渤海和南黄海沉积物中氨氧化微生物对硝化潜势的相对贡献
2023-01-12李明月甄毓李思琦米铁柱贺惠
李明月,甄毓 ,李思琦 ,米铁柱 ,贺惠
( 1. 山东理工大学 资源与环境工程学院,山东 淄博 255049;2. 青岛海洋科学技术试点国家实验室 海洋生态与环境科学功能实验室,山东 青岛 266071;3. 中国海洋大学 海洋环境与生态教育部重点实验室,山东 青岛 266100;4. 中国海洋大学海洋生命学院,山东 青岛 266003)
1 引言
全球氮超载是21世纪一个主要的环境问题,人类活动来源的大部分氮通过河流、地下水和大气进入沿海海洋系统[1]。硝化作用与反硝化或厌氧氨氧化作用相结合,是生态系统中氮去除的重要途径,而氮的去除很大程度上依赖于硝化作用的产物—亚硝酸盐和硝酸盐[2-3]。因此,研究近海生态系统中硝化作用的能力具有重要意义。硝化作用包括铵/氨转化为亚硝酸盐的氨氧化过程和亚硝酸盐转化为硝酸盐的过程。氨氧化过程被认为是硝化作用的限速步骤,因为环境中很少会有亚硝酸盐的大量积累[4]。因此,氨氧化能力通常用来表征硝化能力。潜在硝化速率是指在不受底物( NH+4/NH3)限制情况下的硝化速率,在最优条件下,向培养体系中添加足量的铵,并通过亚硝酸盐和硝酸盐生成量来计算潜在硝化速率[5],潜在硝化速率可用于量化硝化潜势[6]。
氨氧化过程主要由氨氧化古菌(Ammonia-Oxidizing Archaea, AOA)和氨氧化细菌(Ammonia-Oxidizing Bacteria, AOB)来驱动,在AOA被发现之前[7-9],人们一直认为AOB是氨氧化过程的唯一贡献者。随后,AOA被证明在多种环境中普遍存在,例如湖泊[10]、湿地[11]、土壤[12]、污水厂[13]、河口[14-16]和海洋系统[17-19]等。而且,研究表明,与AOB相比,AOA在许多环境(例如农田土壤和海湾沉积物)中都占有优势[12,15,19]。但是,在某些环境(例如湖泊沉积物和河口沉积物)中则是AOB占优势[10,16]。结合现有研究报道,有学者认为,AOA在氮限制、低氧和酸性环境中更容易占有优势[20],但迄今,科学家们并未在它们对硝化过程的相对贡献上达成共识。因此,AOA和AOB对氨氧化过程的相对贡献成为氮循环中最重要的热点问题之一。
在海洋沉积物中,AOA和AOB的相对贡献研究多基于分子生态学的方法,利用AOA和AOB氨单加氧酶基因(amoA)的丰度来比较AOA和AOB的重要性[17,21]。但是,首先,基于DNA功能基因丰度的研究不能代表AOA和AOB实际发挥功能的微生物数量;其次,有研究表明,AOB平均每个细胞的硝化潜势是AOA的1~3倍[22];最后,AOA和AOB功能基因丰度和硝化速率的准确关系并不明确;因此,AOA或AOB功能基因丰度不能直接代表其对氨氧化过程或硝化过程贡献的多少。研究者开始结合潜在硝化速率的测定来进一步探讨AOA和AOB对硝化过程的贡献,但是,大部分研究都是直接测定总潜在硝化速率,通过AOA和AOB丰度与总潜在硝化速率的相关性来区分其对硝化作用的相对贡献,但效果欠佳[10,15,23-25]。近几年来,有研究者开始使用抗生素、炔烃类抑制剂和传统硝化抑制剂等来抑制AOA或AOB,用培养实验直接表征AOA和AOB的硝化潜势,从而揭示其对硝化作用的相对贡献[16,19,26-27]。然而,海洋沉积物中的相关研究较少,更缺乏跨海域大面积的研究。综上,基于抑制剂区分AOA和AOB的硝化潜势,用培养实验直接表征海洋沉积物中AOA和AOB对硝化作用相对贡献的研究十分必要。
