四川省红火蚁分布调查及地理种群遗传结构研究
2023-01-06张添翼李旭阳贡常委汪雨蒙朱新成王学贵
张添翼,李旭阳,贡常委,汪雨蒙,文 耀,朱新成,李 彬,王学贵*
(1.四川农业大学农学院,成都 611130;2.成都绿金高新技术股份有限公司,成都 610023)
红火蚁(Solenopsis invicta Buren)是一种原产于南美洲的外来入侵物种[1],主要取食农林作物种子、果实及根系,筑巢行为引起电线短路或设施故障,叮蛰人畜造成灼伤疼痛甚至休克和死亡。红火蚁在世界范围内扩散速度十分迅速。上世纪开始在美国广泛扩散[2];2003年入侵台湾后迅速蔓延[3];2004年入侵广州[4],随后经过调查发现广西、福建、湖南等省亦有红火蚁的存在[5]。根据农业农村部公布的全国农业植物检疫性有害生物分布行政区名录,四川省于2012年首次在攀枝花市发现红火蚁,2013年在西昌发现红火蚁,2019年红火蚁已广泛分布于攀西地区[6]。据调查,红火蚁在四川省具有高度危险性[7-8],其适生范围广、抗逆性强、繁殖速度极快[9],且四川地区气候温暖、资源丰富,为红火蚁的入侵提供了绝佳的环境条件,每年的雨季是红火蚁婚飞的高峰期[10],为四川省的红火蚁防控带来极大压力。以往红火蚁主要分布于攀西地区,但近年来红火蚁在四川省迅速扩散,红火蚁疫情发生程度有加重的趋势,给四川省的生态环境和人民安全带来严重威胁。
红火蚁的传播方式多样,以人为传播为主,随苗木调运等人为因素入侵新的区域[11]。不同入侵来源的红火蚁及其在新的入侵区域的适生性可能导致红火蚁种群出现一定的遗传分化[12]。就红火蚁检疫而言,明确入侵红火蚁种群来源尤为重要。