CRISPR/Cas9基因组编辑技术在病原真菌耐药性研究中的应用
2023-01-06张舒戴榕辰季聪华耿圆圆李若瑜龚杰
张舒 戴榕辰 季聪华 耿圆圆 李若瑜 龚杰
(1.中国疾病预防控制中心传染病预防控制所,传染病预防控制国家重点实验室,感染性疾病诊治协同创新中心,北京 102206;2.浙江中医药大学公共卫生学院,杭州 310053;3.北京大学第一医院皮肤性病科,国家皮肤与免疫疾病临床医学研究中心,北京大学真菌和真菌病研究中心,皮肤病分子诊断北京市重点实验室,北京 100034;4.北京大学第一医院-中国疾病预防控制中心传染病预防控制所“病原真菌联合实验室”,北京 102206)
病原真菌逐渐成为人类安全的重要威胁。据统计,在全球范围内,近10亿人患有皮肤、指甲和头发等浅表真菌感染性疾病,每年约150多万人因深部真菌感染性疾病死亡[1]。其中,曲霉、念珠菌、隐球菌以及双相真菌如荚膜组织胞浆菌等是导致大多数严重真菌病的主要病原真菌[1]。
对于病原真菌引起的感染,通常采用抗真菌药物进行治疗。传统的抗真菌药物主要包括多烯类、唑类、棘白菌素类,其数量有限,随着抗真菌药物的广泛使用,病原真菌在抗真菌药物的选择压力下逐渐产生了耐药性,甚至产生了多重耐药情况[2]。因此,研究病原真菌耐药性及机制是对抗病原真菌的一项重要任务。在这种情况下,亟需寻找高效的遗传操作工具以深入理解病原真菌的耐药机制。成簇的规律间隔的短回文重复序列及其相关蛋白(clustered regulatory interspaced short palindromic repeats/associated protein 9,CRISPR/Cas9)克服了过去基因工程在真菌研究中遇到的许多障碍,该系统通常参与细菌对噬菌体的适应性免疫,其中Cas9核酸酶不仅可以靶向病毒DNA,而且几乎能靶向任何物种的DNA,这一发现为该技术在真核生物中的应用奠定了基础[3]。本文总结了CRISPR/Cas9在基因功能研究中的应用,以及在人类主要致病真菌(念珠菌、曲霉等)耐药性研究中的应用和该技术的局限性,为更清晰地阐明真菌发病机制、更有针对性地防控和治疗真菌感染奠定基础。
1 CRISPR/Cas9技术在基因功能研究中的应用
真菌耐药机制的研究离不开分子遗传学的手段,然而,转化过程中的低同源重组率、有限的选择标记等阻碍了对基因功能的有效分析[4]。新型基因编辑技术——CRISPR/Cas9在基因工程领域得到广泛关注,其有两个关键组分:CRISPR相关核酸内切酶(CRISPR associated protein 9,Cas9)和单链向导RNA(single guide RNA,sgRNA),sgRNA引导Cas9在目标基因处发生DNA双链断裂(DNA strand breaks,DSB)[3,5]。断裂的DNA双链可以通过同源定向修复(homology-directed repair,HDR)或非同源末端连接(non-homologous end joining,NHEJ)的方式进行修复[6]。
目前,CRISPR/Cas9已成功应用于多种病原真菌的遗传操作,如烟曲霉(Aspergillusfumigatus)[7-8]、黑曲霉(Aspergillusniger)[9]、新生隐球菌(Cryptococcusneoformans)[10-11]、白念珠菌(Candidaalbicans)及其他念珠菌[12-13]。研究表明,在这些病原真菌中,利用CRISPR/Cas9系统的精准切割、HDR的高效修复,可以敲除基因、整合外源基因、替换基因(如替换启动子)、引入单碱基突变等(见图1),这对病原真菌的耐药性研究、新药物的发现以及真菌病的治疗具有重要意义。
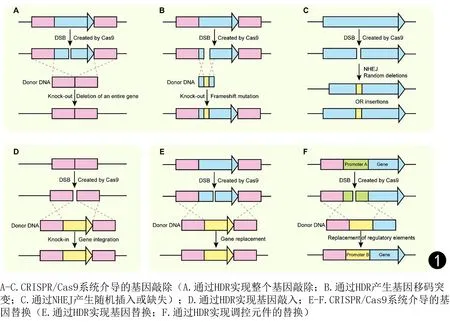
图1 CRISPR/Cas9系统介导的基因敲除、基因敲入和基因替换[14]Fig.1 CRISPR/Cas9 system-mediated gene knock-out, gene knock-in and gene replacement[14]
