波尔多液对烟草叶际微生物群落结构与代谢功能的影响
2022-12-27刘亭亭汪汉成孙美丽尹国英向立刚蔡刘体孟建玉张长青
刘亭亭, 汪汉成, 孙美丽, 尹国英, 张 盼,向立刚, 蔡刘体, 孟建玉, 张长青
(1. 贵州省烟草科学研究院,贵阳 550081;2. 长江大学 农学院,湖北 荆州 434025)
烟草Nicotiana tabacumL. 是我国重要的经济作物,烟草赤星病 tobacco brown spot 是其成熟期的主要真菌性病害,其病原菌为子囊菌门链格孢属Alternariaspp.。该病原菌寄主范围广,烟草、棉花和柑橘等均可被其侵染[1-3]。赤星病的危害常给烟农造成重大损失。生产上常用波尔多液进行预防,该药剂是一种广谱性、不易产生抗药性、成本低的保护性杀菌剂[4],被广泛用于烟草、果树等作物病害的防控[5]。波尔多液有效成分为碱式硫酸铜,在酸性条件下转化为可溶的硫酸铜,产生少量铜离子 (Cu2+),Cu2+通过破坏病菌细胞的蛋白酶,从而使菌体代谢作用受阻,起到杀灭病菌的作用,对真菌和细菌均有抑菌活性[6]。
叶际微生物,1981 年由Blackman 首次提出,即附生或寄生于植物叶面的微生物[7],其中还含有许多有益微生物,它们通过与寄主植物的相互作用,促进植物生长[8];可以提高植物对病原菌的抗病性[9]等。当叶际微生物菌群失调后,往往会引起严重的后果,如冰核活性细菌可以诱发植物产生冻害[10],病原菌数量增加,引起植物病害[11]等。烟草叶际存在包括病原菌在内的大量微生物,它们影响着烟叶的健康与产量。叶际微生物的群落结构与代谢功能是评价叶际微生态的2 个重要指标。波尔多液作为经典药剂,在烟草上已使用多年,而大众对烟叶叶际微生物的调控规律尚缺乏了解。为此,本研究采用菌丝生长速率法和孢子萌发法测定了波尔多液对烟草赤星病菌的毒力,并采用高通量测序与Biolog 代谢表型技术分别测定了波尔多液对烟叶健康与感病组织叶际微生物群落结构和代谢功能的影响,旨在了解药剂的微生态调控规律。
1 材料与方法
1.1 供试材料
1.1.1 供试药剂 80%波尔多液可湿性粉剂 (WP)(有效成分:硫酸铜 + 氢氧化钙),购自沈阳红旗林药公司。
1.1.2 试验地点及品种 供试烟草品种为‘云烟105’。于2020 年8 月底,在贵州省毕节市威宁彝族回族苗族自治县黑石头镇,选择赤星病病叶率约为10%的烟地进行田间试验。
1.1.3 供试菌株与培养 供试菌株为烟草赤星病菌菌株CXB (保藏于贵州省烟草研究院真菌实验室)。马铃薯葡萄糖琼脂培养基 (PDA):将200 g去皮的马铃薯放于1 000 mL 沸水中蒸煮30 min,用8 层纱布过滤得到澄清滤液,向滤液中加入20 g葡萄糖和15 g 琼脂混匀,并用蒸馏水定容至1 000 mL,灭菌、备用。
