1株致病性黄疸出血群钩端螺旋体病流行菌株与其同血清群疫苗株的比较基因组学分析
2022-12-28李彬石刚杜宗利辛晓芳叶强徐颖华
李彬,石刚,杜宗利,辛晓芳,叶强,徐颖华
中国食品药品检定研究院 国家卫生健康委员会生物技术产品检定方法及其标准化重点实验室,北京 102629
钩端螺旋体病是由致病性钩端螺旋体(简称钩体)引起的一种急性人兽共患传染病,也是洪涝、地震等自然灾害后需重点监测的主要传染病之一[1]。随着环境卫生、经济条件的改善及钩体疫区高风险人群疫苗接种率的提高,近30年钩体病在中国的发病率和病死率显著下降[2-3]。由于洪水灾害及全球气候变暖等因素,每年仍有不少钩体感染病例的报道,在局部地区甚至出现暴发流行[4-5],严重威胁人类健康和畜牧业发展。
对流行地区易感人群、兽类及抗洪救灾等高危人员进行疫苗接种是预防和控制钩体病最有效和经济的手段。中国目前所用预防钩体病的疫苗是由几种不同血清群灭活钩体混合配制而成的菌体疫苗[3,6]。生产用菌种作为疫苗生产最重要的物质基础直接影响疫苗质量。我国钩体疫苗生产用菌种主要依据20 世纪60~70年代各地流行疫区流行病学和病原学调查结果筛选而来[3,7],为明确目前钩体病流行菌株与沿用至今的生产用菌种间的差异,本研究采用高通量测序技术对1 株我国钩体最主要流行血清群——黄疸出血群菌株进行全基因组测序,并与其相同血清群疫苗株进行比较基因组学分析,明确两者的差异,探讨我国流行钩体基因组变异特点和流行趋势,以期为促进钩体疫苗的改进提供实验依据。
1 材料与方法
1.1 菌株 2005年分离的致病性黄疸出血群钩体流行菌株200502(简称流行株200502)由中国食品药品检定研究院中国医学细菌保藏管理中心保存,其多序列位点(MLST)型为ST17,为我国当前流行的黄疸出血群中两个ST型(ST1和ST17)之一[8]。
1.2 主要试剂 EMJH 培养基购自美国BD 公司;兔血清购自天津市正江现代生物技术有限公司;DNA提取试剂盒购自德国Qiagen 公司;其他化学试剂均为国产分析纯。
1.3 细菌培养及DNA 提取 将流行株200502 接种于含10%兔血清的EMJH培养基,于28 ℃培养箱中培养14 d,8 000 ×g离心20 min,收集菌体,用DNA 提取试剂盒提取菌株的全基因组DNA,置-70 ℃保存。
1.4 全基因组测序及生物信息学分析 由上海人类基因组研究中心采用Illumina 公司solexa 测序技术对基因组DNA 样品进行全基因组测序。应用velvet 1.2.03软件对检测数据进行拼装,结合生物信息学分析软件Glimmer 3.02,并参考SEED蛋白数据库进行基因预测、功能注释及直系同源簇(clusters of orthologous groups,COG)分类。将流行株200502 测序获得全基因组序列与同血清群疫苗株56601(简称疫苗株56601)和波氏基因种代表菌株56142(简称波氏代表株56142)进行比较基因组学分析,包括菌株基因种、配对基因组平均核苷酸一致性(average nucleotide identity,ANI)及共有与特有基因比较分析。