渤海和黄海为典型半封闭型陆架边缘海,沿岸河网交错、人类活动区域密集,而且,黄、渤海水体停留时间较长[28-29],因此,人类活动对其氮负荷等的影响可能比其他开放水域更为深远。本研究以渤海和南黄海海域为研究区域,以表层沉积物为研究对象,通过室内模拟培养实验,结合环境因子数据,直观地研究目标海域内AOA和AOB对硝化潜势的相对贡献。本研究对于阐明AOA和AOB对硝化作用的贡献率具有重要意义,可为丰富近海氮元素的生物地球化学循环理论提供依据。
2 材料与方法
2.1 样品采集
2019年5月2-10日搭乘“中渔科102”号调查船、5月4日搭乘“北 斗”号 调 查 船 于 渤 海(294、1251、3562、5262、5094、6251、B22和B24站位)进行样品采集;2019年4月12-17日搭乘“北斗”号调查船于南黄海(H6、H7、H9、H12、H16、H18、H20、H22、H27、H30、H31、H34站位)进行样品采集(图1)。现场采集各站位沉积物样品和底层水样品,表层沉积物(1~3 cm)用箱式采泥器采集并充分混合后置于无菌封口袋内并立即储存于4℃冰箱,带回实验室后立即进行培养实验。底层水样品(5 L)通过船载CTD多参数温盐深仪进行采集,经0.45 μm醋酸纤维滤膜过滤后,水样分为两份,一份储存于-20℃冰箱以测定氨盐( NH+4)、亚硝酸盐( NO-2)、硝酸盐( NO-3)和磷酸盐( P O34-)浓度,另一份加入氯仿后常温保存以测定硅酸盐( S iO23-)浓度[30]。
图1 渤海和南黄海海域采样站位Fig. 1 Sampling sites in the Bohai Sea and South Yellow Sea
2.2 环境因子测定
各站位底层水温度、盐度、电导率、溶解氧浓度和pH等理化参数通过船载CTD多参数温盐深仪进行原位测定。底层水样带回实验室后,其 NH+4、 NO-2、和 SiO23-等营养盐参数用Quaatro营养盐自动分析仪(Seal Analytical Ltd.)测定[30]。
2.3 潜在硝化速率培养实验
潜在硝化速率测定参考Bernhard等[23]和Zheng等[16]的方法。简要步骤如下:取1.0 g沉积物(湿重)加入含30 mL海水(经0.22 μm滤膜过滤并灭菌)的100 mL玻璃锥形瓶中;在锥形瓶中加入终浓度为300 μmol/L的NH4Cl和60 μmol/L的KH2PO4;将锥形瓶放置在震荡培养箱中,以现场温度、120 r/min在黑暗条件下连续震荡培养;在培养期间的0 h、24 h、48 h和72 h进行取样。样品经离心、过滤后立刻放入-20℃冻存,以测定 NO-2和 NO-3浓度。为了区分AOA和AOB对硝化潜势的相对贡献,每个站位设置两个实验组,其中一个实验组加入1.0 g/L的氨苄青霉素。作为一种β-内酰胺抗生素,氨苄青霉素能在细胞生长过程中作用于细菌细胞壁,抑制细菌的生长,但不会对古菌产生影响[31]。因此,不添加氨苄青霉素的实验组可表征AOA和AOB的总硝化潜势,添加氨苄青霉素的实验组可表征AOA的硝化潜势。所有实验组设置3个平行培养体系。
2.4 潜在硝化速率计算
潜在硝化速率根据每克沉积物中 NO-2和 NO-3含量随时间的变化进行计算。通过0 h、24 h、48 h和72 h时 NO-2和 NO-3积累量进行线性回归分析,回归方程的斜率即为相应培养体系的潜在硝化速率。每个站位的总潜在硝化速率通过未添加氨苄青霉素的实验组进行计算,AOA的潜在硝化速率通过添加氨苄青霉素的实验组进行计算,AOB的潜在硝化速率则为两者之差。
2.5 统计分析
采用曼-惠特尼(Mann-Whitney)检验分析两个独立样本之间的差异,皮尔森(Pearson)相关系数法分析两组变量之间的相关性,曼-惠特尼检验、皮尔森相关性分析和潜在硝化速率回归分析均通过R v4.0.3软件实现。