CRISPR/Cas9介导的基因敲除可通过HDR或NHEJ来完成。当CRISPR/Cas9在目标基因处产生DSB时,加入含有同源序列的供体DNA进行修复,若供体DNA仅由目标基因两侧的同源序列构成,则利用HDR可敲除整个基因(见图1A);若供体DNA由包含目标基因的同源序列构成,则会在HDR时引起移码突变从而破坏目标基因(见图1B)。当不提供供体DNA时,NHEJ修复系统会在DSB处随机引入小片段的“突变”(碱基的插入或缺失),导致目标基因发生移码突变或终止密码子提前出现,最终使靶基因丧失功能(见图1C)。例如,在葡萄牙念珠菌(Candidalusitaniae)中利用瞬时表达的CRISPR/Cas9系统进行基因敲除时,通过HDR成功进行ADE2的敲除[15]。Vyas等[12]在白念珠菌中设计了将CRISPR/Cas9系统整合到基因组中的质粒系统,在ADE2中提前引入终止密码子,从而阻断了该基因的功能。此外,在烟曲霉中,Fuller等[7]以产生黑色素所必需的pksP为研究对象,通过NHEJ途径破坏了pksP。
CRISPR/Cas9介导的基因敲入通常以HDR方式来完成。当CRISPR/Cas9系统在基因组的靶位点产生DSB后,加入含有同源序列的供体DNA,该供体DNA的两侧是DSB处的同源序列、中间是待敲入基因,即可通过HDR将外源基因整合进基因组,实现基因敲入(见图1D)。Zhang等[16]通过HDR介导的基因敲入,在预测位点高效、精确地整合了外源GFP标签。此外,利用CRISPR/Cas9系统还可实现基因的替换,包括调控元件(如启动子)的替换(见图1E、F)。当CRISPR/Cas9系统在基因组的靶位点产生DSB后,加入含有同源序列的供体DNA,该供体DNA的两侧是目的基因两侧的同源序列、中间是待替换基因,通过HDR完成基因的替换。
CRISPR/Cas9技术不仅可以通过DSB进行基因编辑,还能进行单核苷酸突变,即碱基编辑。碱基编辑是通过将失去催化活性的Cas蛋白突变体(如deactivated Cas,简称dCas)或具有切割单链活性的Cas蛋白(如nickase Cas,简称nCas)和作用于单链DNA(single-stranded DNA,ssDNA)的脱氨酶进行融合,实现对靶点的碱基替换。这种基因组编辑方法的优点是它不会产生双链DNA断裂,也不需要供体DNA模板,而是直接编辑单个核苷酸。Huang等[9]首次报道了丝状真菌中基于CRISPR/Cas9系统的单碱基编辑,在黑曲霉中融合nCas9和胞嘧啶脱氨酶,在靶位点实现了使C向T的高效转变。
2 CRISPR/Cas9技术在病原真菌耐药性中的应用
病原真菌通常通过干扰药物对于药物靶位点的作用或降低细胞内药物水平来实现耐药[17]。然而,病原真菌耐药的分子机制在不同物种甚至不同株系内并不完全相同,CRISPR/Cas9基因组编辑技术能对病原真菌的基因组进行精准编辑,在耐药性研究中发挥了至关重要的作用。
2.1 CRISPR/Cas9技术与传统耐药
真菌唑类和多烯类耐药性的产生通常与麦角甾醇合成相关基因的突变或过表达有关。其中Erg11/Cyp51(羊毛甾醇14α‐去甲基化酶)与唑类耐药的相关性被广泛研究和表征。已有研究利用CRISPR/Cas9技术,对ERG11/CYP51进行单碱基编辑,阐明了Erg11/Cyp51与唑类耐药的关系[18],还证明了在Cyp51A中直接参与唑类耐药的位点Gly138[8,19]。此外,陆续有研究者利用CRISPR/Cas9介导的基因敲除手段,证明了曲霉中CYP51A、CYP51B的地位和作用[20-21],从而建立了碱基突变及基因缺失与耐药表型之间的联系(见表1)。
药物转运蛋白编码基因的过表达也是真菌耐药的主要原因之一。利用CRISPR/Cas9技术,证明了多种病原真菌的转运蛋白Cdr1、Mfs7以及转运蛋白的调控因子Tac1B、Mrr1等,在三唑类药物及5-氟胞嘧啶耐药中的作用[22-24]。另外,还用CRISPR/Cas9对葡萄牙念珠菌的多烯类敏感性进行了研究,证明了参与甾醇生物合成的Erg3/Erg4与多烯耐药的关系[24](见表1)。
CRISPR/Cas9基因编辑还揭示了病原真菌对棘白菌素的耐药机制。棘白菌素的抗性通过葡聚糖合酶Fks亚基高保守区内氨基酸的突变来实现,使用CRISPR/Cas9技术在棘白菌素敏感的光滑念珠菌及白念珠菌中构建Fks突变体,能获得棘白菌素敏感性降低的突变菌株,从而筛选出与棘白菌素耐药/敏感性相关的基因及位点[25-26](见表1)。
2.2 CRISPR/Cas9技术与新药耐药
