贵州尾矿区野生豆科牧草根瘤菌及其抗性测定
2023-01-03王海瑾曾庆飞韦兴迪王佳楚函
王海瑾, 曾庆飞, 韦兴迪, 王佳楚函, 韦 鑫, 陈 超*
(1. 贵州大学动物科学学院, 贵州 贵阳 550025; 2. 贵州省农业科学院草业研究所, 贵州 贵阳 550006)
自20世纪中叶以来,随着世界工农业的快速发展、重金属矿产资源和化石燃料的大规模开发、化肥与农药的持续使用,大量的重金属化合物进入农用土壤。进入土壤的重金属难以被土壤微生物降解,致使土壤重金属污染具有隐蔽性、长期性、累积性以及不可逆转等特点,自然净化过程十分缓慢,土壤重金属污染已经变成了一个普遍而持久的环境问题[1]。重金属污染土壤的修复方法包括物理修复、化学修复和生物修复三类技术,实践证明,其中的生物修复是行之有效的绿色而经济的修复方法[2]。生物修复主要分为植物修复、微生物修复、“植物-微生物”联合修复三种类型,其中植物-微生物联合修复能够弥补单纯植物修复与微生物修复的不足,能够显著提高生物修复重金属的能力,已成为土壤重金属污染生物修复研究的热点之一[3]。
豆科植物与根瘤菌形成的共生固氮系统也属于植物与微生物之间的特定联合体系。近年来的研究发现,具有固氮能力的部分根瘤菌同时也具有重金属的吸附固定与转导功能,尤其当根瘤菌与豆科植物形成共生根瘤后,所建立的“豆科植物-根瘤菌”共生系统对重金属污染环境的修复效果更为显著,被认为是比传统的“植物-植物”根际促生菌(PGPR)联合修复系统更具潜力的共生修复体系[4],早在1983年,Rothen在研究洗矿地带土壤中Cd,Pb,Zn对豆科植物根瘤菌固氮能力的影响时发现,白三叶(Trifoliumrepens)根瘤菌的固氮能力几乎不受重金属的影响,即使在Cd浓度为216 mg·kg-1、Zn浓度为200 mg·kg-1、Pb浓度为300 mg·kg-1的污染土壤中,白三叶根瘤菌的固氮能力也只是略有下降[5]。大量研究已经证实,在豆科植物对土壤重金属的修复过程中,根瘤菌起着非常重要的协同作用。根瘤菌不仅能让豆科植物在重金属污染土壤中正常生长,缓解高浓度重金属与类金属元素对植物所造成的环境压力[6],而且还能通过将根际土壤中的重金属吸附到根表或转化成难溶形态,或者直接吸收进入根内,强化植物对重金属的固定效果[7];甚至将土壤重金属通过根系富集到地上部的植物组织中,增加豆科植物对土壤重金属的提取效率[8]。
贵州属于矿产资源大省,已经开采过的铅锌矿以及正在开采的锰矿、磷矿等不可避免的会对周边环境造成不同程度的土壤重金属污染,利用“豆科植物-根瘤菌”共生体系修复被污染的尾矿区及耕地土壤具有重要的生态价值和现实意义。然而,共生修复体系的利用首先必须依赖于共生固氮根瘤的有效形成,共生根瘤的形成同时受到细菌、植物及生态地理环境三个因素的共同影响,作为宿主的豆科植物自身要适应环境的压力,而与其结瘤的根瘤菌则面临共生宿主和环境的双重选择,结果只能是既能与宿主植物识别又能适应当地环境的菌株才能与匹配植物形成根瘤固氮共生系统[9]。