使用R v4.0.3中的vegan软件包对潜在硝化速率和环境因子数据执行约束性排序分析,分析环境因子对硝化潜势的影响。首先通过去趋势对应分析(Detrended Correspondence Analysis, DCA)选择线性排序冗余分析(Redundancy Analysis, RDA),然后通过蒙特卡罗置换检验(置换次数为999)验证环境因子对潜在硝化速率分布解释量的显著性,并通过函数envfit检验每个环境因子对潜在硝化速率影响的显著性。当环境因子数量较多时,通过BIOENV分析挑选最佳环境因子组合后,再进行约束性排序分析。
3 结果与分析
3.1 环境因子分析
各站位环境因子参数如表1所示,在目标海域内,站位水深范围为13.0~81.0 m,除H27站位水深最浅外,南黄海所有站位深度均大于渤海,渤海和南黄海海域水深具有显著差异(曼-惠特尼检验,p<0.01)。站位温度范围为8.50~12.30℃,渤海和南黄海海域底层水温度无显著性差异(p>0.05)。站位盐度范围为29.82~33.44,与深度具有显著正相关性(皮尔森相关性,r=0.653,p<0.01),渤海站位盐度显著低于南黄海站位盐度(p<0.01)。站位电导率范围为33.54~37.38 mS/cm,与温度和盐度具有显著正相关性(r=0.588,p<0.01;r=0.731,p<0.001),渤海站位电导率显著低于南黄海站位(p<0.01)。站位溶解氧浓度范围为7.81~9.85 mg/L,与温度具有显著负相关性(r=-0.539,p<0.05),渤海和南黄海海域底层水溶解氧浓度无显著性差异(p>0.05)。站位pH范围为8.17~8.63,与深度和盐度具有显著负相关性(r=-0.587,p<0.01;r=-0.581,p<0.01),渤海站位pH显著高于南黄海站位(p<0.001)。总体来说,渤海海域水深、盐度、电导率均显著小于南黄海,pH显著高于南黄海,而温度和溶解氧浓度则无显著差异。

表1 渤海和南黄海海域采样站位底层水理化参数Table 1 Physiochemical parameters of bottom water in sampling sites of Bohai Sea and South Yellow Sea
整体来看,渤海海域营养盐参数均小于南黄海海域,其中, N O-2浓 度、浓度和 S iO23-浓度具有显著差异(p<0.05)。具体来说, N H+4浓度为2.393~6.105 μmol/L,浓度较高的站位为H6、H18和H16(表1)。 NO-2浓度和 NO-3浓度分 别为0.064~0.713 μmol/L和0.215~13.366 μmol/L, NO-2和 NO-3浓 度 较 低 的 站 位 均 为1251和H9, NO-2浓度最高的站位为H20, NO-3浓度较高的站位依次为5094、H30和H27。 PO34-浓度为0.028~0.937 μmol/L, SiO23-浓度为1.253~16.277 μmol/L,浓度最高的站位均为H30。除 NO-3外,水深和营养盐参数具有显著的正相关关系(r>0.593,p<0.01),而温度对营养盐参数无显著影响(p>0.05)。此外, NO-2浓度与盐度和电导率具有显著的正相关关系(r>0.447,p<0.05),而与溶解氧浓度具有显著的负相关关系(r=-0.511,p<0.05); PO34-浓度与盐度和电导率也具有显著的正相关关系(r>0.570,p<0.01),而与pH具有显著的负相关关系(r=-0.954,p<0.001); SiO23-浓度与pH具有显著的负相关关系(r=-0.657,p<0.01)。在各个营养盐参数之间, NO-2浓 度与 NH+4、和 SiO23-浓度具有显著的正相关关系(r>0.447,p<0.05);浓度与 NO-3和 PO34-浓度具有显著的正相关关系(r>0.608,p<0.01)。
