贵州省野生皂荚种实表型性状多样性分析
2023-01-04田红红陆春云
田红红,杨 菊,陆春云,肖 枫,赵 杨*
(1 贵州大学 林学院,贵阳550025; 2 贵州省森林资源与环境研究中心,贵阳 550025)
皂荚(GleditsiasinensisLam.)是豆科(Leguminosae)皂荚属(Gleditsia)的高大落叶乔木,分布广泛,喜温暖湿润气候及深厚肥沃稍湿润的土壤,具有较高的生态价值[1-3]。同时皂荚种子富含“皂角米”,是一种经济价值极高、口感细腻、润滑的营养保健食品[4-5];皂角刺还可用作中药[6],因此皂荚具有很大的开发和应用前景。
植物表型多样性是环境多样性与遗传多样性的综合体现,它主要是研究种群在其生境内受各种环境影响而产生的变异,是生物多样性的重要研究内容[7],表型性状的研究具有快捷简单等优点,至今在群体遗传多样性的研究中仍然是一个常用的方法[8]。国内近年来对豆科植物扁蓿豆(Medicagoruthenica)[9]、鹰嘴豆(CicerarietinumL.)[10],以及云南茶树(CamelliasinensisL.)[11]、新疆野杏(ArmeniacavulgarisL.)[12]、垂珠花(StyraxdasyanthusPerkins)[13]等植物进行表型多样性研究,并在一定程度上解释了其遗传变异规律。
目前,国内学者对于皂荚的研究主要集中在种实运动[14]、引种栽培[15]、皂荚刺的发育[16]及其药用成分[17]、以及抗逆性及种源试验[18]等方面,李伟等[19]在2013年研究了中国南方皂荚天然群体种实的表型多样性,仅包含了贵州省两个区域的皂荚群体,而对于贵州省内皂荚资源的分布及群体内的变异情况未做全面分析。因此本研究在全面调查贵州省皂荚资源分布区域的基础上,对皂荚群体内和群体间的种实表型多样性、变异规律及相关性进行了分析,以期为皂荚资源的挖掘、创新和保护提供科学支撑,对贵州省优良皂荚品种选育及遗传改良提供理论基础。
1 材料和方法