鉴于此,本研究从生长在贵州地区重金属尾矿地的天然豆科牧草中分离保存根瘤菌株,对尾矿污染地的共生根瘤菌进行系统发育和区系分析,在认识贵州尾矿区野生豆科牧草根瘤菌种类分布特征和遗传多样性的基础上,根据尾矿地存在的主要重金属种类,从中筛选对主要几种重金属具有较强耐受能力的抗性菌株,目的在于挖掘利用本地尾矿区的共生根瘤菌资源,使筛选出来的抗性根瘤菌既能定殖共生于本地宿主豆科牧草根系,又能适应贵州岩溶山区的喀斯特环境,为下一步筛选对复合重金属污染土壤具有较强修复能力的“豆科牧草-根瘤菌”优势组合类型奠定共生微生物基础,为采用适合在本地区推广使用的“豆科植物-根瘤菌”共生体系治理修复重金属污染环境提供可用的物质和技术支持。
1 材料与方法
1.1 根瘤样品
1.1.1根瘤采集地点 野生豆科牧草根瘤样品于2021年分别采自贵州省铜仁市松桃苗族自治县的锰矿区、毕节市赫章县的废弃铅锌矿区和六盘水市钟山区的铅锌矿区。18个尾矿采样地的详细信息见表1。
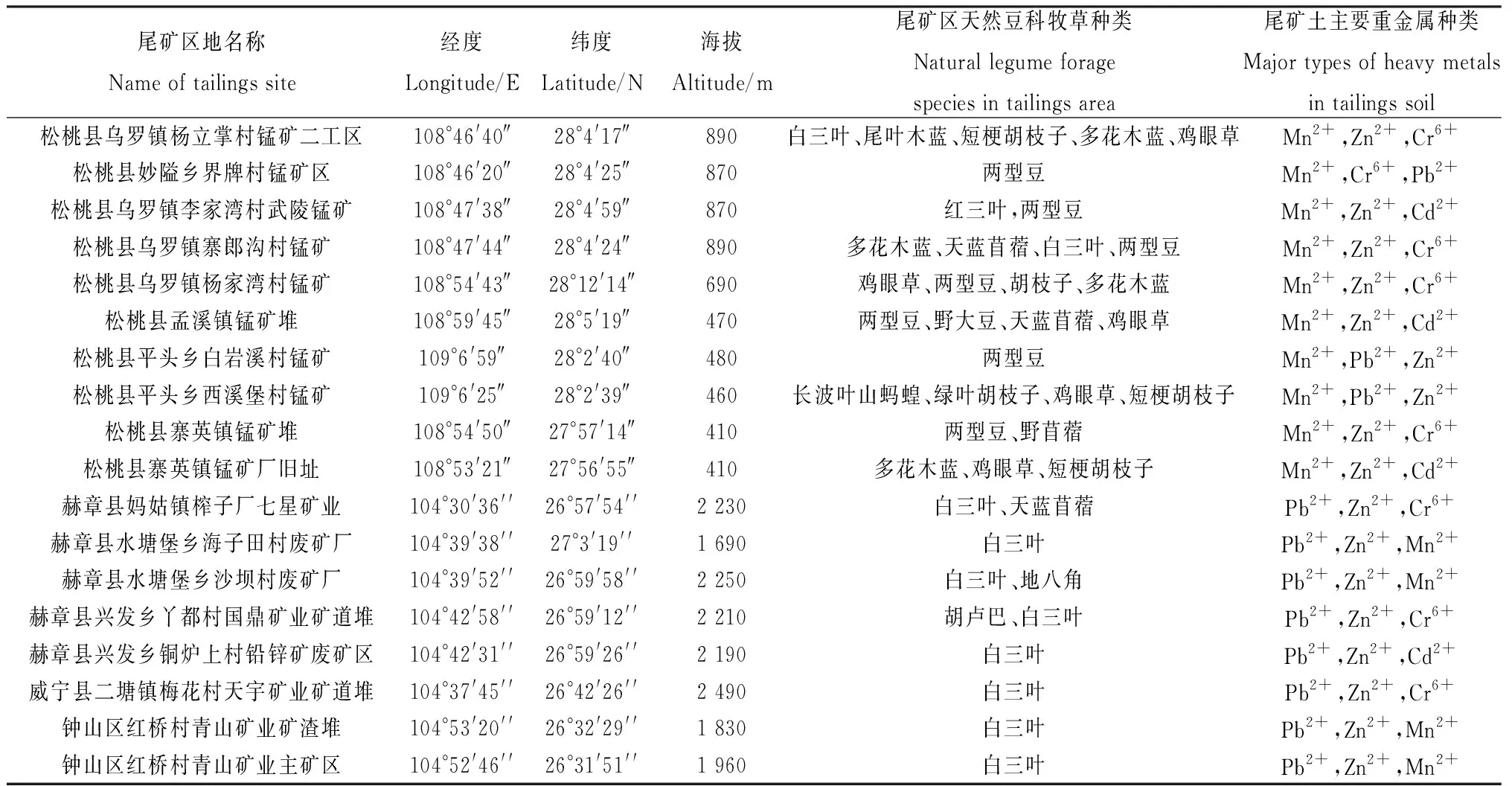
表1 贵州尾矿区天然豆科牧草根瘤样品采集地信息表Table 1 Information of collecting sites of natural legume herbage nodule samples in Guizhou tailings area
1.1.2根瘤样品采集及保存方法 选取各尾矿区已被污染且土壤已经变色的地块作为采样点,挑选在尾矿地生长正常的野生豆科牧草植株,以使选择的采样地和野生豆科牧草在各自的尾矿区具有代表意义。挖取植物根部,采集新鲜健壮、颜色鲜艳的成年根瘤,剪时稍带一段根,编号并记录采集时间、地点、生境、根瘤形状、颜色、大小、着生位置、寄主豆科牧草种类等内容,将采集样品保藏于装有变色硅胶的离心管中密封,4℃冰柜中冷藏保存备用。
1.2 根瘤菌株的分离纯化
选取个体相对较大、颜色新鲜的根瘤,干燥的根瘤在无菌水中浸泡至吸胀,新鲜的根瘤用无菌水冲洗干净后,立即挑取10~20颗健康饱满的单个根瘤,先用95%乙醇浸泡5 min后再用0.1%HgCl2浸泡5 min,其间轻轻摇晃3~5次,无菌水冲洗5~10次后放入无菌研钵中,加入1 mL无菌水,磨棒研磨1 min,研磨液用无菌枪头转至加有刚果红的酵母甘露醇琼脂(Yeast mannitol agar,YMA)培养平板上,涂布均匀后放入培养箱内,28℃恒温倒置培养3~7 d,3 d后作每天1次的连续观察。挑取菌落圆形、透明或半透明、边缘整齐不吸色、湿润黏稠、表面光滑隆起的典型单菌落,平板划线接种至另一新的YMA平板上,28℃恒温倒置培养,长出菌落后挑取单菌落进行革兰氏染色镜检,若不纯,进一步纯化,确保获得纯化菌株。将纯化菌株接种至YMA液体培养基中,28℃振荡培养7 d后用30%甘油保种,-80℃超低温冰箱中冻存。
1.3 根瘤菌株的鉴定与区系分析