1.1.4 其他供试材料 Biolog ECO 代谢板 (货号:1056),购自美国Biolog 公司 (USA,CA,Hayward);GeneJET 胶回收试剂盒 (货号:K0691) 和Ion Plus Fragment Library Kit 48 rxns 建库试剂盒 (货号:4471252),均购自Thermo Scientific 公司。DSF01A-20-100 多功能喷雾施肥器,购自贵州黔丰源农业科技开发有限公司。
1.2 波尔多液对烟草赤星病菌的毒力
采用菌丝生长速率法[12]测定波尔多液对烟草赤星病菌菌丝生长的毒力。用6 mm 打孔器打取烟草赤星病菌CXB 边缘菌丝块,挑取菌丝块分别置于波尔多液终质量浓度为1 600、800、400、200和100 mg/L 的PDA 平板上,以不加药的PDA 平板作对照,每个处理设4 个重复。于30 ℃生化培养箱中培养至对照组菌落将长满平皿时定,采用“十字交叉”法量取每个平板的菌落直径。
采用孢子萌发法[13]测定波尔多液对烟草赤星病菌孢子萌发的毒力。用无菌去离子水将在PDA培养基中培养7 d 的烟草赤星病菌CXB 产生的分生孢子洗下,双层纱布过滤得到孢子悬浮液,在显微镜下调节孢子浓度为105~106个/mL。将药剂与PDA 培养基混匀,采用倍半稀释法,配制质量浓度梯度为1 600、800、400、200 和100 mg/L 的含药PDA 平板,移取100 μL 孢子悬浮液均匀涂布在培养基表面,置于30 ℃生化培养箱中黑暗培养。以不加药剂为空白对照,每个处理设3 个重复。培养8 h 后显微镜下观察孢子萌发情况,以芽管长度大于孢子短半径视为萌发,待对照萌发率达到90%以上时记录各处理孢子萌发情况。
1.3 波尔多液对烟叶叶际微生物群落结构与代谢功能的影响
1.3.1 试验设计 设波尔多液和清水对照2 个处理,各小区随机排列,每小区60 株烟株。80%波尔多液WP 的用量为1 500 g/hm2,用水量为900 L/hm2。用背负式喷雾器正、反叶面均匀喷施,以药液能够均匀喷雾至小区所有植株叶片为准;对照组喷施清水。小区之间设保护行,避免重喷、漏喷或影响到相邻小区。
1.3.2 样品采集 分别于施药前0 d 及施药后5、10 和15 d 取样,采集波尔多液处理前后的烟叶样品用于后续试验。用消过毒的剪刀剪取下部感病部位与健康部位烟叶样品,分别装入50 mL 无菌离心管中,每个处理3 次重复。样品采集后放入低温保存箱,并迅速带回实验室开展Biolog ECO代谢功能研究,剩余样品置于 -80 ℃冰箱保存、备用,样品编号如表1 所示。
表1 样品采集信息Table 1 Samples information