1.1 材料来源以及采种地的自然条件
于2020年10~12月在贵州省皂荚自然分布区内,按照贵州省行政区域将皂荚群体进行划分,采集每个单株不同方位的荚果,记录采集编号,置于室温保存。采样单株生长健壮,且无明显病虫害或其他缺陷。观测并记录采种地地理信息,各皂荚野生群体的立地条件见表1。

表1 贵州省7个皂荚群体基本信息
1.2 表型性状测定
从采集的单株中选取成熟的荚果,荚果在实验室自然阴干处理,每个单株随机测定30个荚果及种子,用直尺测定荚果长(pod length,PL)、荚果宽(pod width,PW)(最宽处直径测量),精确到0.01 cm,游标卡尺测定荚果厚(pod thickness,PT)(最厚处测量)、种子长(seed length,SL)、种子宽(seed width,SW)、种子厚(seed thickness,ST),精确到0.01 mm。计算种子长宽比(product seed aspect ratio,SL/SW)(形态指数)、种子长宽乘积(seed length plus width,SL×SW)(种子大小)。用0.001电子天平测量荚果质量(pod mass,MP),并记录每荚粒数(number of seeds produced,PNS)。
1.3 数据分析
利用Excel 2010 软件对皂荚各种实表型性状数值进行整理,并计算表型变异系数和多性状综合评价值。表型变异系数公式:CV(%) =σ/μ×100,式中σ为表型标准差,μ为某一行状群体平均值。
参照葛颂等[20]的方法,采用SPSS tatistics 25软件对各种实表型性状进行巢氏方差分析,方差线性模型为:Yijk=μ+Si+T(i)j+e(ij)k,式中,Yijk为第i 个群体第j个单株第k 个观测值;μ为总均值;Si为种群效应(固定);T(i)j为群体内单株效应(随机);e(ij)k为试验误差;