1.3.1根瘤菌株的形态学鉴定 参照《微生物分类学》中原核细菌形态学鉴定方法[10],对纯化后革兰氏染色为红色的阴性(G-)菌株进行菌体形状、荚膜和鞭毛有无等形态特征的观察。菌体形状在菌株革兰氏染色鉴定时一并观察记录;采用石碳酸复红染色液初染、黑色素复染方法油镜观察菌体荚膜的存在,如有荚膜,在灰色或黑色背景中,菌体呈红色,荚膜无色;采用硝酸银鞭毛染液染色法,油镜观察菌体鞭毛的存在及着生位置,菌体呈深褐色,体表鞭毛呈浅褐色。
1.3.2根瘤菌株的16S rRNA基因序列比对 采用杭州倍沃医学科技有限公司的细菌基因组DNA抽提试剂盒提取各菌株的总DNA,选用7f-1540r细菌16S rRNA基因扩增通用引物对扩增各菌株的16S rDNA片段;选用质粒PMD18-T载体与纯化回收的PCR产物连接后转化大肠杆菌DH5α;利用生工生物工程(上海)股份有限公司的质粒提取试剂盒提取阳性转化子的质粒DNA,酶切验证后送上海生工成都测序实验室测序。在NCBI(www.ncbi.nlm.nih.gov)网站中将测得的各菌株序列分别进行Blast在线比对,并从GenBank数据库中下载与各菌株序列相近的模式菌株的16S rDNA序列,将待分析菌株序列与模式菌株序列选用DNAMAN软件进行多重比对,结合各菌株的形态特征,确定其种属分类地位。
1.3.3根瘤菌株的区系分析 根据分离菌株的分类鉴定结果,对分离到的贵州尾矿区野生豆科牧草根瘤菌的种类组成、分布频率以及优势种类进行系统分析。分布频率采用微生物区系分析普遍使用的相对频率计算公式分别计算[11]:相对频率(%)=(有某菌种发生的各样品数/有菌种发生的总样品数)×100。
1.4 根瘤菌株的系统发育分析
使用Clustal X(1.8)软件包进行各菌株种类16S rRNA基因的多序列匹配排列,进化距离运用MEGA4软件包中的Kimura2-Parameter Distance模型计算;采用贝叶斯软件进行聚类分析,PAUP和TreePuzzle软件构建系统发育树,并进行系统发育分析。
1.5 重金属抗性根瘤菌株的筛选
1.5.1供试菌株 由于菌株的抗性检测需要设立5种重金属离子、各3个浓度梯度的试验处理,工作量较大,因此本研究从分离纯化并经鉴定确为根瘤菌的保存菌株中,优先选择种类不同、宿主牧草地域不同且在YMA平板上菌落生长良好的根瘤菌作为抗重金属试验的供试菌株。
1.5.2重金属平板筛选 根据贵州尾矿地重金属的分布特点,选择其中含量较高、分布较广的铅、锌、铬、锰、镉五种重金属用于配制筛选平板。参照抗性根瘤菌筛选最常使用的重金属离子的浓度范围[12],分别配制Pb2+,Zn2+,Cr6+,Mn2+,Cd2+五种重金属离子各1 mmol·L-1,2 mmol·L-1,3 mmol·L-1的YMA平板,接种挑选出的供试菌株,每个菌株、每种重金属及每个梯度浓度接种3个平板作为3个重复,接种无重金属盐的YMA平板作对照,28℃恒温箱中倒置培养,分别在第2 d、第5 d、第7 d、第10 d、第14 d连续5次观察菌落的生长情况,记录各菌落的直径大小,计算各菌株菌落直径的平均值。
1.6 数据统计分析