3.2 培养过程中 N O-2 浓度和 N O-3浓度的变化
图2和图3展示了各站位表层沉积物培养体系(含1 g沉积物和30 mL海水)中 NO-2和 NO-3随时间的积累量(以氮计,下同),在不受 NH+4限制的情况下,所有培养体系中 NO-2和 NO-3浓度均持续上升,表明硝化过程活跃。其中,未添加氨苄青霉素的培养体系代表AOA和AOB共同驱动下 NO-2和 NO-3积累量随时间的变化情况,添加氨苄青霉素的培养体系代表AOA驱动下 NO-2和 NO-3积累量随时间的变化情况。结果表明,氨苄青霉素的添加能很好地抑制细菌的生长,以此区分细菌和古菌对硝化潜势的相对贡献(图2,图3)。
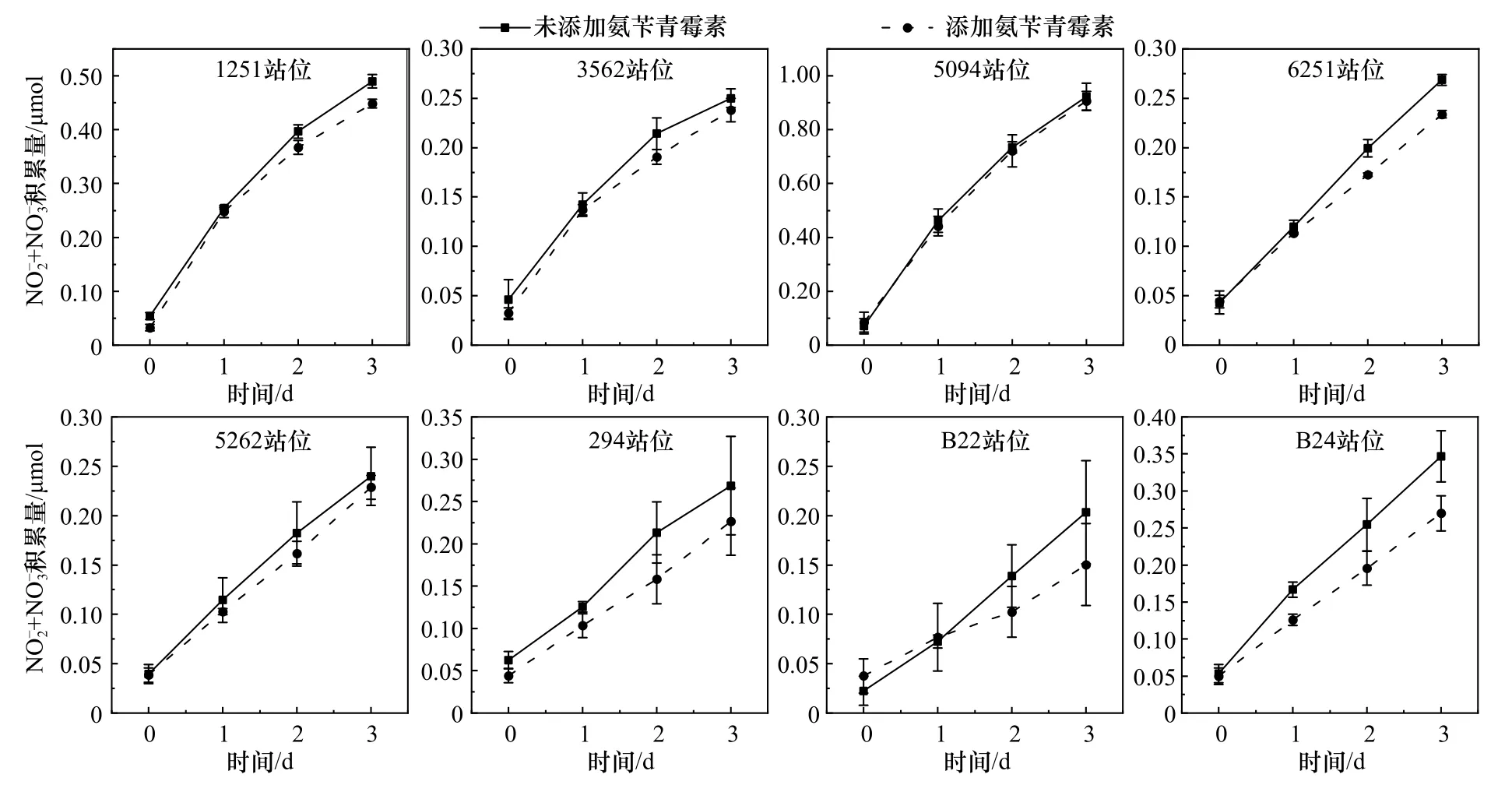
图2 渤海海域各站位表层沉积物样品培养体系中亚硝酸盐和硝酸盐积累量Fig. 2 The accumulation of nitrite and nitrate during the cultures of surface sediment samples from Bohai Sea
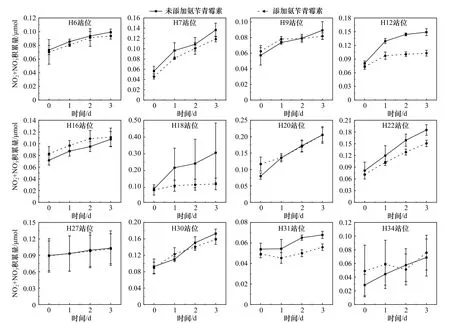
图3 南黄海海域各站位表层沉积物样品培养体系中亚硝酸盐和硝酸盐积累量Fig. 3 The accumulation of nitrite and nitrate during the cultures of surface sediment samples from South Yellow Sea
3.3 潜在硝化速率的分布特征
渤海和南黄海海域表层沉积物中潜在硝化速率如图4所示,渤海海域表层沉积物总潜在硝化速率(以氮计,下同)为0.060 9~0.283 1 μmol/(g·d),其中AOA潜 在 硝 化 速 率 为0.036 4~0.274 3 μmol/(g·d),AOB潜在硝化速率为0.001 4~0.024 5 μmol/(g·d);南黄海海域表层沉积物总潜在硝化速率为0.004 6~0.068 7 μmol/(g·d),其中AOA潜在硝化速率为0.004 3~0.030 2 μmol/(g·d),AOB潜 在 硝 化 速 率 为0.000 4~0.056 0 μmol/(g·d)。渤海海域总潜在硝化速率显著高于南黄海海域(曼-惠特尼检验,p<0.001),AOA潜在硝化速率也显著高于南黄海海域(p<0.001),但AOB潜在硝化速率与南黄海海域无显著差异(p>0.05)。

图4 渤海和南黄海海域表层沉积物中潜在硝化速率Fig. 4 The potential nitrification rates in surface sediments of Bohai Sea and South Yellow Sea
图5展示了总潜在硝化速率、AOA和AOB潜在硝化速率在渤海和南黄海海域的分布情况。在渤海海域,总潜在硝化速率、AOA潜在硝化速率均在5094和1251站位具有较大值,远高于其他站位;AOB潜在硝化速率则在B22、B24和294站位出现较大值(图4,图5)。5094和1251站位离海岸较近,B22和B24站位离海岸较远,因此,推测在渤海海域近海岸硝化潜势高于远海岸,AOA在离岸较近的地方更活跃,AOB在离岸较远的地方更为活跃(图5)。南黄海海域未表现出上述规律,H27站位总潜在硝化速率、AOA和AOB潜在硝化速率均呈现出最低值,说明该站位硝化作用较弱;H18站位总潜在硝化速率和AOB潜在硝化速率均表现出最高值,但是其平行培养体系间差别较大(图4),若去除一个极大值,则H20和H22站位表现出较高的潜在硝化速率(图4,图5)。