以已发表的疫苗株56601 为参考,对流行株200502进行插入缺失(indel)和单核苷酸多态性(single nucleotide polymorphism,SNP)分析,以进一步了解不同菌株间基因多态性。同时为了解黄疸出血群钩体的分子进化发育情况,选取GenBank 已公布的包括疫苗株56601 在内的9株不同基因种的代表性钩体菌株(见表1)进行全基因组水平系统发育进化树的构建和分析。
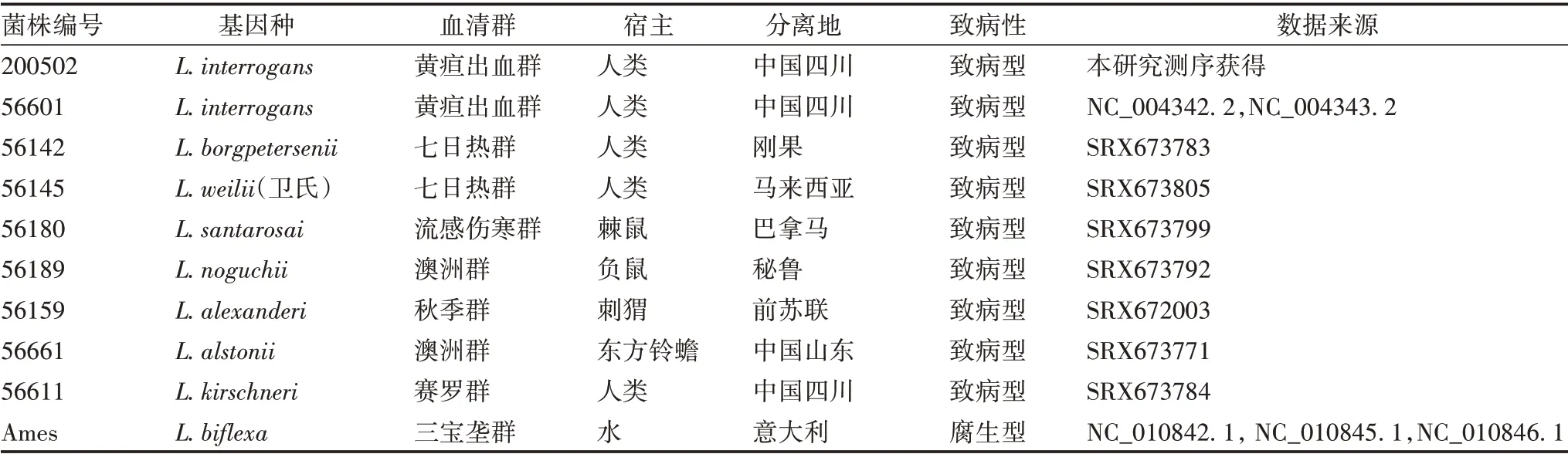
表1 用于构建系统发育进化树的菌株信息Tab.1 Information of strains used to construct phylogenetic evolutionary tree
2 结 果
2.1 流行株200502 的全基因组特征 流行株200502全基因组序列全长为4 543 190 bp,预测所含编码基因3 580 个,平均长度为951 bp,占整个基因组序列的74.9%,鸟嘌呤(G)和胞嘧啶(C)两种碱基之和占总碱基比例(即GC 含量)为36.6%。基因组中非编码基因区域长度为1 139 012 bp,占整个基因组序列的25.1%,GC 含量为30.0%。流行株200502基因组中含有IS1533、ISLbp5和IS1501 3个插入元件。
2.2 流行株200502 全基因组序列基因功能分析 流行株200502全基因组中有1 295个基因无法进行COG分类,2 192 个基因具有明确的生物学功能,与细胞壁、细胞膜、细胞外膜生物合成(216 个)、信号转导机制(185 个)及氨基酸转运和代谢(147 个)相关基因数量排前3位,见表2。另外,有200个(9.1%)未知基因无法获得功能注释,有待后续进一步分析。
2.3 比较基因组学分析 流行株200502 基因组与疫苗株56601 染色体在全基因组水平上表现出较高的共线性和高度的同源性,核苷酸序列呈反向镜面结构,ANI 为99.2%,表明两菌株基因组结构高度相似;流行株200502 基因组的基因种与疫苗株56601相同,均为L.interrogans基因种;与波氏代表株56142 基因组比较,核苷酸序列呈现结构交叉、共线性较少,ANI仅为80.51%。见图1。

图1 黄疸出血群钩体流行株200502、疫苗株56601 和波氏株56142基因组核苷酸序列的共线性分析Fig.1 Collinearity analysis of genomic nucleotide sequences of Leptospira Icterohaemorrhagiae circulating strain 200502,vaccine strain 56601 and L.borgpetersenii 56142