微卫星(simple sequence repeat,SSR)分子标记是目前预测红火蚁种群来源的一种快速、简便和准确的分析技术[13]。邵敬国[14]利用微卫星位点对红火蚁种群遗传多样性进行分析,推测我国红火蚁入侵源可能为美国。黄奕雯等[12]对我国各省红火蚁种群进行微卫星分析,认为国内红火蚁入侵种群与美国亚拉巴马州红火蚁种群在遗传距离上十分接近。利用微卫星位点的多态性来分析物种进化、遗传多样性已有大量报道[15-17]。对四川省红火蚁种群分布调查和遗传分化分析可以明确红火蚁在四川省的入侵来源、入侵路线及进化过程,有助于四川省今后制订更有针对性的红火蚁检验检疫措施。本试验使用SSR分子标记对四川省红火蚁地理种群的遗传结构进行探索,希望揭示四川省红火蚁的遗传进化过程及地理种群间的亲缘关系,为控制红火蚁在四川省的扩散提供帮助。
1 材料和方法
1.1 调查地点及调查方法
根据四川省农业农村厅和四川省林业和草原局印发的四川省红火蚁发生县、乡级行政区名录,于2021年7—8月前往攀枝花市、泸州市、绵阳市、广元市、达州市和凉山州西昌市等红火蚁发生区域,在当地植保站人员的协助下进行红火蚁疫情等级调查,根据红火蚁疫情监测规程[18]确定其发生程度。一级:轻度,活蚁巢为≤0.1个/100 m2;二级:中度,活蚁巢为(0.11~0.5)个/100 m2;三级:中偏重,活蚁巢为(0.51~1.0)个/100 m2;四级:重度,活蚁巢为(1.1~10)个/100 m2;五级:严重,活蚁巢>10个/100 m2。
1.2 采样
使用100 mL离心管作为诱集瓶,瓶内装有涂抹蜂蜜的火腿肠放置于距蚁巢30 cm处诱集红火蚁。诱集30 min后取回诱集瓶,加入10%丙酮水溶液,待红火蚁全部死亡后使用100目筛网过滤收集红火蚁,于-80℃超低温冰箱中保存备用。