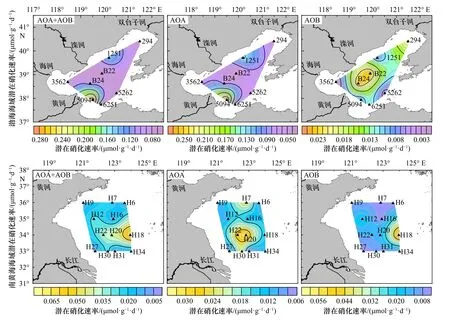
图5 渤海和南黄海海域表层沉积物潜在硝化速率分布Fig. 5 The distribution of potential nitrification rates in surface sediments of Bohai Sea and South Yellow Sea
3.4 氨氧化古菌和氨氧化细菌对硝化潜势的相对贡献
无论是总体分析,还是各海域单独分析,AOA对硝化潜势的贡献都显著高于AOB(曼-惠特尼检验,p<0.05)(图6),特别是渤海海域,AOA对硝化潜势的贡献远高于AOB(p<0.001),贡献为59.79%~97.95%,AOA和AOB硝化潜势的比值为1.49~47.82;南黄海海域则分别为18.47%~94.26%和0.22~16.41。对于渤海海域,3562、5094和1251站位AOA对硝化潜势的贡献率较大,且均离海岸较近(图6)。在南黄海海域,AOA对硝化潜势贡献率较大的站位依次为H7、H27和H6,而H12和H18站位则是AOB对硝化潜势的贡献高于AOA(图6)。
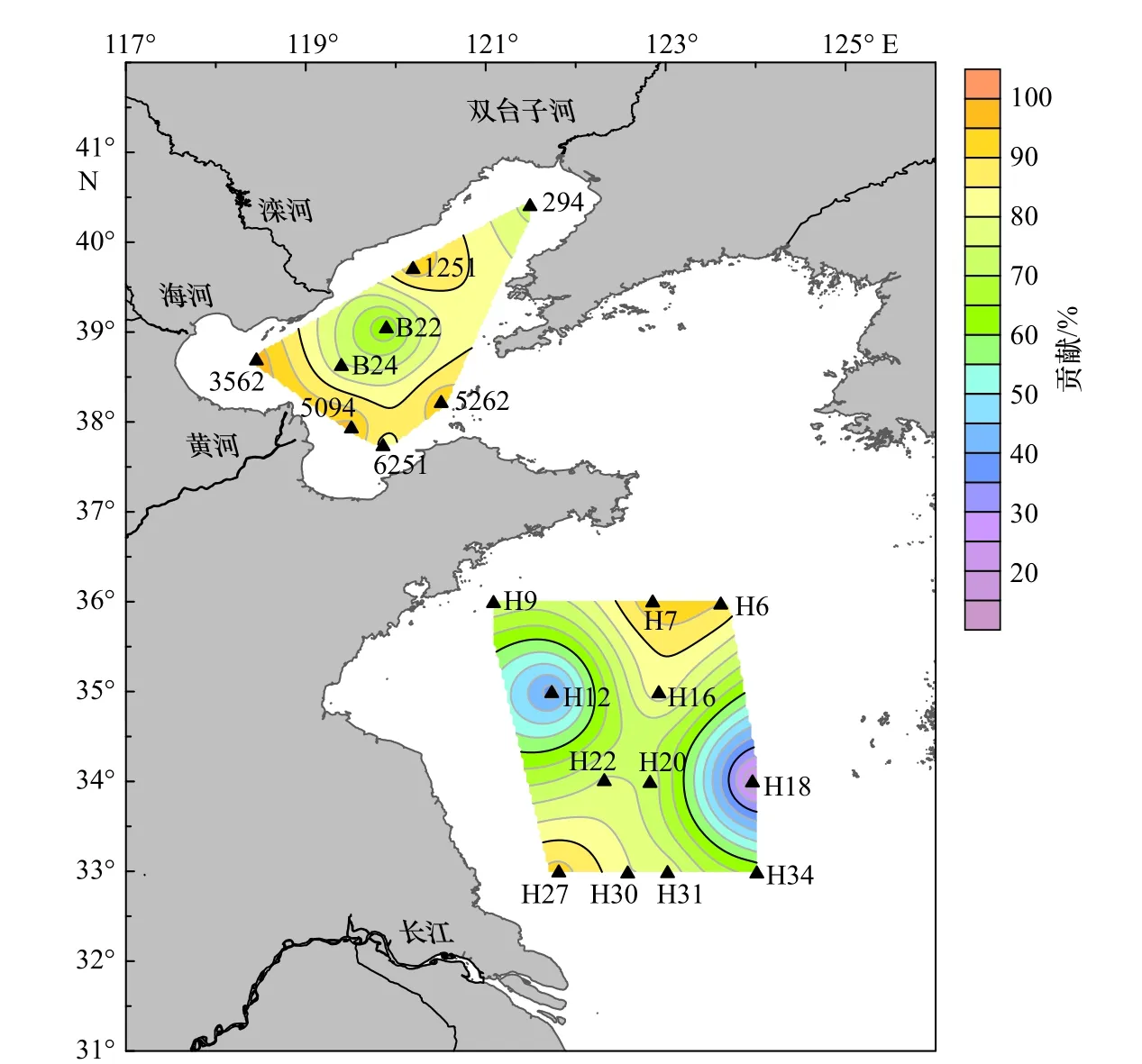
图6 渤海和南黄海海域表层沉积物中AOA对硝化潜势贡献的分布Fig. 6 The distribution of the contributions of AOA to nitrification potential in surface sediments of Bohai Sea and South Yellow Sea
3.5 环境因子对氨氧化古菌和氨氧化细菌硝化潜势的影响
DCA结果表明,RDA更适合对所有站位的潜在硝化速率和环境因子进行约束性排序分析(图7a)。两个排序轴的解释量分别为80.34%和1.41%,结果同样表明,渤海海域站位和南黄海海域站位潜在硝化速率分布具有明显差异(图7a)。蒙特卡洛全模型置换检验结果表明环境因子对潜在硝化速率的解释量不具有显著性(p>0.05),但是,盐度(p<0.05)、电导率(p<0.05)、pH(p<0.01)和 PO34-浓度(p<0.01)显著影响渤海和南黄海海域潜在硝化速率的分布。其中,pH对渤海各站位的潜在硝化速率分布影响大于南黄海,盐度、电导率和 PO34-浓度则对南黄海海域的潜在硝化速率影响更大,因此,不同海域影响硝化潜势的关键环境因子可能存在差异,这一现象有不少报道,例如,在不同区域潮间带沉积物中,影响潜在硝化速率的环境因子也存在差异[16,32]。皮尔森相关性分析表明,总潜在硝化速率和AOA潜在硝化速率与pH呈正相关关系,与水深、盐度和电导率呈负相关关系(图7b,|r|>0.495,p<0.05)。前文差异分析也表明,渤海海域总潜在硝化速率、AOA潜在硝化速率和pH显著高于南黄海海域,深度、盐度和电导率则显著低于南黄海海域。此外,总潜在硝化速率和AOA潜在硝化速率与PO34-浓度、总潜在硝化速率与 S iO23-浓度也呈现出显著的负相关关系(|r|>0.454,p<0.05)。

图7 潜在硝化速率和环境因子RDA和皮尔森相关性分析Fig. 7 RDA and Pearson correlation analysis of potential nitrification rates and environmental parameters