流行株200502 基因组与疫苗株56601 共有3 245个同源基因,与波氏代表株56142共有2 679个同源基因;3 株菌株间共鉴定出2 635 个同源基因。同时,与各自基因组比较,流行株200502、疫苗株56601 和波氏代表株56142 分别有198、232 和618 个基因无同源性,表明为各菌株的特有基因,见图2 和表2。COG 功能注释显示,与疫苗株56601 比较,流行株200502 基因组特有的242 个基因中,74.0%的基因功能未知;在已知功能基因中,与复制、重组、修复和信息转导相关基因数量均为8个。
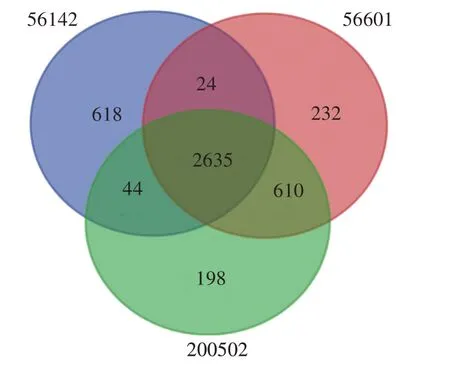
图2 黄疸出血群钩体流行株200502、疫苗株56601 和波氏株56142基因组中的共有和特有基因维恩图Fig.2 Venn diagram of common and unique genes in the genomes of Leptospira Icterohaemorrhagiae circulating strain 200502,vaccine strain 56601 and L.borgpetersenii 56142

表2 黄疸出血群钩体流行株200502与疫苗株56601基因组的COG分析比较Tab.2 Genomic COG analysis of Leptospira Icterohaemorrhagiae circulating strain 200502 and vaccine strain 56601
2.4 indel和SNP分析 流行株200502中核苷酸序列中共发现2 005 个indel 位点和19 799 个SNP 位点,其中编码区有8 485 个同义SNP 位点(syn)和3 484个非同义SNP 位点(nonsyn),非编码区有7 830 个SNP位点。多数SNP和indel位点分散在不同区域和基因中,与菌株的细胞运动、信号转导机制、防御机制等相关,另外,还发现一些已知与钩体致病相关的脂蛋白与外膜蛋白,如脂蛋白LipL32 和LipL21 分别存在1 个和2 个同义SNP 位点,LipL41 含4 个同义SNP 位点和11 个非同义SNP 位点;外膜蛋白OmpL1发现存在31个同义SNP位点和7个indel位点。
2.5 系统发育进化分析 腐生型Ames 为近祖先菌,位于进化树最外围,其他致病型与其进化距离较远,形成两个最大分支,流行株200502 与L.interrogans(56601)、L.noguchii(56611)和L.kirschneri(56189)3个基因种进化亲缘性较近,见图3。
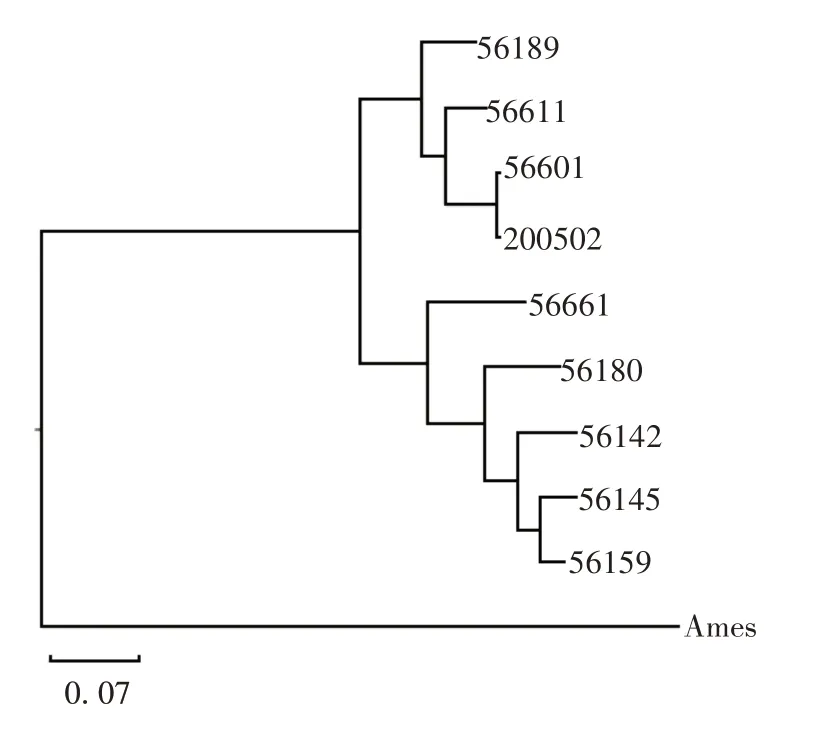
图3 不同钩体基因种的全基因组系统发育进化树分析Fig.3 Analysis of genome-wide phylogenetic evolutionary tree of various Leptospira gene species
3 讨 论