群体间表型分化系数的定义和分析参照王娅丽[21]的方法。表型分化系数公式为:VST=(σ2t/s)/(σ2t/s+σ2s),式中σ2t/s为群体间方差分量,σ2s为群体内方差分量,VST表示群体间变异占遗传总变异的百分比。
利用ggbiplot package进行主成分分析(Principal components analysis,PCA)及各表型性状间的相关性分析。
2 结果和分析
2.1 贵州省野生皂荚各群体表型性状的平均值
贵州省7个皂荚群体的荚果和种子表型性状的差异见表2,不同群体间各表型性状均存在不同程度的差异。
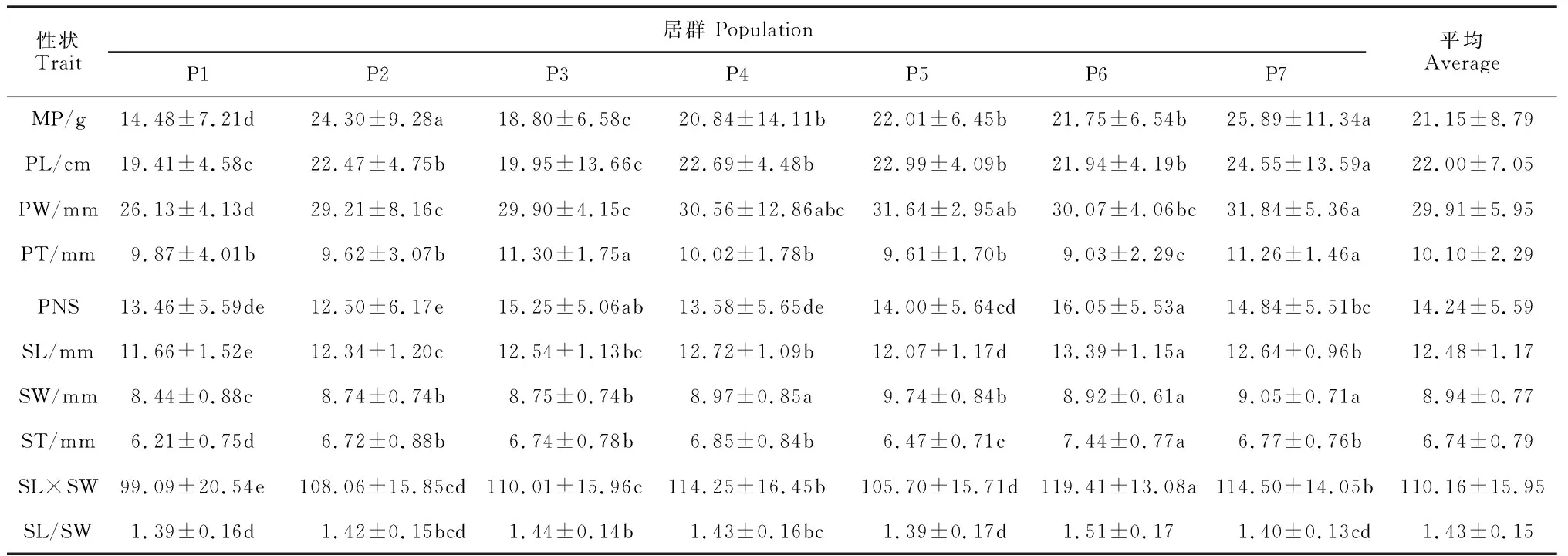
表2 野生皂荚群体各表型性状的平均值
在荚果表型性状中,惠水县(P7)群体的荚果质量(25.89 g)、荚果长(24.55 cm)、荚果宽(31.84 mm)均最大,总体显著大于安顺市(P1)和贵阳市(P3)两个群体;独山县(P6)群体的荚果厚(9.03 mm)最小,且显著小于贵阳市(P3)和惠水县(P7)两个群体。在5个种子表型性状中,思南县(P5)的种子宽(9.74 mm)最大,与福泉市(P2)和贵阳市(P3)2个群体无显著差异,与安顺市(P1)、凯里市(P4)、惠水县(P7)3个群体差异显著,独山县(P6)群体其余种子性状平均值均为最大,且与其他几个群体差异显著。
2.2 皂荚表型性状的方差分析
皂荚各表型性状的方差分析结果表明,除每荚粒数、种子宽、种子长宽积及种子长宽比在群体间未达到显著水平外,其余表型性状的分析指标在群体间和群体内均达极显著水平(表3)。说明皂荚的荚果表型性状在群体间和群体内都具有较高的多样性,而种子的表型性状在群体内表现出更高的多样性。
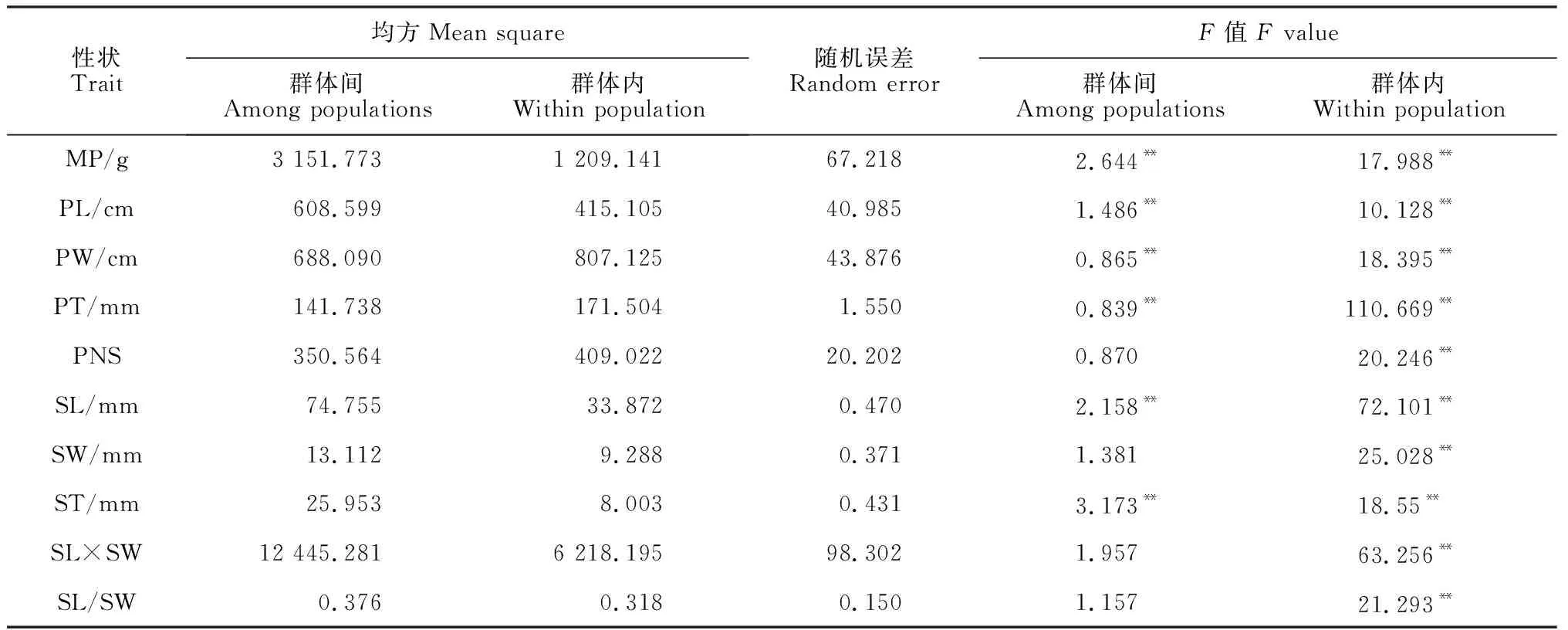
表3 皂荚各群体间及群体内荚果和种子表型性状的方差分析
F值的大小表现了结果可靠性的高低,在一定程度上反映了不同表型性状在群体间和群体内差异情况。因此皂荚群体间种子厚变异最为丰富,荚果宽和荚果厚的变异程度较小;群体内荚果厚的变异指数最高,其变异最丰富,而荚果长的变异最小。