1.3.3 波尔多液对烟叶叶际微生物群落结构与多样性的影响 采用CTAB (Cetyltrimethylammonium Bromide) 法[14]进行烟叶叶际微生物基因组DNA的提取,样品DNA 的纯度和浓度用琼脂糖凝胶电泳检测。将提取的DNA 置于离心管中,用无菌水稀释至浓度为1 ng/μL,以此为模板,使用引物ITS1-5F-F (5′-GGAAGTAAAAGTCGTAACAAGG-3′) 和ITS1-1F-R (5′-GCTGCGTTCTTCATCGA TGC-3′)、及515F (5′-GTGCCAGCMGCCGC GGTAA-3′) 和806R (5′-GGACTACHVGGGT WTCTAAT-3′) 分别对真菌ITS1 区域与细菌V4 区域进行扩增。参照刘畅等[15]和Chen 等[16]的方法进行多样性测序与分析,真菌和细菌分别通过UNITE(7.2) 和SILVA132 的SSUrRNA 数据库进行注释,此过程在北京诺禾致源科技股份有限公司完成。
1.3.4 波尔多液对烟叶叶际微生物代谢功能的影响 分别取不同时期不同部位烟叶混合样品各1 g,置于盛有50 mL 0.8%的无菌生理盐水的100 mL三角瓶中,于28 ℃、180 r/min 振荡摇培2 h,静置30 min 后取100 μL 上清液,分别加入到ECO代谢板的测试孔中[17]。将接菌后的ECO 代谢板置于OmniLog 恒温培养箱中,于28 ℃条件下培养7 d,采用Biolog D5E_OKA_data.exe 软件收集烟叶叶际微生物在生长过程中代谢孔内颜色变化值,使用HemI 软件制作热图[18],分析叶际微生物的代谢功能。
1.4 数据处理及统计分析
使用Excel 2019、DPS7.5 软件进行数据处理及统计分析。
2 结果与分析
2.1 波尔多液对烟草赤星病菌的毒力
如表2 所示,波尔多液对烟草赤星病菌菌丝生长和孢子的萌发均有抑制作用,且随着药剂浓度的增加,其抑制作用逐渐增强,其中对菌丝生长的抑制活性较强。波尔多液抑制菌丝生长的EC50值和EC95值分别为450.19 和4 987.45 mg/L;抑制孢子萌发的EC50值和EC95值分别为757.17 和3 108.01 mg/L。在药剂质量浓度为1 600 mg/L时,对菌丝生长和分生孢子萌发的抑制率分别为79.56%和82.41%。

表2 波尔多液对烟草赤星病菌的毒力Table 2 Toxicity test of Bordeaux mixture to Alternaria alternata
2.2 波尔多液对烟叶叶际微生物群落结构与多样性的影响
2.2.1 波尔多液对烟叶叶际微生物群落结构的影响2.2.1.1 对细菌群落结构的影响 在细菌群落结构门水平上,施药前健康 (BEJ0) 与感病 (BEB0) 烟叶叶际优势菌门均为变形菌门 (Proteobacteria,6.93%和39.07%) 和厚壁菌门 (Firmicutes,16.45%和0.65%)。波尔多液处理5 d 时,健康 (BEJ1) 和感病 (BEB1) 烟叶叶际变形菌门的相对丰度均有所下降,健康 (BEJ1) 烟叶叶际厚壁菌门的相对丰度下降较多;处理10 d 时,感病 (BEB2) 烟叶叶际厚壁菌门相对丰度显著增加,而健康 (BEJ2) 烟叶叶际变形菌门相对丰度有所增加;处理15 d 时,健康 (BEJ3) 和感病 (BEB3) 烟叶叶际变形菌门相对丰度均有增加 (图1)。