采用Excel 2010整理数据;使用SPSS 18.0软件进行统计学分析。
2 结果与分析
2.1 分离获得的根瘤菌菌株及其形态特征
从55份尾矿区野生豆科牧草根瘤样品中,经分离纯化和染色鉴定,共培养出83个革兰氏阴性(G-)纯化菌株。分别观察各G-菌株的菌体形态特征,显示菌体形态均为杆状,长短各不相同,菌体表面有荚膜或无荚膜,但均有鞭毛着生,着生位置端生、侧生或周生,符合根瘤菌的基本形态特征。
2.2 根瘤菌株的种类鉴定与区系分析结果
83个G-保存菌株的总DNA采用细菌基因组DNA抽提试剂盒提取,琼脂糖凝胶电泳均检测到23 kb左右的条带。选用16S rDNA序列扩增通用引物对P1—P6,全部菌株都扩增出约1.5 kb的序列片段。根据菌株的形态鉴定和16S rDNA的分子鉴定,分离培养的尾矿区野生豆科牧草根瘤菌的种属鉴定结果见表2。结果表明,在83个G-菌株中共鉴定出根瘤菌71株,其余12个菌株属于在根瘤内与根瘤菌一起共生的根瘤内生细菌。
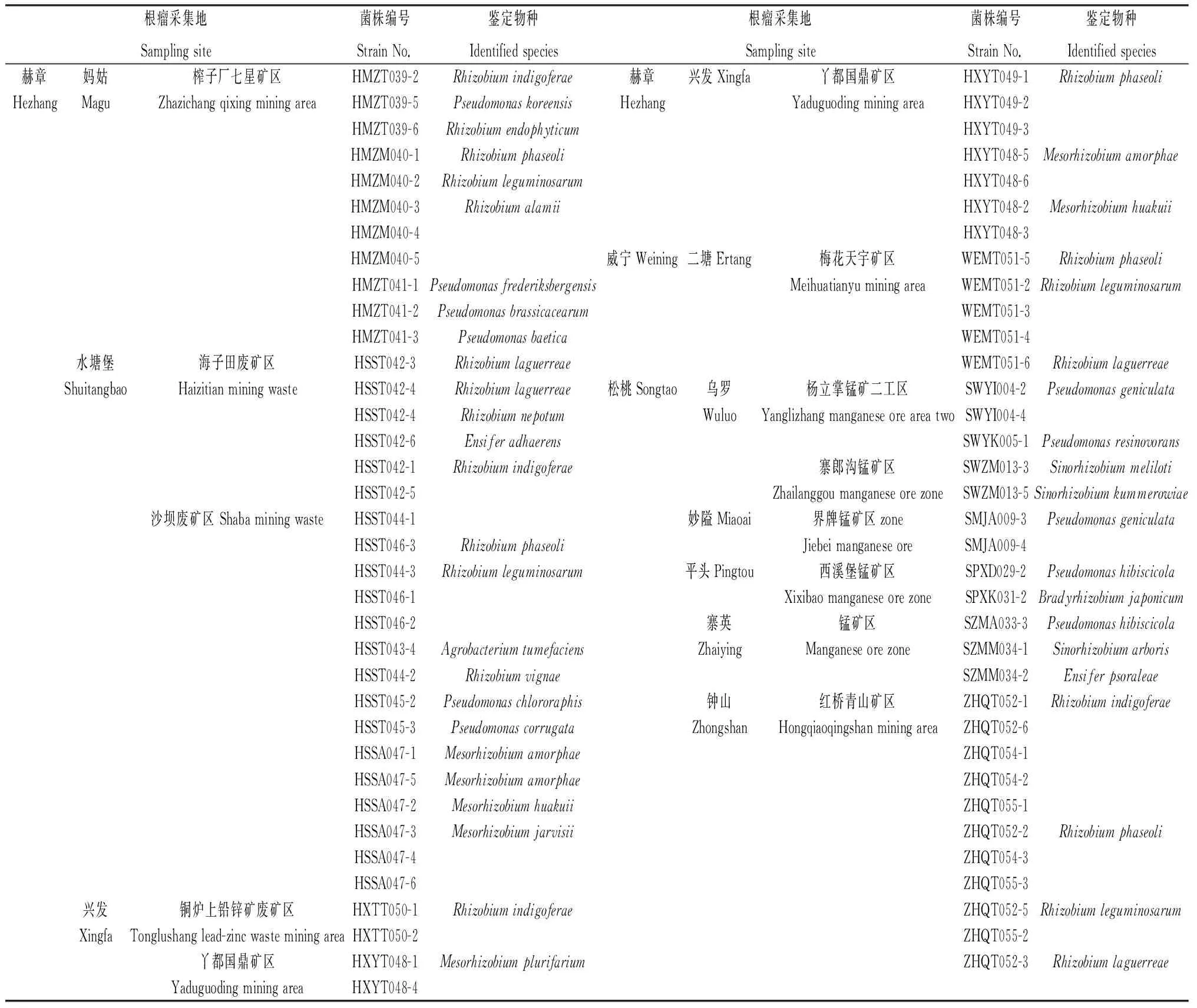
表2 基于16S rDNA序列比对的分离根瘤菌株分子鉴定结果Table 2 Taxonomic identification results of rhizobial strains based on 16S rDNA sequence alignment
将鉴定出的根瘤菌种类进行区系分析(表3),结果显示,分离获得的贵州尾矿区野生豆科牧草根瘤菌分属于7个属28个种。7个属分别为:根瘤菌属(Rhizobium)、慢生根瘤菌属(Bradyrhizobium)、中慢生根瘤菌属(Mesorhizobium)、中华根瘤菌属(Sinorhizobium)、剑菌属(Ensifer)、土壤杆菌属(Agrobacterium)和假单胞菌属(Pseudomonas)。其中根瘤菌属的分布频率最高,达54.93%,为优势属;假单胞菌属和中慢生根瘤菌属次之,分别为18.31%和16.90%;28个种中的木兰根瘤菌(R.indigoferae)分布频率最高,达到15.49%,为优势种;菜豆根瘤菌(R.phaseoli)和豌豆根瘤菌(R.leguminosarum)次之,同为12.68%。

表3 贵州尾矿区野生豆科牧草根瘤菌区系分析结果Table 3 Results of rhizobia flora analysis of wild legume forages in Guizhou area near tailing dam
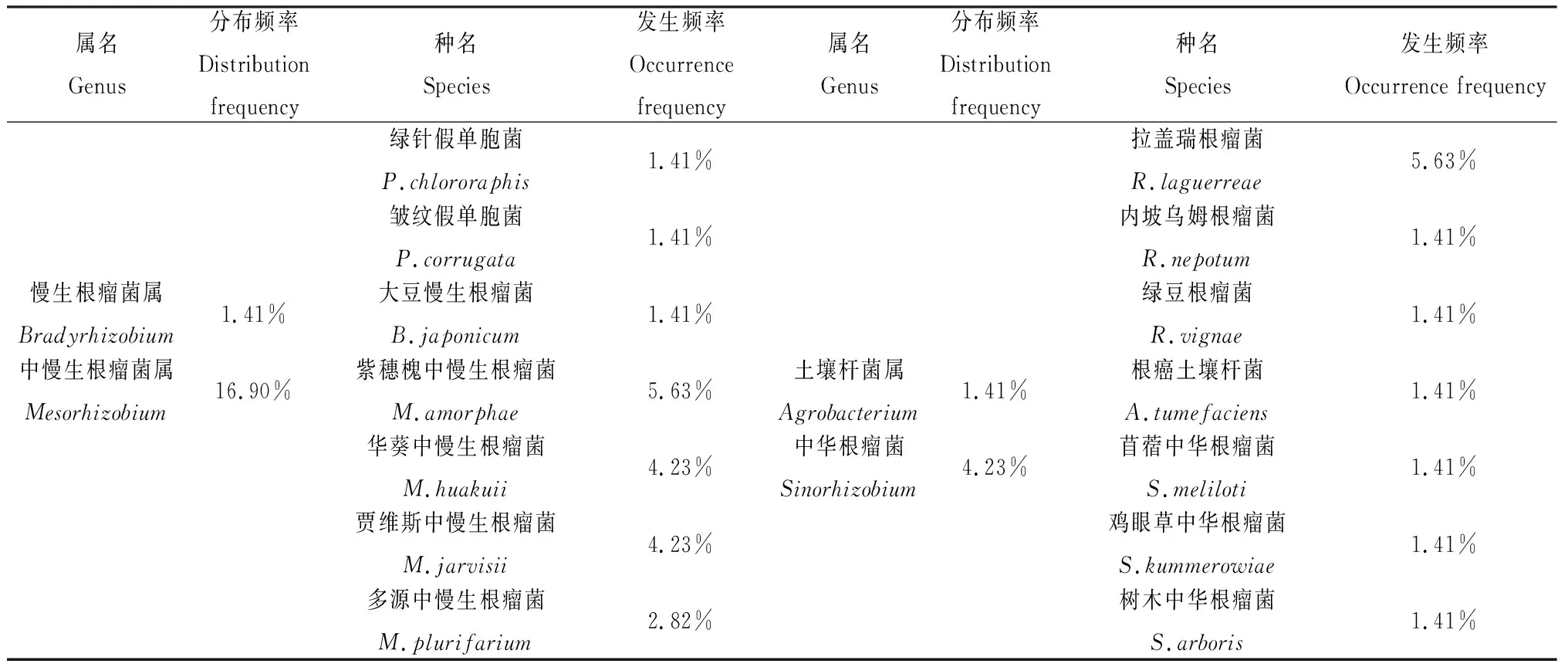
续表3
2.3 根瘤菌株的系统发育分析