2.3 皂荚表型性状变异分析及群体间表型分化
由表4可见,皂荚群体10个表型性状的变异系数存在较大差异,平均变异系数为21.16%,平均变异幅度为8.76%~41.59%。在各表型性状中,群体间荚果的变异(29.22%)高于种子的变异(11.04%),变异系数最大为荚果质量(41.59%),种子长(9.41%)及种子宽(8.76%)的变异程度较小,说明在皂荚各群体间种子的表型性状更趋于稳定。此外皂荚种实表型性状在不同群体间的变异程度也不同,平均表型变异幅度为16.96%~24.44%,惠水(P6)群体的平均变异系数低于其他各群体,说明其表型多样性较低,而凯里(P4)群体的平均变异系数高于其他群体,说明P4的表型多样性高于其他群体。
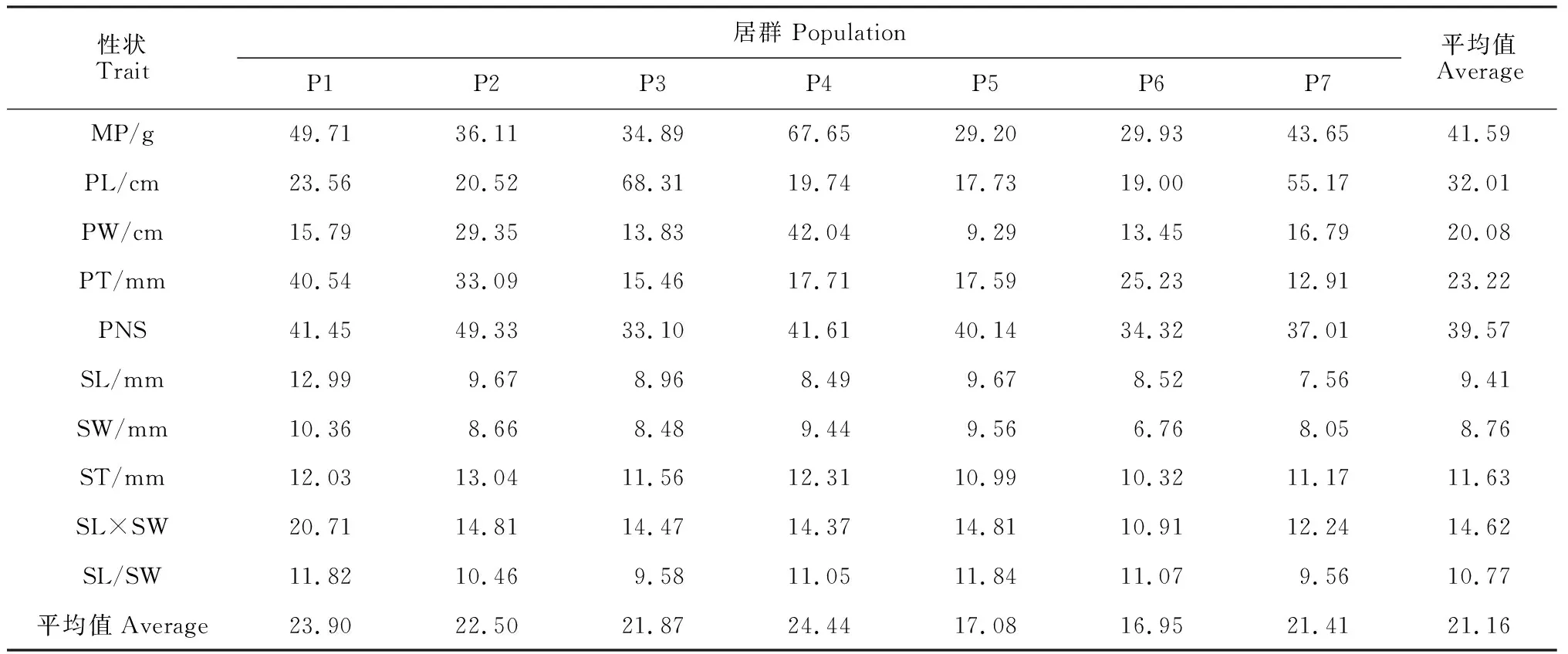
表4 皂荚野生群体各表型性状的表型变异系数
按巢式设计方差分量比组成分析了各方差分量占总变异的比例以及表型分化系数VST。表5表明群体间的方差占9.91%,群体内的方差占17.45%;皂荚形态性状的VST变异幅度为17.8494%~57.5813%,其中种子长、种子厚的VST较大,VST最小的为单荚籽数;皂荚10个形态性状群体间的平均VST为36.51%,说明皂荚在群体间的表型变异的贡献率为36.51%,群体内的贡献率为63.49%,群体内的变异大于群体间的变异,群体间多样性小于群体内多样性,且群体内变异是皂荚种实形态性状变异的主要来源。
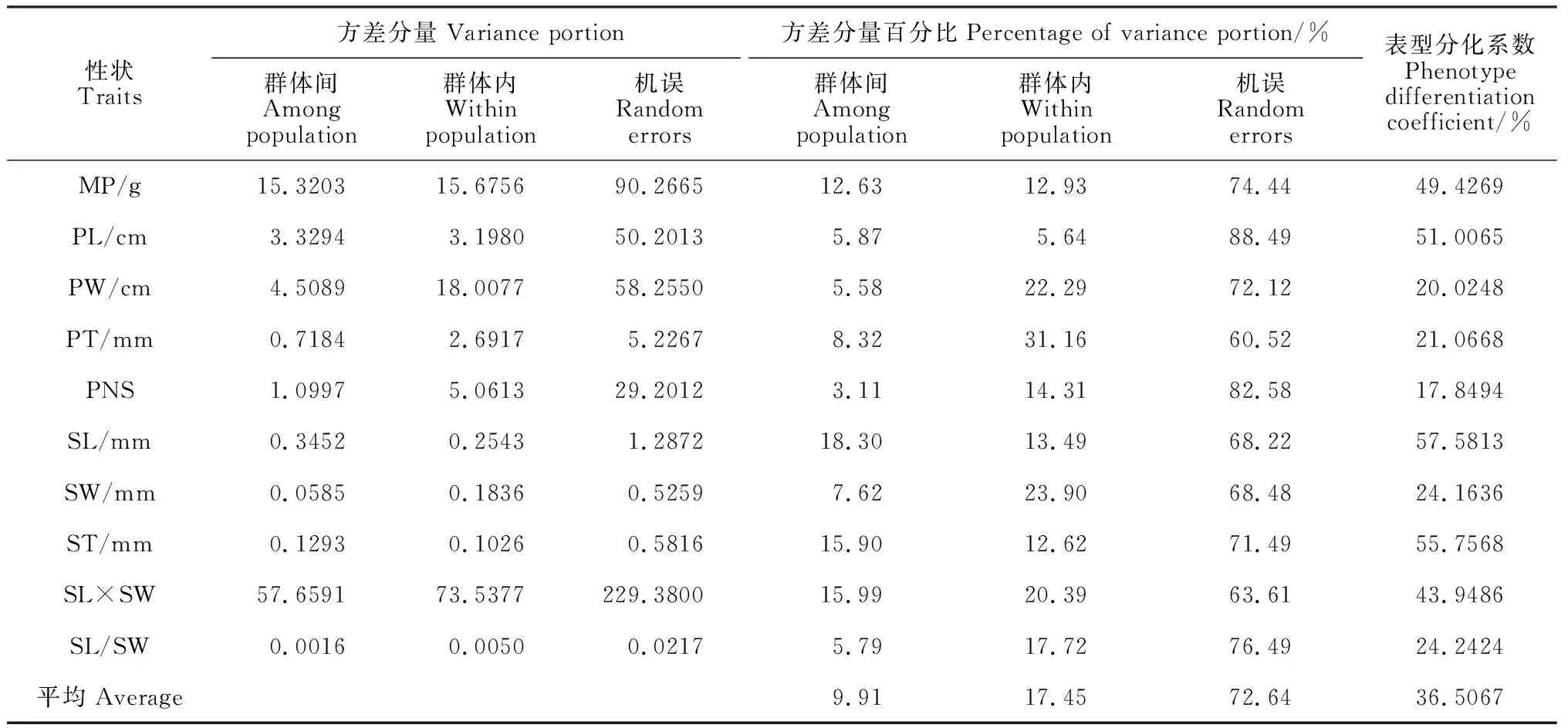
表5 皂荚形态性状的方差分量及群体间表型分化系数
2.4 皂荚各表型性状间的相关性
对群体间所有个体进行种实表型性状间的相关性分析,结果(图1)表明,荚果质量与荚果长、荚果宽、荚果厚、单荚籽数、种子长、种子宽、种子厚、种子长宽积与种子长宽比10个指标之间呈极显著正相关,且荚果质量、荚果长、荚果宽与单荚籽数各指标之间的相关性均达极显著正相关水平,从种子表型性状来看,种子长、种子宽、种子厚与种子长宽积这4者之间互为极显著正相关水平;种子长宽比与种子长呈极显著正相关(r= 0.61),与种子宽呈极显著负相关(r=-0.52)。