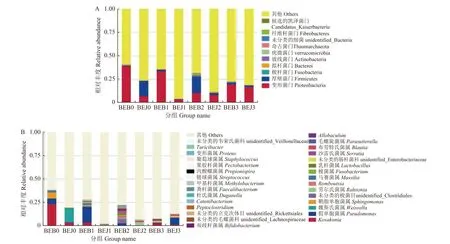
图1 波尔多液对烟叶叶际细菌门 (A) 和属(B)水平上的群落组成的影响Fig.1 The effects of Bordeaux mixture on the community composition of bacteria in the phyllosphere of tobacco at phyla(A) and genera(B) levels
在细菌群落结构属水平上,施药前健康 (BEJ0)与感病 (BEB0) 烟叶叶际优势菌属为Kosakonia(3.46%和22.38%) 和假单胞菌属 (Pseudomonas,0.22%和5.95%),与感病 (BEB0) 烟叶相比,健康(BEJ0) 烟叶的魏斯氏菌属 (Weissella,15.26%) 显著高于感病 (BEB0) 烟叶 (0.00%)。波尔多液处理5 d 时,健康 (BEJ1) 和感病 (BEB1) 烟叶叶际Kosakonia、鞘脂单胞菌属 (Sphingomonas) 和乳杆菌属 (Lactobacillus) 相对丰度降低,而假单胞菌属(Pseudomonas)、劳尔氏菌属 (Ralstonia)、马赛菌属 (Massilia)、未分类的肠杆菌科 (unidentified_Enterobacteriaceae)、未分类的立克次体目 (unidentified_Rickettsiales)、沙雷氏菌属 (Serratia) 相对丰度明显上升。处理10 d 时,健康 (BEJ2) 和感病 (BEB2)烟叶叶际Kosakonia和鞘脂单胞菌属相对丰度持续降低或不变,假单胞菌属和马赛菌属 (Massilia) 相对丰度开始降低,而劳尔氏菌属和沙雷氏菌属相对丰度持续增加。处理15 d 时,健康 (BEJ3) 和感病 (BEB3) 烟叶叶际Kosakonia相对丰度开始上升,沙雷氏菌属相对丰度持续增加 (图1)。
2.2.1.2 对真菌群落结构的影响 在真菌群落结构门水平上,施药前健康 (BEJ0) 与感病 (BEB0) 烟叶叶际优势菌门均为子囊菌门 (Ascomycota,63.82%和93.74%) 和担子菌门 (Basidiomycota,6.82%和2.53%)。波尔多液处理5 d 时,感病 (BEB1)烟叶叶际子囊菌门相对丰度小幅度增长,担子菌门相对丰度小幅度下降,而健康 (BEJ1) 烟叶中子囊菌门相对丰度大幅度减少,担子菌门相对丰度大幅度增长。处理10 d 时,感病 (BEB2) 烟叶中子囊菌门相对丰度出现较大幅度降低,担子菌门相对丰度出现较大幅度增长,而健康 (BEJ2) 烟叶中则与之相反。处理15 d 时,感病 (BEB3) 烟叶中子囊菌门和担子菌门相对丰度同时降低,而健康 (BEJ3) 烟叶中子囊菌门相对丰度降低,担子菌门相对丰度增长 (图2)。