CRISPR/Cas9可用于验证新型抗真菌药物的耐药机制。在新型抗真菌药物的选择压力下,可获得新药耐药的突变株。利用CRISPR/Cas9技术对突变位点进行靶向改造,可验证新型抗真菌药物的耐药机制。如在敏感的光滑念珠菌中对新药靶酶Gwt1(糖基磷脂酰肌醇合成酶)按预测位点进行反向突变,获得抗新型抗真菌药物的耐药株,从而揭示新型抗真菌药物的耐药机制[27](见表1)。
2.3 CRISPR/Cas9技术与新药靶点
利用CRISPR/Cas9靶向病原真菌的必需基因,可作为挖掘潜在的新药靶点的策略。通过使用CRISPR/Cas9对病原真菌进行基因操作时,可将与生长和毒力相关的必需基因作为药物靶点。如念珠菌中与酿酒酵母同源的关键基因CDC8和CDC43[28-29]、编码GTP环水解酶(产核黄素的关键酶和限速酶)的基因RIB1[30]等均已证明可作为新型抗真菌药物的靶点(见表1)。

表1 CRISPR/Cas9系统在病原真菌耐药性研究中的应用Tab.1 Application of CRISPR/Cas9 system in drug resistance of pathogenic fungi
3 CRISPR/Cas9系统的局限性
利用CRISPR/Cas9介导的基因编辑技术能有效地对真菌靶基因进行编辑,但该技术在应用中也有局限性,如存在PAM(protospacer adjacent motif,前原间隔序列邻近基序)的限制、脱靶效应以及突变体筛选工作量大等问题。
CRISPR/Cas9系统对靶位点的识别受限于特定的PAM识别序列。以经典的酿脓链球菌Cas9蛋白为例,其识别的PAM序列为NGG,因此目标基因中必须含有NGG序列才能进行基因编辑,从而极大地限制了其应用。长期以来,研究者们致力于寻找拓展PAM识别序列的方法,如对Cas9蛋白进行改造,构建出可识别NGA、NGCG[31]、NGG、NG、GAA、GAT[32]的Cas9蛋白变体;此外,在其他菌株中寻找新型Cas9蛋白也是拓展PAM序列的方法,如Deveau等[33]发现嗜热链球菌的Cas9识别的PAM序列为NNAGAAW。这些研究均大大丰富了Cas9可识别的PAM范围,从而能靶向更多的序列。
在利用CRISPR/Cas9系统对目标基因进行编辑时,设计的sgRNA会与非靶点DNA序列发生错配,从而引入非预期的基因突变,产生脱靶效应(off-target effects),严重限制了CRISPR/Cas9基因编辑技术的广泛应用。有研究通过改造Cas9来提高与靶序列结合的保真性,如换用单切口酶活性突变体dCas9,每条sgRNA引导的Cas9只能切开一条DNA链,只有两条sgRNA同时在靶位点完成切割,产生DSB,才能实现基因组DNA的断裂。如果一条sgRNA错配,只会形成一个切口,不会形成DSB,很快被修复;而两条sgRNA同时错配,形成双链断裂的脱靶机率很小,这样就极大地提高了靶点专一性,减少脱靶效应。此外,设计特异性的sgRNA也是解决脱靶效应的策略之一,如有报道指出在sgRNA的5' 端加上GG[34]、将sgRNA的引导序列截短到17~18个核苷酸[35],在保证结合率的基础上,可以大大降低脱靶效应。
此外,CRISPR/Cas9系统在进行少量的基因编辑时具有高效性,而当需要构建大规模突变体时,单独转化和筛选突变体也是一项艰巨的任务。
4 总结与展望
CRISPR/Cas9基因组编辑技术,可进行基因敲除、基因敲入、基因替换、碱基编辑等,已在许多医学真菌中成功应用,并在耐药性研究中做出巨大贡献。CRISPR/Cas9的有效性和灵活性使其在医学真菌学领域产生巨大影响,除研究耐药机制外,CRISPR/Cas9系统还应用于阐明病原真菌的致病机制,解析致病因子的作用,研究潜在的新药物靶点等。本文中我们关注了临床上涉及的主要病原真菌,而CRISPR/Cas9方法在本文未提及的其他人类病原真菌,如马拉色菌[36]、镰刀菌[37]和双相真菌[38]中也很重要,同样值得关注。然而,为了建立一个有效的CRISPR/Cas9真菌系统,还需要解决一些局限性和挑战,如脱靶效应等。当前CRISPR/Cas9系统在模式真菌中正在快速发展,未来可将其推广到这些病原真菌中来,如利用CRISPR/Cas9介导的大染色体缺失和融合来研究大染色体重排与耐药的相关性,以进一步解决对抗真菌药物的耐药问题。