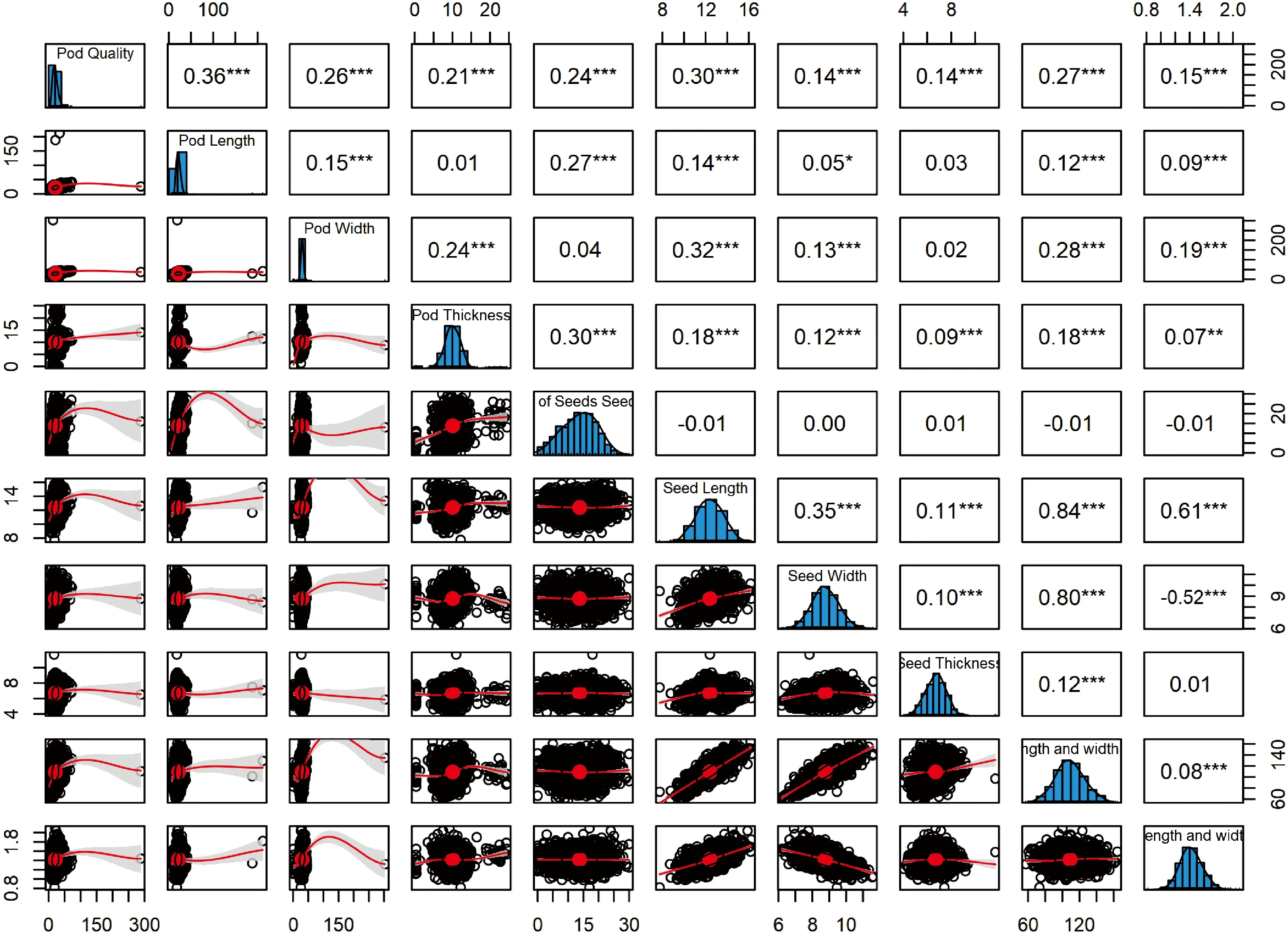
***表示0.001水平显著差异,**表示0.01水平差异显著;*表示0.05水平差异显著图1 皂荚荚果和种子表型性状间相关分析*** mean significant differences at 0.001 level; ** mean significant differences at 0.01 level; *mean significant differences at 0.05 levelFig.1 Correlation analysis of different provenances of pod and seed of G. sinensis
2.5 皂荚各表型性状间的主成分分析
对贵州省7个皂荚群体进行主成分分析,结果(表6)显示:前4个主成分的累积贡献率达69.783%,表明前4个主成分可基本反映皂荚表型性状的大部分信息。
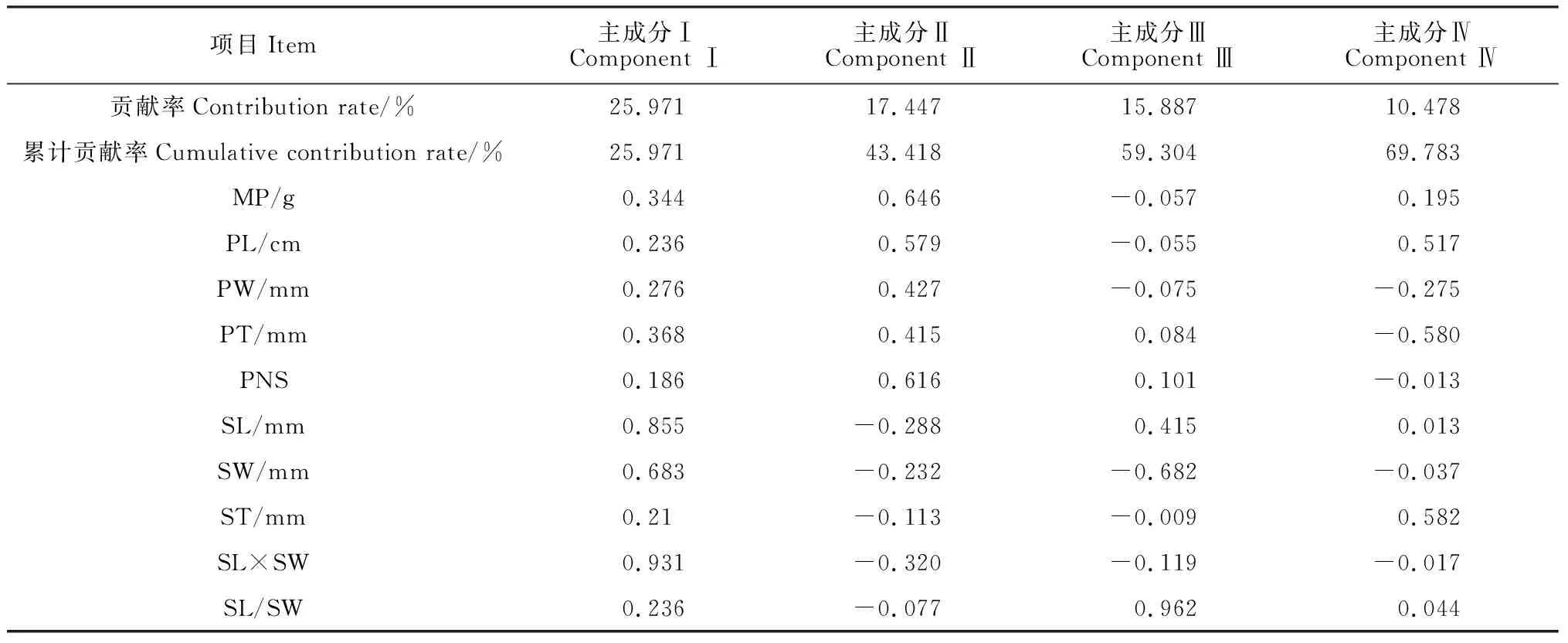
表6 皂荚野生群体各表型性状的主成分分析
第1主成分的贡献率为25.971%,在10个表型性状中,特征向量值较高的性状为种子长(0.855)、种子宽(0.683)、种子长宽积(0.931),说明第1主成分主要反映皂荚种子大小。第2主成分的贡献率为17.447%,其大小主要由荚果重(0.646)、单荚籽数(0.616)决定,说明主要解释了皂荚单个荚果出籽数的大小;第3主成分的贡献率为15.887%,其大小主要由种子长、种子长宽比解释,可命名为皂荚种子形态指数因子;第4主成分的贡献率为10.478%,与荚果长、种子厚相关。
2.6 皂荚各群体的多性状综合评价