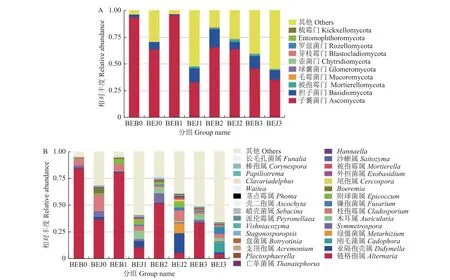
图2 波尔多液处理对烟叶叶际真菌门 (A) 和属(B)水平上的群落组成的影响Fig.2 The effects of Bordeaux mixture on the community composition of fungi in the phyllosphere of tobacco at phyla(A) and genera(B) levels
在真菌群落结构属水平上,施药前健康 (BEJ0)与感病 (BEB0) 烟叶叶际真菌属水平主要菌群为链格孢属 (Alternaria,36.48% 和84.52%)、Symmetrospora(5.56% 和2.27%) 和枝孢霉属(Cladosporium,14.87%和6.66%),与健康 (BEJ0)烟叶相比,感病 (BEB0) 样品的链格孢属 (84.52%)显著高于健康样品 (36.48%) (P<0.05)。波尔多液处理5 d 时,健康 (BEJ1) 与感病 (BEB1) 烟叶叶际链格孢属、Symmetrospora、枝孢霉属、Hannaella、亡革菌属 (Thanatephorus)、Vishniacozyma6 个菌属相对丰度下降,而亚隔孢壳属 (Didymella)、绿僵菌属 (Metarhizium)、木耳属 (Auricularia)、镰孢菌属 (Fusarium)、附球菌属 (Epicoccum)、尾孢属(Cercospora)、被孢霉属 (Mortierella)、沙蜥属(Saitozyma)、Plectosphaerella、Papiliotrema10 个菌属菌群相对丰度均上升。处理10 d 时,健康(BEJ2) 与感病 (BEB2) 烟叶叶际链格孢属、亡革菌属相对丰度持续降低,绿僵菌属、镰孢属、被孢霉属、Plectosphaerella和Papiliotrema相对丰度持续上升,Vishniacozyma相对丰度开始上升。处理15 d 时,健康 (BEJ3) 与感病 (BEB3) 烟叶叶际链格孢属相对丰度持续降低,Papiliotrema相对丰度开始降低,其余则呈不规律增减 (图2)。
花瓣图分析结果表明 (图3),在可操作分类单元 (OTU) 水平下,各样本细菌、真菌群落分别共有OTU 种类数为6 和39 种,共有的细菌属为Kosakonia、假单胞菌属及未分类的立克次体目等,共有的真菌属为链格孢属、亚隔孢壳属、Symmetrospora及枝孢霉属等,施药前健康与感病烟叶细菌群落独有的OTU 种类数分别为12 和6 种,真菌群落独有的OTU 种类数分别为3 和50 种。波尔多液处理5 和10 d 时,健康与感病烟叶细菌、真菌群落独有的OTU 种类数均有所上升。处理15 d 时,其独有的OTU 种类数开始降低,而感病烟叶的真菌群落种类数持续上升。
