正相关,盐度与营养盐浓度对其影响较弱;在南大洋海域,超微型浮游生物丰度与营养盐浓度的相互作用密切,在南极半岛海域浮游植物大量消耗硝酸盐、磷酸盐和硅酸盐,而在罗斯海则主要利用硝酸盐与磷酸盐。
致谢:衷心感谢自然资源部第二海洋研究所韩正兵博士、范高晶博士协助营养盐样品分析!感谢“雪龙”号全体船员对本文样品采集提供支持!同时感谢审稿专家提出的宝贵意见!