1.3 DNA提取
参考改良的CTAB法[19],提取红火蚁样本的DNA,用核酸浓度测定仪检测DNA纯度,-20℃保存备用。
1.4 SSR-PCR扩增及凝胶电泳检测
根据前人报道的SSR序列合成8对引物,引物序列为:
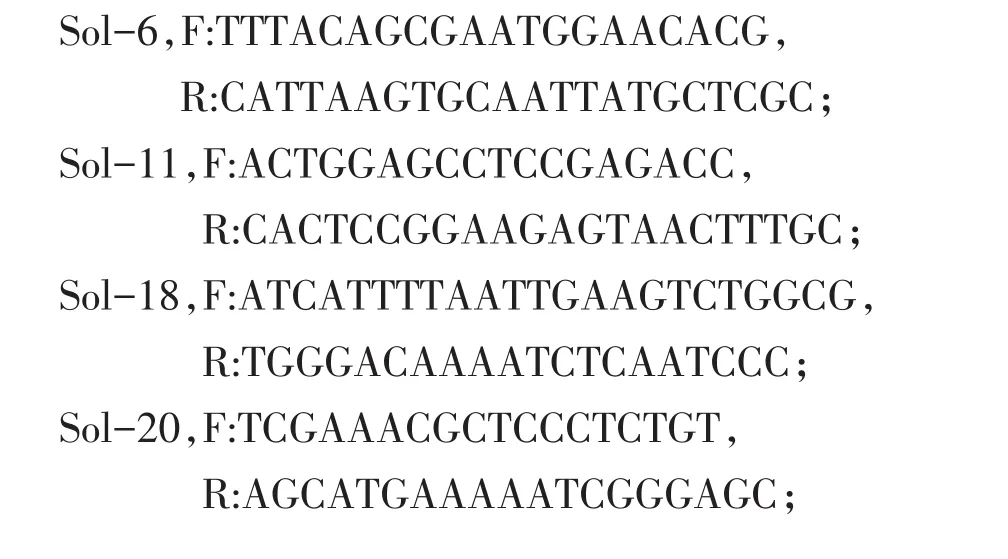

参照其反应体系进行PCR扩增[20]。3%琼脂糖凝胶电泳,控制电压为80 V恒压,电泳100 min。置于凝胶成像仪中成像观察并拍照,将所得图像直接在成像系统中进行处理,对扩增结果读带。所有引物均由成都擎科生物有限公司合成。
1.5 数据处理与分析
按照电泳图谱中同一位置上DNA带进行统计,根据分子量大小对扩增结果读带以二倍体形式记录 ,从大 到 小依 次 记作 A,B,C,……[21]。 用POPGENE 1.32软件在假定种群处于Hardy-Weinberg平衡状态下,对全部种群和各单个种群分别进行遗传参数分析,分别计算多态位点百分率(P)、观测等位基因数(Na)、有效等位基因数(Ne)、基因多样性指数(Nei's)、Shannon信息指数(I)、表观杂合度(Ho)和预期杂合度(He)、种群内近交系数(Fis)、总近交系数(Fit)、种群间分化系数(Fst)、种间信息流(Nem)、Nei's标准遗传距离(D)和遗传相似度(I)。根据D,利用NTSYS-pc 2.1软件对各种群进行非加权组平均法(UPGMA)聚类分析。
2 结果与分析
2.1 疫情分析
2.1.1 各地红火蚁发生调查