依据相关性、主成分分析结果,以皂荚10个种实表型性状为评价指标对贵州省7个群体进行评价,利用公式计算综合得分(Qn)[22],并以Qn值的大小反映不同群体皂荚种实性状综合评价高低。综合得分计算公式为:Qn=0.2597Q1+0.1745Q2+0.1589Q3+0.1048Q4
皂荚各群体种实性状Q值见表7。依据种实性状为选择指标,对7个野生皂荚群体进行综合评价,其排名为:P7>P6>P4>P2>P5>P3>P1,惠水(P7)群体综合表现最优,其荚果质量、荚果长、荚果宽、荚果厚、单荚籽数、种子长、种子宽、种子厚和种子长宽积的平均值(表2)分别高于总体平均值(22.39%、11.59%、6.46%、11.47%、4.21%、1.28%、1.18%、0.40%和3.95%);相较于得分最低的安顺(P1)群体,惠水(P7)群体各表型性状平均值分别高出78.80%、26.48%、21.85%、14.08%、10.25%、8.40%、7.23%、9.02%、15.55%和0.72%。

表7 皂荚野生群体的多性状综合评价
3 讨 论
3.1 皂荚的表型分化情况
表型性状是资源收集、保存、鉴定和利用的基础[23],植物形态指标既具有变异性,又有其稳定性,是个体基于内在基因对环境变化所做出的形态调整,是植物对环境异质性的一种适应,同时受植物自身遗传物质和生态环境两方面的影响[24-25],可在一定程度上反映物种的遗传多样性。关于表型多样性的研究国外早在20世纪60、70年代就已经很成熟,早期主要研究经济和速生类植物的表型多样性,揭示了美国黄松、班克松和美国五针松生长速度的相对变量在种源间大于家系间[26-27]。
本研究对贵州省7个皂荚群体进行了表型形态上的相关分析,方差分析结果表明,皂荚种实的10个形态性状在群体内均达到极显著水平(P<0.01),每荚粒数、种子宽、种子长宽积及种子长宽比这4个形态性状在群体间达到极显著水平(P<0.01),与姜英等[28]研究香合欢在不同种源间的种子千粒重、种子长、种子宽、种子厚和长宽比差异显著结果相似,各种实形态性状差异显著,说明皂荚种实形态性状具有丰富的多样性。从各群体的形态性状平均值来看,惠水群体平均值最大,而独山群体的种子性状平均值更大,说明皂荚群体间各形态性状有着明显的差异性,皂荚群体内的变异更高,遗传多样性更丰富。