对渤海8个站位潜在硝化速率和环境因子进行分析,通过BIOENV筛选出6个环境因子,即盐度、电导率、pH、 NH+4浓 度、浓度和 SiO23-浓度,进行约束性排序分析。RDA分析结果如图7c所示,两个排序轴的解释量分别为97.91%和0.85%,蒙特卡洛置换检验结果表明,上述环境因子对潜在硝化速率的解释量不具有显著性,且单个环境因子的影响也不具有显著性(p>0.05)。皮尔森相关性分析表明(图7d),总潜在硝化速率和AOA潜在硝化速率与 NO-3浓度呈正相关关系(r>0.798,p<0.05),例如,在5094站位, NO-3浓度高于其他站位(表1),硝化潜势也高于其他站位(图4)。此外,AOB潜在硝化速率与pH具有显著的负相关关系(r=-0.772,p<0.05),而与 PO34-浓度具有显著的正相关关系(r=0.807,p<0.05)。
对南黄海12个站位潜在硝化速率和环境因子进行分析,通过BIOENV筛选出6个环境因子,即盐度、溶解氧浓度、pH、 NH+4浓度、 P浓度和 SiO23-浓度,进行约束性排序分析。RDA结果如图7e所示,两个排序轴的解释量分别为52.58%和11.45%,蒙特卡洛置换检验结果表明,上述环境因子对潜在硝化速率的解释量不具有显著性,且单个环境因子的影响也不具有显著性(p>0.05)。皮尔森相关性分析表明(图7f),总潜在硝化速率与盐度具有显著的正相关关系(r=0.606,p<0.05),总潜在硝化速率和AOB潜在硝化速率与溶解氧浓度具有显著的负相关关系(|r|>0.662,p<0.05),而AOA潜在硝化速率与 NO-2浓度具有显著的正相关关系(r=0.589,p<0.05)。
黄、渤海20个站位AOA和AOB对硝化潜势的相对贡献与环境因子均不存在显著相关性(皮尔森相关性,p>0.05),AOA和AOB硝化潜势的比值与盐度呈显著负相关关系(r=-0.481,p<0.05),与pH呈显著正相关关系(r=0.562,p<0.01),与 PO34-浓度呈显著负相关关系(r=-0.534,p<0.05)(图8a)。渤海8个站位AOA对硝化潜势的相对贡献与pH呈显著正相关关系(r=0.723,p<0.05),与P O34-浓度呈显著负相关关系(r=-0.792,p<0.05),AOB对硝化潜势的相对贡献与pH和 PO34-浓度则表现出相反的相关关系(图8b)。此外,AOA和AOB硝化潜势的比值与 SiO23-浓度呈显著正相关关系(r=0.768,p<0.05)(图8b)。南黄海海域AOA和AOB对硝化潜势的相对贡献以及AOA和AOB硝化潜势的比值与环境因子均不存在显著相关性(p>0.05,图8c)。
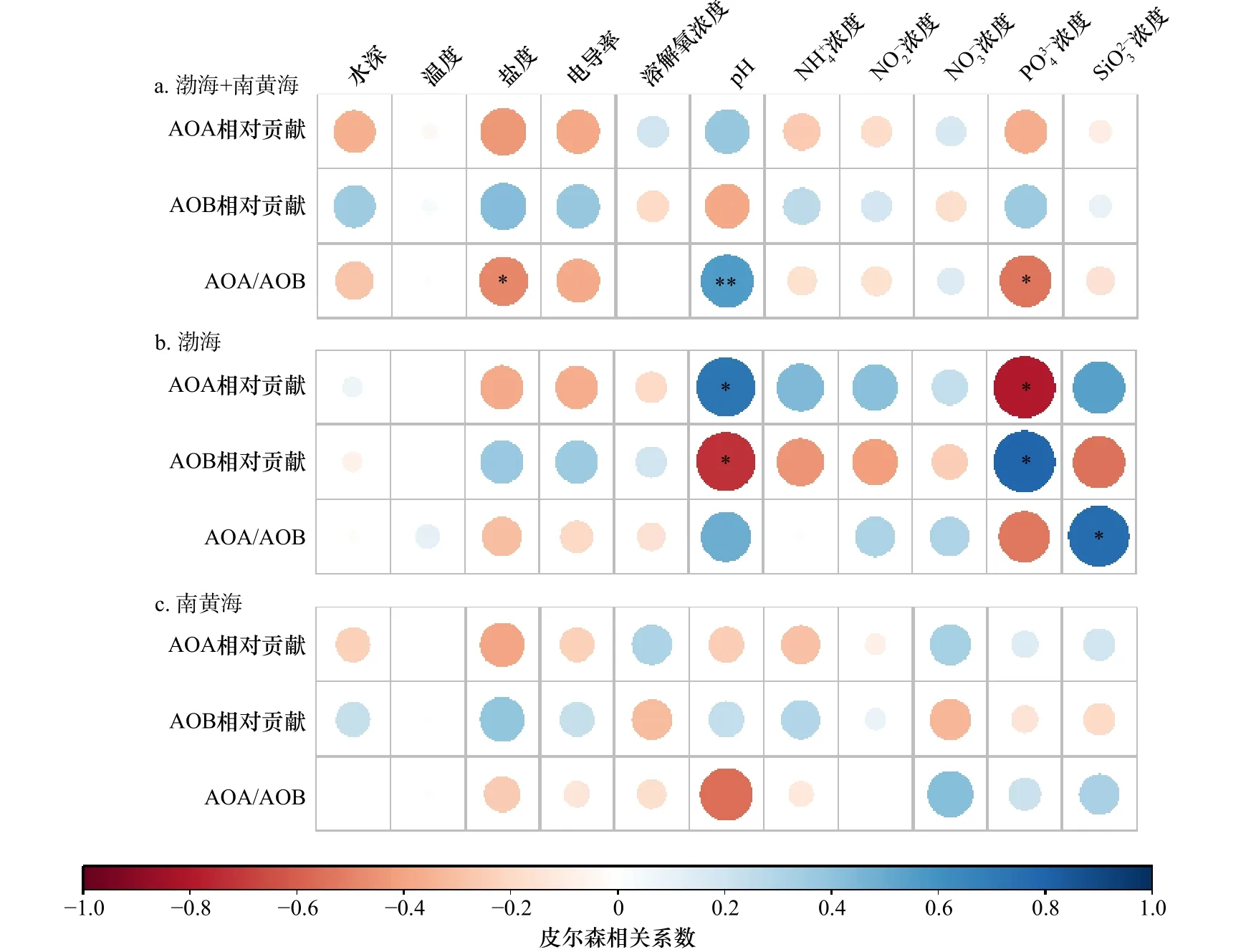
图8 AOA和AOB对硝化潜势的相对贡献及AOA和AOB硝化潜势的比值与环境因子皮尔森相关性分析Fig. 8 Pearson correlation analysis between the relative contribution to nitrification potential of AOA and AOB and the nitrification potential of AOA to AOB with the environmental parameters
4 讨论