根据鉴定出的28种根瘤菌的16S rDNA序列,构建的贵州尾矿区天然豆科牧草根瘤菌的系统发育树如图1。28种根瘤菌分为5大类群18个分支,而其中有3个分支是由不同属的种类聚成的,分别是α-变形菌纲中的绿豆根瘤菌(Rhizobium.vignae)和多源中慢生根瘤菌(Mesorhizobium.plurifarium)、内坡乌姆根瘤菌(R.nepotum)和根癌土壤杆菌(Agrobacterium.tumefaciens)、中华根瘤菌(Sinorhizobium.arboris)和补骨脂剑菌(Ensifer.psoraleae);另外,γ-变形菌纲内假单胞菌属中的3个种类:油菜假单胞菌(Pseudomonasbrassicacearum)、皱纹假单胞菌(P.corrugata)和食樹脂假单胞菌(P.resinovorans)独立形成一个分支;α-变形菌纲内慢生根瘤菌属中的大豆慢生根瘤菌(Bradyrhizobiumjaponicum)独立形成一个分支;剑菌属中的粘着剑菌(Ensiferadhaerens)独立形成一个分支。这说明贵州尾矿区天然豆科牧草根瘤菌具有复杂的遗传背景和明显的丰富多样性。
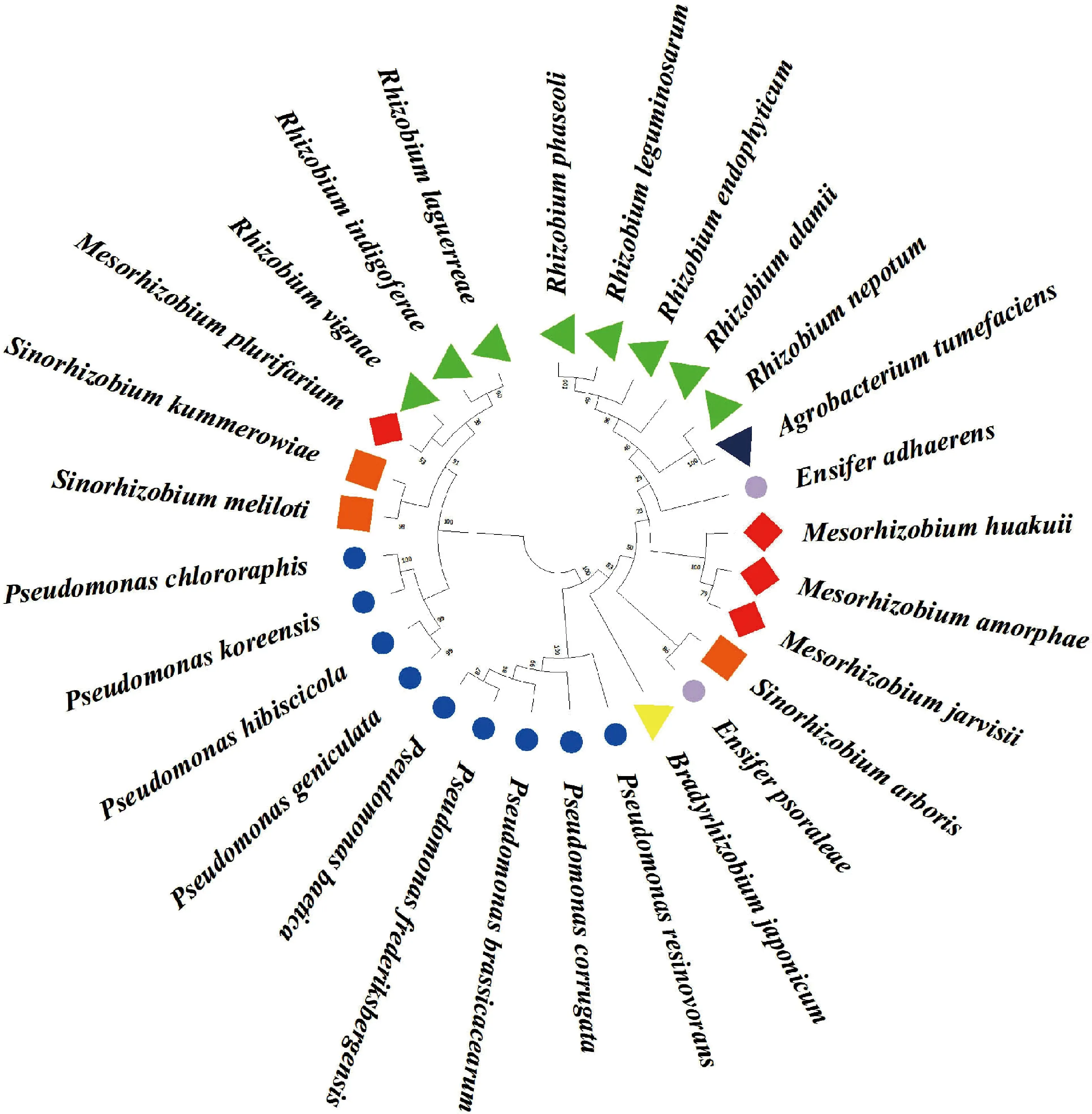
图1 基于16S rDNA序列构建的28种根瘤菌的系统发育树Fig.1 Phylogenetic tree of 28 species of rhizobia based on 16S rDNA sequence
2.4 重金属抗性根瘤菌株的筛选结果
根据贵州尾矿区野生豆科牧草根瘤菌的分离鉴定结果,按照不同地域、不同种类且菌落生长良好的挑选原则,共选出17个菌株作为重金属抗性检测筛选对象,菌株名分别为:HMZT039-2,HMZT039-6,HMZM040-1,HMZM040-3,HSST042-1,HSST042-6,HSST044-1,HSST044-2,HSST046-1,HSSA047-1,HXYT048-1,HXYT049-1,HXTT050-1,WEMT051-2,ZHQT052-1,ZHQT054-1,ZHQT055-1。
将17个供试菌株分别接种5种重金属离子3个浓度梯度的YMA平板后,分5次观察记录各平板上菌落的生长情况,记录各菌落的直径大小,计算各菌株菌落直径的平均值。研究证明,根瘤菌在含重金属盐的平板上生长相对较慢,而且在3 mmol·L-1盐浓度的平板上能正常生长的根瘤菌属于高抗菌株[12],因此在该浓度平板上第14 d的菌落生长直径是较为有效的筛选指标。本筛选实验中最终第5次(第14 d)的观察记录结果见表4。结果显示,17个供试菌株在不含重金属盐的YMA平板上都能正常生长,但在某些重金属盐平板上无菌落形成。根据3个重复平板均有菌落生长为判断标准,得出的具体测定结果为:菌株HMZM040-1和ZHQT055-1可耐受3 mmol·L-1的Mn2+和Cr6+,2 mmol·L-1的Zn2+;ZHQT054-1可耐受3 mmol·L-1的Mn2+和Cr6+,2 mmol·L-1的Zn2+,1 mmol·L-1的Pb2+;HXYT048-1和HXYT049-1可耐受3 mmol·L-1的Mn2+,Cr6+和Zn2+,1 mmol·L-1的Pb2+;HSST044-1,HSST046-1和WEMT051-2可耐受3 mmol·L-1的Mn2+和Cr6+,2 