结果表明(表1),绵阳的红火蚁发生程度达到中偏重程度,疫情等级为3级;西昌的红火蚁发生程度从轻度到重度不等,疫情等级为1~4级;攀枝花的红火蚁发生程度从轻度到重度不等,疫情等级为1~4级;会理的红火蚁发生程度达到重度,疫情等级为4级;达州的红火蚁已被彻底防控,疫情等级为0级;广元的红火蚁发生程度为轻度,疫情等级为1级。
2.1.2 不同生境类型的红火蚁发生情况
结果表明(表1),各地的红火蚁发生的生境包括:草坪、建筑荒地、农田、公园、绿化带、果园、山路、公路等。红火蚁发生较多的生境为:草坪、公园、果园、农田、公路5种生境,发生程度达到重度程度,疫情等级为4级。
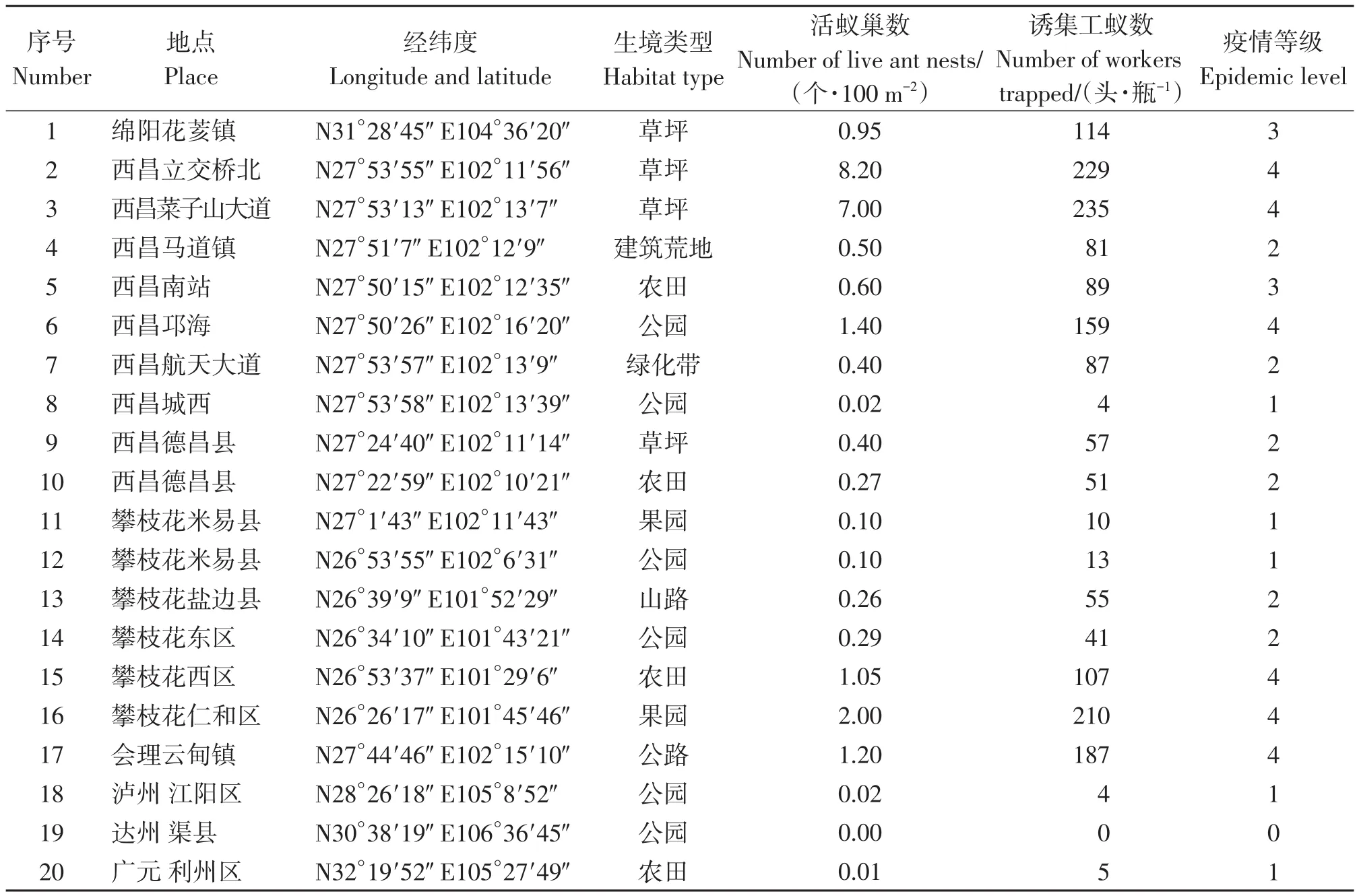
表1 2021年7—8月四川省红火蚁疫情发生情况Table 1 Epidemic situation of Solenopsis invicta in Sichuan Province from July to August 2021
2.2 SSR-PCR扩增结果
选取8个SSR位点对5个红火蚁地理种群的基因组DNA进行PCR扩增和电泳分析,共获得出60个多态条带,部分扩增结果见图1。
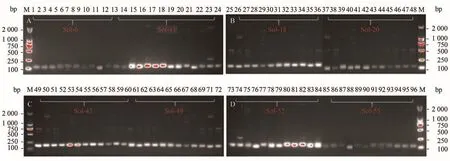
图1 8对SSR引物对部分红火蚁种群基因组DNA的PCR扩增Figure 1 PCR amplification of 8 pairs of SSR primers on genomic DNA of some Solenopsis invicta populations
2.3 5个红火蚁地理种群的遗传多样性