由于溶解氧和底物不受限制,潜在硝化速率通常远高于原位硝化速率。Dang等[33]的研究表明,净硝化速率低于潜在硝化速率,但是仅低11.1%~61.1%。一些研究证明,潜在硝化速率与原位硝化速率具有很好的相关性,能用来表征原位硝化能力的高低[34-35]。渤海和南黄海海域表层沉积物中总潜在硝化速率为0.004 6~0.283 1 μmol/(g·d),低于其他河口或近海岸沉积物[16,18-19,23-24,32]。例如,美国马塞诸塞州Plum Island Sound河口沉积物(干重)潜在硝化速率为0~0.35 μmol/(g·d)[23];崇明岛东部潮间带沉积物潜在硝化 速 率 为0.55~0.98 μmol/(g·d)(夏 季)和0.05~0.17 μmol/(g·d)(冬季)[32];长江口潮间带沉积物潜在硝 化 速 率 为0.10~4.48 μmol/(g·d)(夏 季)和0.07~0.74 μmol/(g·d)(冬季)[16];辽河口海岸沉积物潜在硝化速率为0.000 1~0.336 8 μmol/(g·d)(春季)[24];乳山湾邻近海域沉积物中潜在硝化速率为0.782~1.666 μmol/(g·d)(夏季,显著高于冬季)[18-19]。而且,在渤海海域,潜在硝化速率一般在近岸站位较高,综上推测,河口和近海岸沉积物硝化潜势总体高于远海岸沉积物硝化潜势,这可能是因为近海更容易受到来自陆地的氮污染的影响。
由于细菌和古菌的细胞被膜以及中枢代谢和信息过程存在巨大差异,常见的抑制细菌的抗生素往往不会对古菌造成影响[36]。因此,抗生素是常用来区分AOA和AOB对硝化潜势的相对贡献的抑制剂类型之一。但是,也有研究发现抗生素可能会降低AOA的多样性[37]。此外,需要注意的是,抗生素的抑制作用表现出一定的时效性,例如,Wang等[38]发现18 d后氨苄青霉素对硝化作用的抑制效应开始减弱,这可能是由于其自身的降解或微生物的适应性。本研究的培养周期为3 d,氨苄青霉素能较好地发挥抑制效果,以区分AOA和AOB对硝化作用的相对贡献。研究表明,AOA和AOB对硝化潜势的相对贡献受到时间和空间的影响[16,19]。在长江口潮间带夏季和冬季沉积物中,添加和未添加氨苄青霉素的培养体系不存在显著差异,添加青霉素后硝化潜势仅下降了13.1%~26.6%,说明该区域AOA对硝化潜势的贡献率更大[16]。在乳山湾邻近海域沉积物中,AOA对硝化潜势的贡献在冬季高于AOB,在夏季则小于AOB[19]。本研究结果表明,在春季渤海和南黄海海域沉积物中,除南黄海H12和H18站位外,其他站位AOA对硝化潜势的贡献均高于AOB。以上研究表明,在河口及近海沉积物中,AOA的潜在硝化速率高于AOB,AOA在硝化作用中占有更加重要的地位,许多基于AOA和AOB丰度测定的分子生态学方法也证明了这一点[14-15,19,23]。然而,在一些淡水生态系统沉积物中,AOB对硝化过程的贡献常常高于AOA[10,33]。
一直以来,盐度对硝化速率的影响一直是研究者关注的焦点。盐度是影响河口沉积物中硝化速率的主要因素,通常随着盐度的增加,硝化速率会降低[16,23],乳山湾邻近海域也发现了盐度和潜在硝化速率显著的负相关性[19]。在本研究黄渤海海域,AOA和总潜在硝化速率均与盐度呈显著的负相关关系,与先前研究具有一致性;AOB潜在硝化速率与盐度呈正相关关系,但是相关性不显著(图7)。盐度对硝化作用的影响,一方面是因为沉积物中 NH+4的吸收受到盐度的影响,另一方面则是因为盐度对微生物代谢的影响[39-40]。在本研究的潜在硝化速率培养体系中, NH+4的添加及连续震荡培养,可以排除第一方面的原因。此外,电导率也是影响黄、渤海海域硝化潜势的一个重要环境因子。电导率与盐度具有一定的关系,可以通过盐度和温度进行换算,常用来表征盐度的高低[41]。安第斯山高纬度湿地沉积物研究表明,氨氧化潜能随着电导率的升高而降低[41]。但是,渤海海域和南黄海海域单独分析时,盐度却没有与潜在硝化速率表现出显著的负相关关系,推测是因为在单个海域内,盐度的变化范围较小。