mmol·L-1的Pb2+和Zn2+;HMZT039-2,HSST044-2和HSSA047-1可耐受3 mmol·L-1的Mn2+,Cr6+和Zn2+,2 mmol·L-1的Pb2+;HMZM040-3可耐受3 mmol·L-1的Mn2+,Cr6+,Pb2+和Zn2+;HXTT050-1可耐受3 mmol·L-1的Mn2+,Cr6+和Zn2+,2 mmol·L-1的Cd2+,1 mmol·L-1的Pb2+;HSST042-1可耐受3 mmol·L-1的Mn2+,Cr6+,Zn2+和Cd2+,1 mmol·L-1的Pb2+;ZHQT052-1可耐受3 mmol·L-1的Mn2+,Cr6+,Pb2+和Zn2+,1 mmol·L-1的Cd2+;HSST042-6可耐受3 mmol·L-1的Mn2+,Cr6+,Pb2+和Zn2+,2 mmol·L-1的Cd2+;菌株HMZT039-6能耐受3 mmol·L-1的全部5种重金属离子。
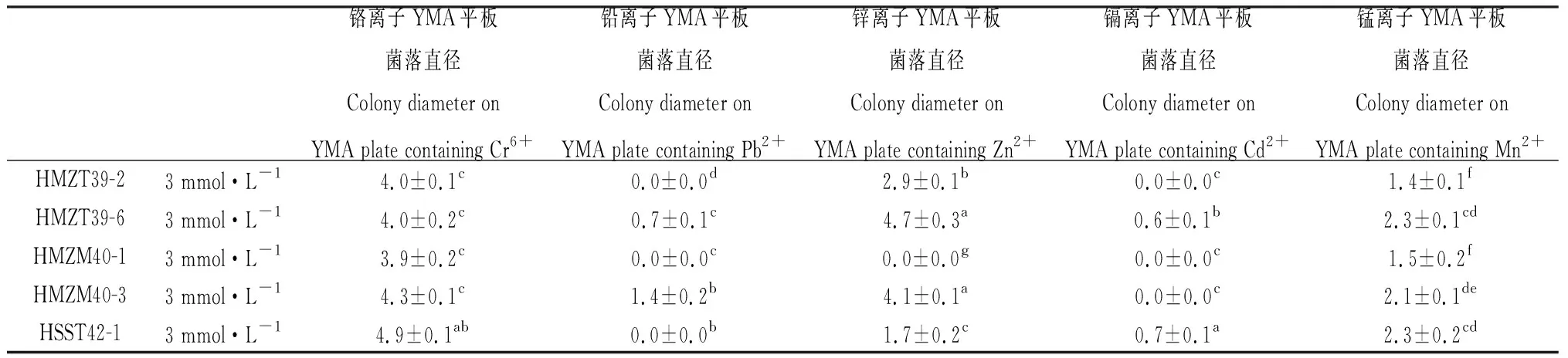
表4 接种后第14 d供试菌株在重金属平板上菌落生长直径测定结果Table 4 Measurement results of colony growth diameter of tested strains on heavy metal plates when inoculated for 14 days 单位:mm
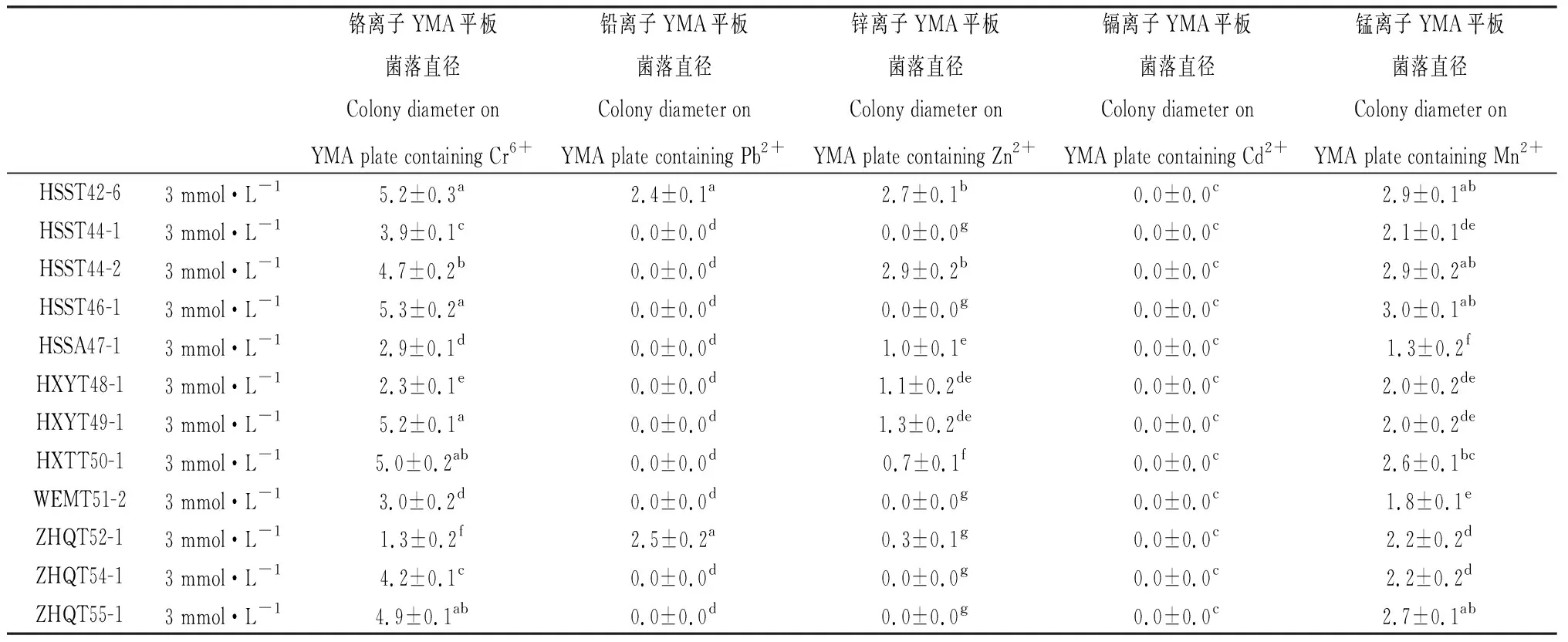
续表4
归纳起来,测定结果显示出下列4个特征:①部分菌株对某种重金属没有抗性的,就算在低浓度(1 mmol·L-1)的平板上也一直没有菌落生长;②部分菌株对某种重金属只能在1 mmol·L-1低浓度的平板上生长,有的菌株对某种重金属可以在2 mmol·L-1中浓度的平板上生长,但不能耐受3 mmol·L-1的高浓度;③全部供试菌株都能耐受3 mmol·L-1的Mn2+和Cr6+,2~3 mmol·L-1的Zn2+,17个菌株中的15株都能耐受1~3 mmol·L-1的Pb2+,能耐受Cd2+的菌株相对较少,17个供试菌株中只有5株;④所有菌株都能耐受至少3种不同种类、不同浓度的重金属离子。
根据耐受重金属的种类数量及耐受浓度来划分,筛选出的优良菌株抗复合重金属的能力大小依次排序为:HMZT039-6>HSST042-6>HSST042-1,ZHQT052-1>HXTT050-1>HMZM040-3>HMZT039-2,HSST044-2,HSSA047-1>HSST044-1,HSST046-1,WEMT051-2>HSST042-1>HXYT048-1,HXYT049-1>ZHQT054-1>HMZM040-1,ZHQT055-1。其中,分离自赫章县妈姑镇榨子厂七星矿区白三叶根瘤的HMZT039-6,分离自赫章县水塘堡乡海子田村废矿区白三叶根瘤的HSST042-6和HSST042-1,分离自六盘水钟山区红桥村青山矿业区白三叶根瘤的ZHQT052-1,以及分离自赫章县兴发乡铜炉上村铅锌废矿区白三叶根瘤的HXTT050-1等5个菌株,能耐受供试的全部5种重金属离子,为筛选出的最优抗性菌株。