结果表明(表2),8对微卫星引物在5个红火蚁地理种群中共检测到60个等位基因,每个微卫星位点上平均有7.5个等位基因,在位点Sol-55上的等位基因最多为9个,绵阳种群在位点Sol-18上未检测到等位基因。5个红火蚁地理种群的多态位点百分率为36.67%~78.33%,其中,绵阳种群的多态位点百分率最低,攀枝花种群的多态位点百分率最高;各种群在8个微卫星位点上观测等位基因数的平均值为2.750 0~5.875 0,其中绵阳种群在最低,攀枝花种群最高;各种群在8个微卫星位点上有效等位基因数的平均值为1.899 2~4.065 5,其中绵阳种群最低,攀枝花种群最高;各种群的Shannon信息指数为0.502 7~0.619 1;各种群的表观杂合度为0.570 9~0.622 9;各种群的预期杂合度为0.555 6~0.616 1;西昌种群的基因多样性指数最高为0.429 2,攀枝花种群的基因多样性指数最低为0.315 1。各位点的等位基因表观杂合度与预期杂合度基本一致,均相差不大,均符合Hardy-Weinberg平衡。
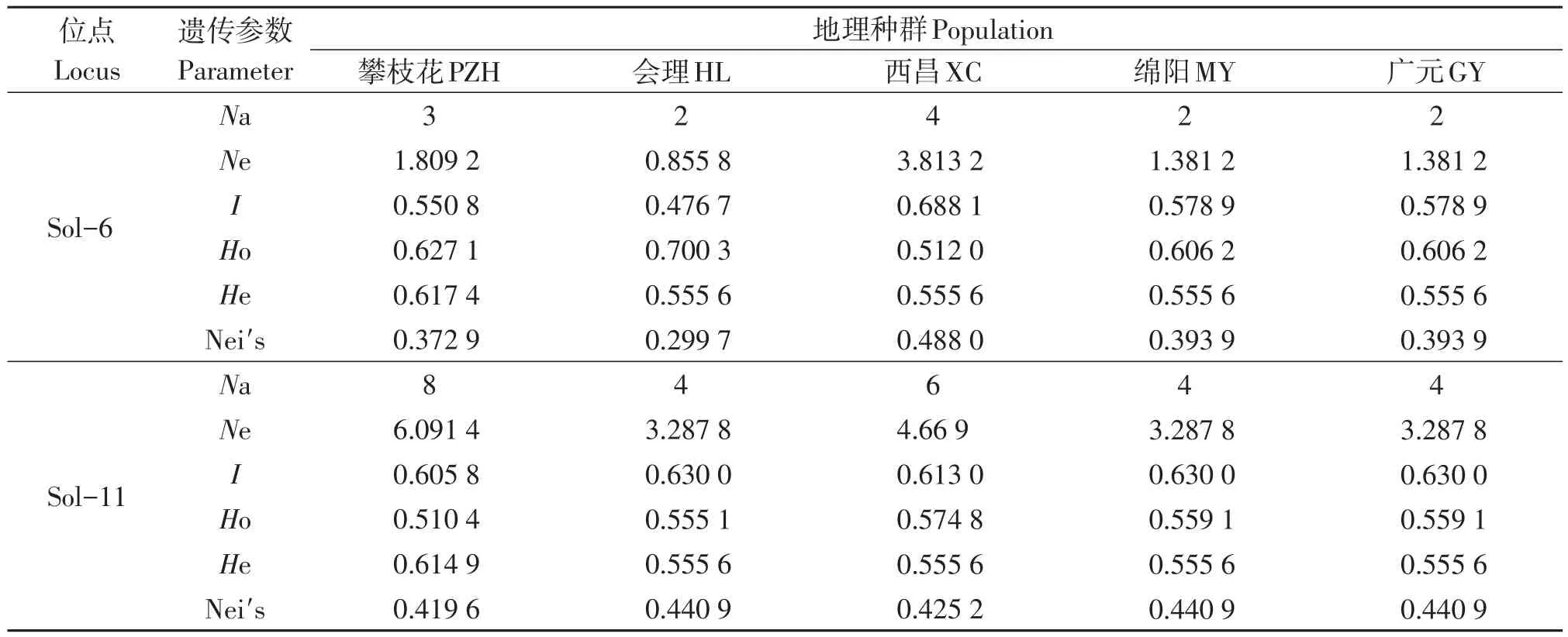
表2 5个红火蚁地理种群在8个微卫星位点的多样性指数Table 2 Diversity index of 5 geographical populations of Solenopsis invicta at 8 microsatellite sites
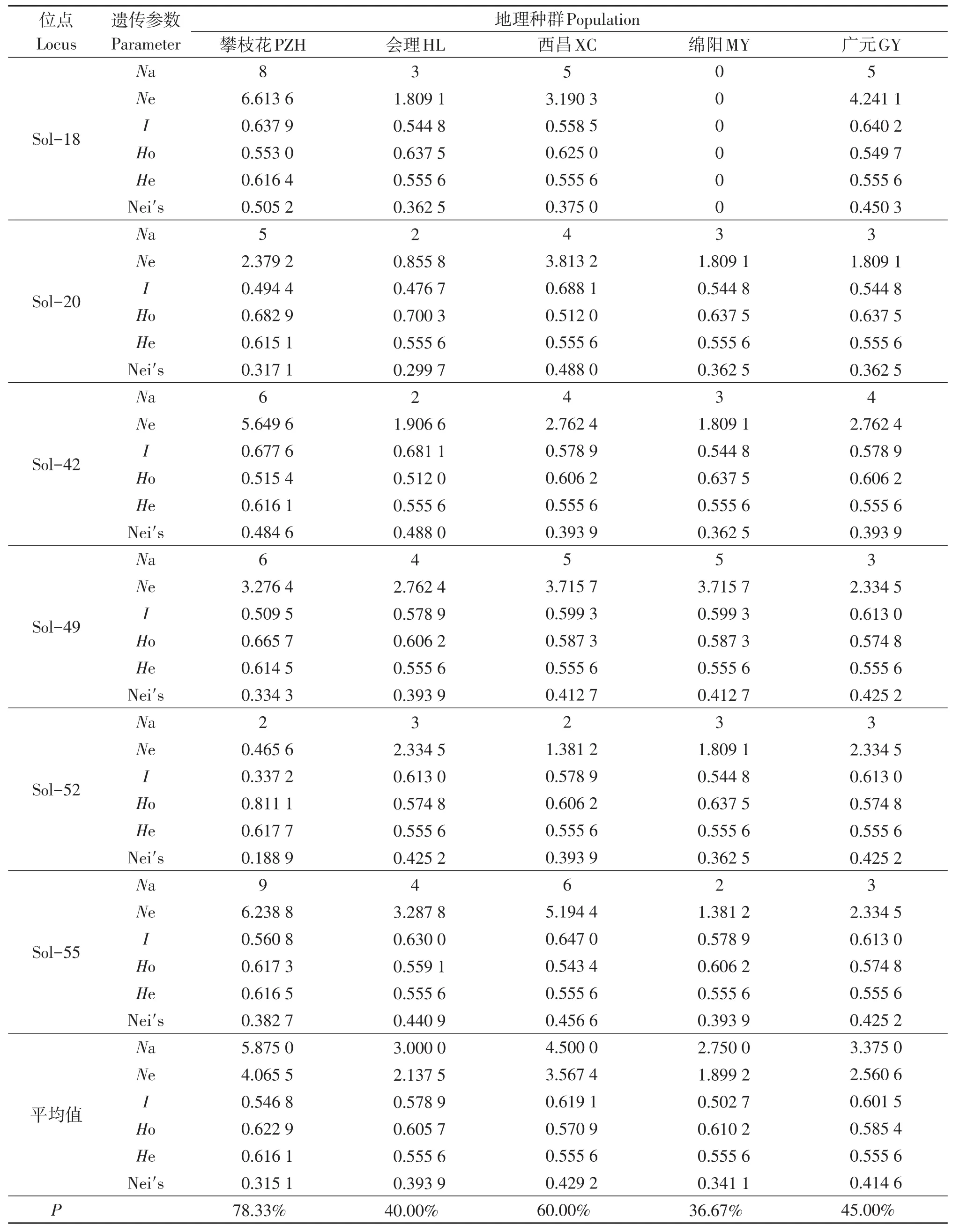
(续 表2)
2.4 5个红火蚁地理种群的遗传分化