渤海和南黄海海域总体分析及渤海海域单独分析均未发现溶解氧浓度与潜在硝化速率具有显著相关性,而南黄海海域单独分析发现溶解氧浓度和总潜在硝化速率及AOB潜在硝化速率具有显著负相关关系。好氧氨氧化为严格需氧过程,溶解氧浓度对AOA和AOB活动应该具有一定的促进作用[42]。推测是因为在潜在硝化速率培养实验中,氨氧化微生物不受溶解氧浓度的限制,潜在硝化速率与现场溶解氧浓度未表现出正相关关系。此外,潜在硝化速率没有表现出与温度的显著相关性。以前发现的AOB均属于中温细菌,最适生长温度一般在20~30℃[42],AOA对温度有极强的适应能力[43]。因此,在温度范围为8.50~12.30℃的春季黄、渤海海域,AOA是硝化潜势的主要贡献者。因其对温度变化适应性较好,因而未表现出随温度变化而变化的特征。
NH+4浓度对氨氧化速率(或硝化速率)具有重要作用。Christman等[17]在北冰洋沿岸水体中发现, NH+4浓度较高的冬天,潜在硝化速率也随之升高。Hou等[10]在太湖和巢湖沉积物中发现,潜在硝化速率随营养程度的增加而增加,但是Chang等[24]在辽河口的研究却表现出相反的趋势,营养物浓度高的站位,潜在硝化速率反而低,营养物质低的站位速率反而高。在本研究的渤海海域表层沉积物中, NH+4浓度与总潜在硝化速率、AOA和AOB潜在硝化速率均没有表现出显著的相关性。在潜在硝化速率培养实验中,氨氧化微生物不受底物 NH+4的限制,这可能是潜在硝化速率与原位 NH+4浓度没有显著相关性的原因之一。研究发现,和AOB相比,AOA对 NH+4的亲和力更高,更适合寡营养环境[12],这也是AOB在淡水生态系统中占主要地位,AOA则在海洋生态系统中占主要地位的原因之一。本研究20个站位 N H+4浓度为2.393~6.105 μmol/L,远低于一些淡水生态系统[10],因此,AOA在生长及硝化作用中占有主要优势。
然而,渤海8个站位AOA潜在硝化速率和总潜在硝化速率与 NO-3浓度具有显著的正相关关系。Dang等[33]也发现,AOB对硝化潜势贡献更大的水库沉积物中,AOB和总净硝化速率与 N O-3浓度具有显著的正相关关系。通常,作为硝化过程的产物, N O-3浓度并不会作为AOA和AOB参与硝化作用的一个关键环境因子。虽然,原位 NO-3浓度不会对AOA和AOB本身产生直接影响,但是其浓度的高低很可能反映了环境中氨氧化作用的强弱,这就解释了硝化潜势与NO-3浓度具有正相关关系的原因。
黄、渤海各站位pH显著影响潜在硝化速率分布,且与AOA和总潜在硝化速率呈现出显著的正相关关系(皮尔森相关性,r>0.645,p<0.01)。然而,研究表明,低pH沉积物中AOA对氨氧化作用的贡献可能比高pH更突出[42]。但是,当渤海海域和南黄海海域单独分析时,pH和潜在硝化速率并不呈现出显著正相关性;黄、渤海海域一起分析时,pH和潜在硝化速率的显著正相关性受到了渤海站位pH显著高于南黄海站位的影响。因此,不同海域影响硝化潜势的关键环境因子可能存在一定的差异,分别分析可能更有助于了解真正的关键环境因子。
5 结论
本研究表明,河口和近海岸沉积物硝化潜势总体高于远海岸;与AOB相比,AOA在氨氧化或硝化过程中起着更加重要的作用。此外,渤海海域硝化潜势显著高于南黄海海域;黄、渤海海域整体来看,盐度是影响潜在硝化速率的关键环境因子,渤海海域分析则表明越高的 N O-3浓度可能指示着越高的硝化潜势。
致谢:本研究采集样品及CTD数据取自“黄渤海渔业资源调查项目”渤海春季(2019年5月)航次和“国家自然科学基金委共享航次计划”2018年度项目渤黄海航次(航次编号:NORC2019-01)春季航次,分别由中国水产科学研究院黄海水产研究所“中渔科102”号和“北斗”号调查船实施,在此一并致谢。感谢中国海洋大学海洋生命学院刘晓收教授和钟鑫同学在样品采集中给予的帮助,感谢中国海洋大学化学化工学院高志梅同学在环境因子分析中给予的帮助。