3 讨论
重金属尾矿区在经过一定时间的自然选择后都会出现不同数量和种类的耐受本土植物,特别是野生豆科植物,并形成能适应矿区重金属环境的“豆科植物-根瘤菌”共生体系,进而形成不同尾矿区豆科植物根瘤菌特定的区域分布特征[13]。2006年,朱毓华等[14]对分离自陕西太白金矿废弃地8种豆科植物的108株根瘤菌进行唯一碳源、氮源利用等151项表型性状分析,发现供试菌株在85%的相似水平上聚成5个类群,显示分离自废弃地同一生境下不同寄主植物的根瘤菌存在着明显的表型多样性;2007年,余建福等[15]又对分离自陕西太白金矿尾矿区豆科植物的55株根瘤菌进行种群分析,发现它们分为Rhizobium,Sinorhizobium,Mesorhizobium,Agrobacterium4个群和1个未知群,暗示尾矿区生态环境的差异也会导致根瘤菌不同的多样性特征。2008年,位秀丽[16]通过多相分类方法和系统发育分析,对西北尾矿区天蓝苜蓿根瘤菌的多样性进行了研究,发现69株供试天蓝苜蓿根瘤菌被分别划分在中华根瘤菌属、根瘤菌属和土壤杆菌属,其中64株属于中华根瘤菌属,表明同一尾矿地与同种豆科植物结瘤共生的根瘤菌具有相对的专一性。2012年,李彪[17]对分离自云南会泽铅锌尾矿3种乡土豆科植物(白三叶、苕子和紫花苜蓿)的49株根瘤菌进行系统发育分析,发现它们在发育树树上属于Rhizobiumsp.,Sinorhizobiumsp.和Bradyrhizobiumsp. 3个系统发育分支,而且按铅锌含量采集地的不同,分别聚集为OTU1,OTU2和OTU3三个群,说明尾矿区豆科植物根瘤菌的遗传多样性还与土壤中重金属的种类与含量存在着相关关系。
本研究在贵州尾矿地采集根瘤样品时发现,不同污染区域内能够自然生长的豆科牧草种类存在着较大差别,而且数量都很有限,这应该是不同地理条件、不同小生态环境以及土壤中不同重金属种类及不同浓度胁迫所形成的结果,因此在取样时不能做到每个矿区每种植物的种类和数量都具有一致性。但是,所选择的采样地一定是明显被污染的地块,污染地块上所有的豆科牧草种类都会被选取,而且每一种牧草都会在5个以上不同的位置挖取根瘤,因而能够代表每一尾矿地域内天然豆科牧草根瘤菌资源的基本情况。在18个尾矿区共采集到15种野生豆科牧草,分离出的根瘤菌分属于7个属28个种,包括已报道的Rhizobium,Sinorhizobium,Mesorhizobium,Bradyrhizobium,Agrobacterium等5个属[18]以及未见报道的Ensife和Pseudomonas2个属的根瘤菌种类,显示贵州尾矿区野生豆科牧草根瘤菌具有明显的丰富多样性。另外,在筛选出的5个优良抗性根瘤菌株中,有4个菌株(HMZT039-6,ZHQT052-1,HSST042-1和HXTT050-1)属于根瘤菌属,只有1个菌株(HSST042-6)属于剑菌属,而其中又有3个菌株(ZHQT052-1,HSST042-1和HXTT050-1)都属于木兰根瘤菌(Rhizobiumindigoferae)。这一分布结果与贵州尾矿区根瘤菌区系分布中根瘤菌属为优势属、木兰根瘤菌为优势种的研究结果具有一致性。