结果表明(表3),四川省5个红火蚁地理种群在8个微卫星位点的种群内近交系数为0.298 8~0.461 2,平均值为0.405 7;总近交系数为0.182 7~0.294 3,平均值为0.245 0。种群间分化系数为0.262 4~0.576 3,平均值为0.400 0,表明约60%的遗传变异发生在种群内,约40%的遗传变异发生在种群间。各种群的基因流为0.517 4~2.456 2,平均值为1.508 8,其中位点Sol-18的最低为0.517 4,位点Sol-20最高为2.456 2。四川省5个红火蚁地理种群间的标准遗传距离和遗传相似度结果表明(表4),5个地理种群两两之间的遗传相似度范围为0.646 4~0.848 6,均大于0.6,这表示各种群间的基因交流较为频繁。攀枝花种群和西昌种群之间的遗传距离最小为0.164 2,绵阳种群和广元种群之间的遗传距离最大为0.432 8。
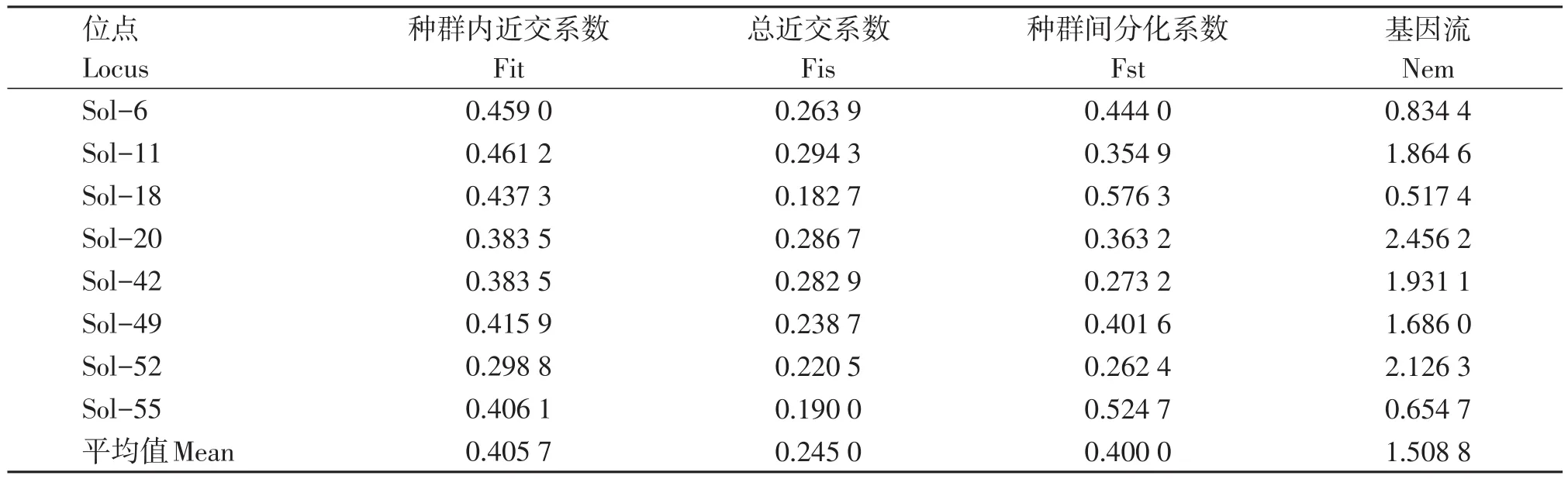
表3 各位点固定指数及基因流Table 3 The fixed index and gene flow of 8 microsatellite loci
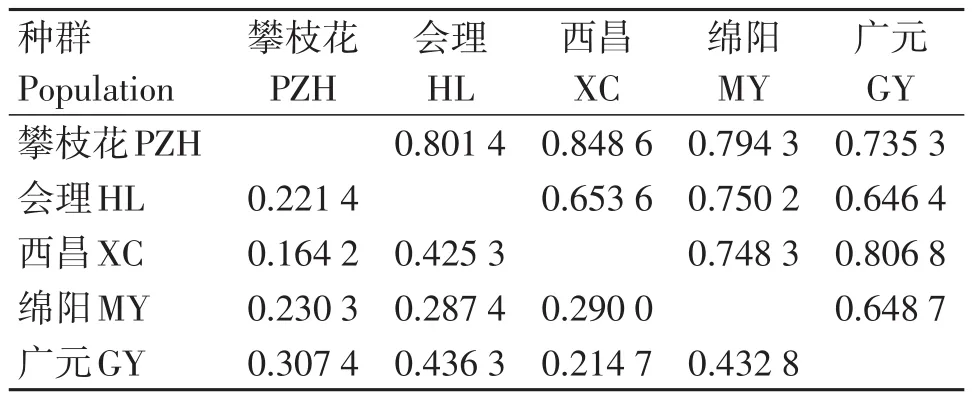
表4 地理种群间Nei's标准遗传距离(左下角)和遗传相似度(右上角)Table 4 Nei's standard genetic distance(lower left)and genetic identity(upper right)among geographical populations
2.5 聚类分析
使用NTSYS-pc 2.1软件对5个种群的遗传距离进行UPGMA聚类分析,结果表明(图2),系统发育树首先分为两大分支,其中攀枝花种群与其它种群间的遗传距离最远,单独为一个亚群;会理、广元、绵阳、西昌4个种群为一个亚群。