大量国内流行病学研究结果证实,当前我国几个钩体流行地区流行的钩体血清群为黄疸出血群[9-11]。近年来,国内外研究人员积累了大量致病性钩体的基因组信息[12-13],同时在临床上也有采用高通量测序技术进行钩体病诊断的报道[4,14]。本研究采用高通量测序技术和系统生物信息学分析、解读了我国近期分离的1 株致病性黄疸出血群菌株的全基因组序列,充实了基因数据库,全面注释了基因组中与钩体生长发育、代谢调控和信号转导等有关的基因,为深入了解与早期分离的钩体疫苗株的全基因组水平差异提供了实验依据。
在钩体疫情严重的泰国,研究人员发现主要流行钩体菌株的血清型并无明显改变,但主要流行菌株与自然环境宿主相互作用过程中通过适应性进化,形成了新的ST 基因型(ST34),呈克隆性扩张,具有较强的传播能力,从而引起钩体病新的流行暴发[15]。对中国食品药品检定研究院与中国疾控中心传染病预防控制所收集的过去50 多年中国不同省市的120 株黄疸出血群钩体菌株进行MLST 多态性分析,发现黄疸出血群群内分化明显,共有17 种不同ST基因型[8]。作为遗传多态性分析常见两种位点标记,indel 和SNP 位点分析能进一步了解不同菌株间的基因多态性[16],本研究基于全基因组水平的比较基因组分析结果,进一步证实近期的黄疸出血群流行菌株基因组与分离于1958年的疫苗株56601 之间存在大量的SNP 和indel 位点,涉及菌株的细胞运动、信号转导机制、防御机制等重要生命活动有关的基因,提示这些差异基因及其编码产物在当前流行克隆群ST17 黄疸出血群钩体适应不同宿主环境的进化过程中发挥重要作用。因此,上述流行菌株分子基因层面的差异对致病菌蛋白表达及其致病性的影响需密切关注。巴西科学家发现,其国内绝大多数发生致病性钩体肺出血综合征(leptospiral pulmonary hemorrhage syndrome,LPHS)患者与未发生LPHS 患者感染的钩体血清型均为黄疸出血群的哥本哈根型,推测这些病原菌之间在基因和蛋白表达水平可能存在一定差异,从而导致引起LPHS的哥本哈根型钩体菌株致病性和毒力比普通哥本哈根型菌株强[17]。本研究后续将进行不同时期钩体流行株的转录和蛋白表达差异的研究,分析差异基因和蛋白的功能,评价钩体疫苗对当前钩体主要流行菌株的免疫保护性;另外,全基因组序列基因功能分析结果表明,有较多基因产物被注释为一般功能预测或未知功能,因此可继续采用反向疫苗学的技术以发现更多钩体特异性外膜或表面蛋白抗原[18-19]。同时,基于全基因组水平的系统进化发育分析发现,流行株200502与L.interrogans(56601)、L.noguchii(56189)和L.kirschneri(56611)3 个基因种进化亲缘性更近,推测可能经历相似的进化历程,与这3 个种菌株的致病性均较强相关[20]。
综上所述,本研究从全基因组水平上分析了我国人用钩体病疫苗株与当前流行菌株的差异,为今后钩体疫苗的改进提供了实验依据,同时也为未来其他相关病原体及其疫苗株比较研究提供了新思路。