3.2 皂荚种实性状表型变异丰富
变异系数包括表型变异系数和遗传变异系数,表型变异系数直接反映表型性状的变异程度[29],间接表明表型多样性的丰富度,即变异系数越大,其性状变异程度越高,表型多样性越丰富。在皂荚10个表型性状中,平均变异系数为21.16%,平均变异幅度为8.76%~41.59%,群体间荚果的变异(29.22%)高于种子的变异(11.04%),说明种子的稳定性好,变异系数最大为荚果质量(41.59%),而种子长(9.41%)及种子宽(8.76%)的变异程度较小,不同群体间荚果的差异较大,种子的表型性状更趋于稳定,这与李伟等[19]的研究结果相似。本次采集的贵州省7个皂荚群体中,凯里群体的平均变异系数最大,其皂荚种实表型多样性最丰富,独山群体的变异系数较低,其表型多样性程度较低,这与独山群体种子性状表达更高相符合。
群体内的多样性是种内多样性的重要组成部分[30],本研究发现皂荚群体内的变异(63.49%)大于群体间的变异(36.51%),说明群体间的多样性低于群体内的多样性,变异来自于群体内,与兰彦平[31]、李伟[19]的研究结果一致,本研究中皂荚的平均分化系数为36.51%,低于砂生槐(Sophoramoorcroftiana)(39.25%)、国槐(SophorajaponicaL.) (53.11%)、香合欢(Albiziaodoratissima)(47.36%),高于紫荆 (CerischinensisBge.)(32.3%)[32-35],与以上植物相比,皂荚群体间的表型分化系数处于中等水平。
3.3 皂荚群体间各表型性状的相关性分析