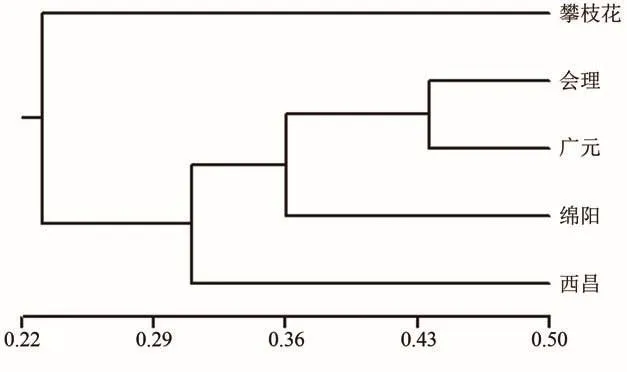
图2 地理种群的UPGMA聚类分析Figure 2 UPGMA cluster analysis of geographical populations
3 讨论
3.1 四川省红火蚁种群的遗传多样性分析
遗传多样性的丰富与否直接影响了红火蚁能否适应复杂的环境变化[22],多态位点百分率、Shan‐non信息指数及基因多样性指数均是衡量种群遗传多样性的指标[23]。种群杂合度反映了地理种群的遗传差异,种群杂合度的高低与群体的遗传一致性成反比[21]。本次调查采集的样品中,攀枝花种群和西昌种群的样本量较多,其在8个SSR位点上所检测出的有效等位基因数也最多,这与闫路娜和张德兴[24]的研究结果一致。根据D.Botstein等[25]提出的多态位点百分率(P)作为衡量遗传多样性的一个重要指标:当P>50%时为高度多态位点;25% 影响物种种群遗传分化有多方面的因素,诸如有性生殖、体细胞突变、选择、基因流、遗传漂变及环境的影响[21]。本次调查的红火蚁采集于不同生境,5个红火蚁种群的种群间分化系数平均值达到0.4,表明种群间出现一定程度的遗传变异。基因流是影响种群遗传分化的一个重要因素[13],基因流水平越高,种群间的遗传分化程度越低,种群间遗传趋于一致[15]。根据S.Wright[26]提出的:当种群间的基因流大于1时,可以抵制种群内因遗传漂变而引起的种群遗传分化。本次研究中的5个红火蚁种群在8个SSR位点上的基因流平均值为1.508 8,种群间基因交流频繁,种群间较高的遗传相似度(均大于0.6)也验证了这一点。在黄奕雯等[12]的研究中,中国内陆各省、香港及美国的红火蚁种群间遗传分化程度较高,可能受地理隔离影响。