由于长期适应与代谢调节的原因,共生根瘤菌的耐受能力必定与土壤中重金属种类与浓度背景值存在着一定关系。但是在实验条件下筛选抗性菌株时,由于菌体处于非共生的自由生长状态,重金属浓度的设置无需与土壤背景值保持一致。本研究选用的是经检验证实确为有效可行的1~3 mmol·L-1的重金属浓度[12],并选择贵州尾矿土含量相对较多的具有代表性的铅、锌、锰、铬、镉5种重金属进行筛选试验。在优先选择的17个供试菌株中,所有参试菌株都能对较高浓度的Mn2+,Cr6+和Zn2+产生抗性,15个菌株都能耐受一定浓度的Pb2+,5个菌株能单独耐受包括Cd2+在内的5种重金属,这与采样地中大部分尾矿土都含有Mn,Cr,Zn,Pb,少部分含有Cd元素这一背景值具有一定的相关性。这说明,在含重金属的尾矿土壤中,能够与豆科植物共生结瘤的根瘤菌,在一定程度上都已经适应并形成了对本土重金属的耐受能力,这种耐受能力应该与土壤中重金属的种类和含量存在着一定的关系;同时不同菌株之间对不同重金属以及复合重金属的耐受能力客观上还是存在着明显的差异。研究已经证实,在实验条件下,同时存在的2种或2种以上的复合重金属会不同程度的抑制菌株对单种重金属的耐受能力[19]。但是在自然条件下,尾矿土壤中存在的重金属都会是2个以上的不同种类[20],从尾矿区豆科植物根瘤中分离筛选出的根瘤菌势必具有耐受复合重金属的能力,它们更容易在地方环境的豆科植物根系内定殖共生,更有利于在本地区发挥重金属污染土壤的共生修复效应,是一笔可贵的共生微生物资源[21]。
由于根瘤菌必须与宿主豆科植物共生结瘤后才能有效发挥土壤重金属污染的修复作用[22],所以本研究的下一步计划是接着将筛选出的最优抗性菌株回接贵州岩溶山区不同种类的天然适生豆科牧草,进一步筛选对复合重金属污染土壤具有较强修复能力的“豆科牧草-根瘤菌”优势组合类型,为采用适合在本地区推广使用的“豆科植物-根瘤菌”共生体系治理修复重金属污染环境提供可用的物质和技术保证。
4 结论
贵州尾矿区野生豆科牧草根瘤菌主要包括根瘤菌属、慢生根瘤菌属、中慢生根瘤菌属、中华根瘤菌属、剑菌属、土壤杆菌属和假单胞菌属7个属的28个种,其中根瘤菌属是优势属,木兰根瘤菌是优势种,本地尾矿区野生豆科牧草根瘤菌具有特定的丰富多样性。
从分离鉴定的尾矿区野生豆科牧草根瘤菌中,优先选择17个来自不同地域和不同种类且菌落生长良好的菌株作为筛选对象,其中分离自4个不同尾矿区而且都是白三叶根瘤样品的5个菌株(HMZT039-6,ZHQT052-1,HSST042-6,HSST042-1和HXTT050-1)对5种供试重金属离子均有较强的耐受能力,在本地重金属尾矿区的土壤和植被恢复中具有潜在的开发应用价值。