相关分析是对两个或两个以上有相关性的变量进行分析,从而确定变量之间的相关程度[36],本研究中皂荚的10个表型性状具有一定程度的相关性,荚果质量与荚果长、荚果宽、荚果厚、单荚籽数、种子长、种子宽、种子厚、种子长宽积与种子长宽比10个指标之间呈极显著正相关,荚果质量的大小是由荚果多个性状指标决定,从种子表型性状来看,种子长、种子宽、种子厚与种子长宽积这4个之间互为极显著正相关水平,当种子的长宽指数越大时,其种子的形态大小也更大,从而荚果的表现也越长、越宽,在后续相关育种工作中,可以通过遗传改良荚果的性状从而让种子形态更满足生产需要。
3.4 皂荚群体的主成分分析及综合评价
皂荚荚果形态多样、表型性状较多,良种选育不明确,利用主成分分析可考察多个表型性状间的相关性,能在尽可能保留原有形态性状信息的前提下,利用较少的综合指数代替原先较多的变量,从而简化形态性状的分类工作[37]。在本研究中,前4个主成分的累积贡献率达69.783%,表明前4个主成分可基本反映皂荚表型性状的大部分信息。也是反映皂荚表型性状差异的主要因子,可作为皂荚多样性保护、性状改良及选育中主要的观测性状。依据相关性、主成分分析结果,以皂荚10个种实表型性状为评价指标对贵州省7个群体进行评价,惠水县群体的综合表型性状最好。在今后的皂荚优株遗传改良工作中,优良个体和单株的选择改良是一条有效的途径,能够加速皂荚选育的进程。
综上所述,贵州省皂荚野生群体的表型性状群体间及群体内都存在丰富的遗传多样性。植物的表型性状存在一定的环境可塑性,是其适应环境因子的一种表现[38],在皂荚野生资源的保护过程中,可选取表型性状差异较大的群体进行重点观测取样,并依据育种目标筛选适宜的育种材料,改良不同的荚果形态达到育种目的。
有关植物遗传多样性的研究方法,从表型、生理生化发展到分子水平。表型性状具有直观、方便、快速等特点,但是从表型研究物种资源的多样性具有片面性[39]。本研究从皂荚10个表型性状作为切入点,研究分析了其多样性水平,在之后的工作中,可结合表型多样性从分子水平进行深入研究,明确贵州省皂荚资源的内在遗传多样性水平。