四川省河流极多,雨季漫长,为红火蚁顺水扩散和婚飞提供了有利条件[10],地理隔离不能完全阻止红火蚁在四川省的传播扩散,各分布区间存在一定的基因交流。 利用种群间的遗传距离可以估测其进化关系[14],目前应用较多的遗传距离指标是M.Nei提出的Nei's标准遗传距离[27]。国内对于我国红火蚁入侵扩散途径已有一些报道:邵敬国等[14]推测我国红火蚁入侵源可能是美国不同地区;黄奕雯等[12]认为地理距离较近的红火蚁种群通过自然扩散形成,地理距离较远的红火蚁种群由人为传播导致。根据Nei's标准遗传距离构建的UPGMA聚类图可以看出,会理、广元、绵阳和西昌种群聚为一支,这些种群间地理距离较远,推测可能是由人为传播导致;攀枝花种群单独聚为一支,推测攀枝花红火蚁种群与四川省其它红火蚁地理种群的入侵源不同。红火蚁在自然条件下的远距离传播能力很弱[28],在短时间内不可能依靠自然传播大面积扩散。由此看出,四川省红火蚁各种群间不仅存在着受环境因素影响如:顺水扩散、婚飞等导致的自然扩散;还存在着因各种人为因素如:苗木调运、长途贸易运输等导致的长距离种群扩散。 红火蚁在攀西地区报道发生已有近10年历史,在四川省其它地区仅近几年零星发生且很快扑灭,但这已为四川省的红火蚁防控敲响警钟。本试验利用SSR分子标记对四川省红火蚁地理种群进行遗传多样性分析发现:5个红火蚁地理种群间具有丰富的遗传多样性,遗传分化程度不高,自然扩散和人为传播并存。建议应加强对目前已有红火蚁疫区尤其是攀西地区的监控与防治,严防红火蚁在雨季的大量繁殖扩散,加强检疫工作防止红火蚁因人为传播导致的远距离扩散。3.2 四川省红火蚁种群的遗传分化
3.3 四川省5个红火蚁种群间的扩散途径
4 结论