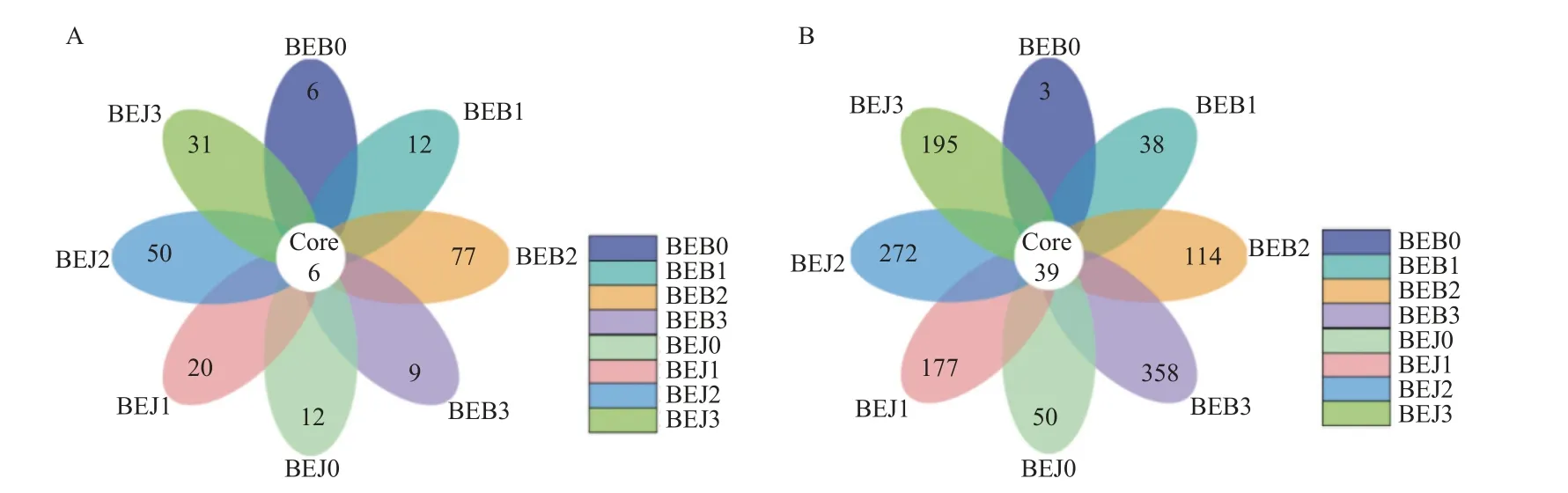
图3 波尔多液处理烟叶叶际细菌 (A) 和真菌(B)群落OTU 分布花瓣图Fig.3 Flower map of OTU distribution of bacterial(A) and fungal(B) communities in phyllosphere of tobacco treated with Bordeaux mixture
2.2.2 波尔多液对烟叶叶际微生物群落多样性的影响 由表3 可知:健康与感赤星病烟叶叶际真菌和细菌测序覆盖度指数均达到0.90 以上,表明所有样品测序数据合理,可以真实、合理地反映微生物群落的多样性。
细菌多样性结果 (表3) 表明:处理前,健康烟叶 (BEJ0) 叶际细菌多样性指数 (Shannon) 和丰富度指数 (Chao1、ACE) 均高于感病烟叶 (BEB0),但无显著性差异。处理5 d 时,感病烟叶 (BEB1) 叶际细菌多样性指数显著高于健康烟叶 (BEJ1)。处理10 d 时,感病烟叶 (BEB2) 与健康烟叶 (BEJ2)叶际细菌丰富度指数达到最高值。处理15 d 时,感病烟叶样品 (BEB3) 与健康烟叶 (BEJ3) 叶际细菌丰富度指数降低,但无显著性差异。
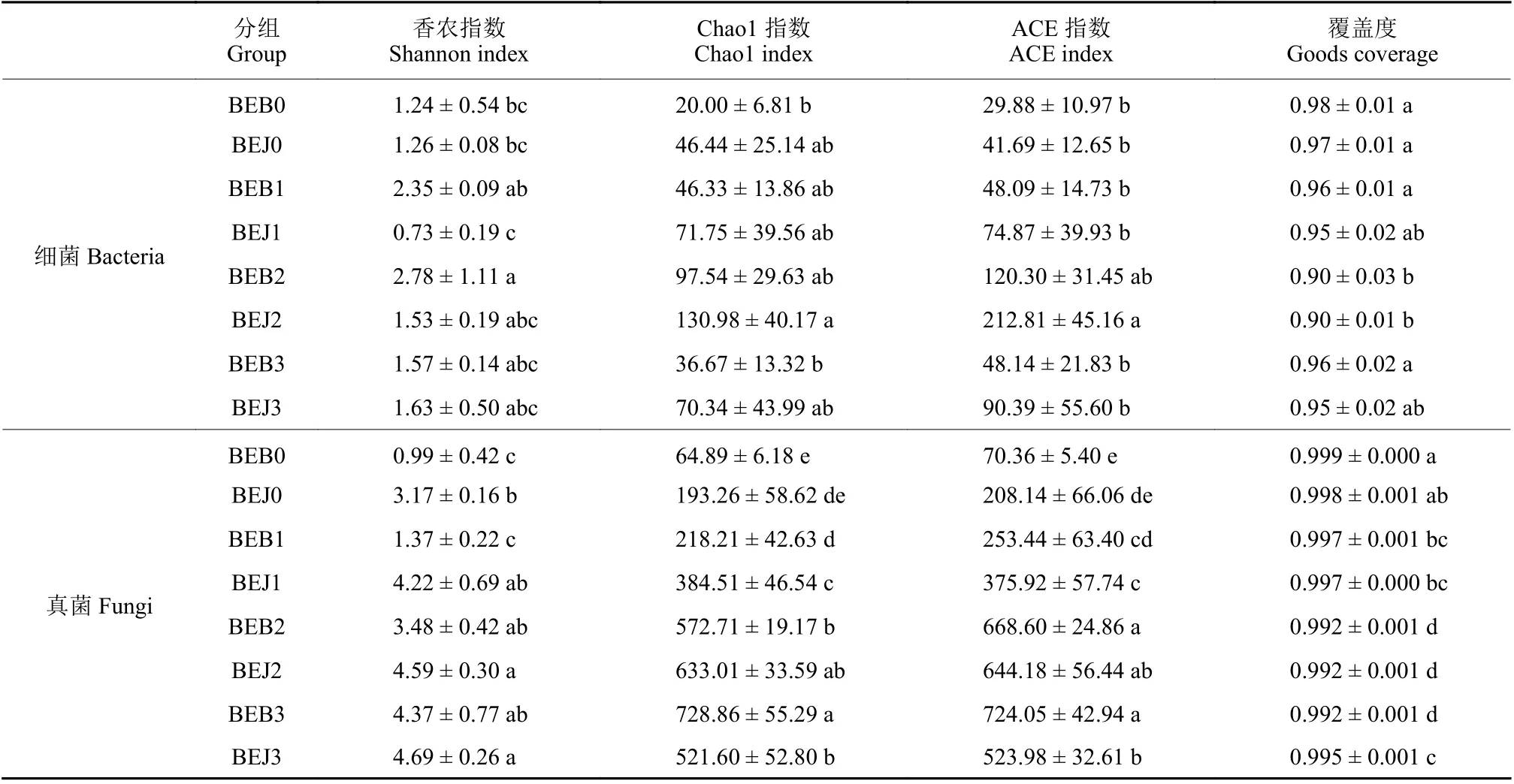
表3 波尔多液对烟叶叶际微生物多样性的影响 (OTU 水平)Table 3 Effect of Bordeaux mixture on the diversity of tobacco phyllosphere microorganism (OTU level)
真菌多样性结果 (表3) 表明,施药前,健康烟叶 (BEJ0) 叶际真菌多样性指数和丰富度指数均高于感病烟叶 (BEB0),但丰富度之间无显著性差异。波尔多液处理5 和10 d 时,感病烟叶 (BEB1、BEB2) 叶际真菌与健康烟叶 (BEJ1、BEJ2) 多样性与丰富度指数增长。处理15 d 时,感病烟叶 (BEB3)叶际真菌丰富度指数显著高于健康烟叶 (BEJ3)。
2.3 波尔多液对烟叶叶际微生物代谢功能的影响
Biolog ECO 微孔板中含有糖类、氨基酸、羧酸类、双亲化合物、聚合物和胺/氨基化合物共31 种碳源。波尔多液对烟叶叶际微生物代谢功能的影响结果如图4 所示,施药前,感病 (BEB0) 与健康 (BEJ0) 烟叶微生物均可高效代谢 (颜色值变化值>200) 除氨基酸类的L-苏氨酸 (L-threonine) 和羧酸类的α-丁酮酸 (α-ketobutyric acid) 外的29 种碳源,感病 (BEB0) 烟叶无特有高效代谢的碳源。
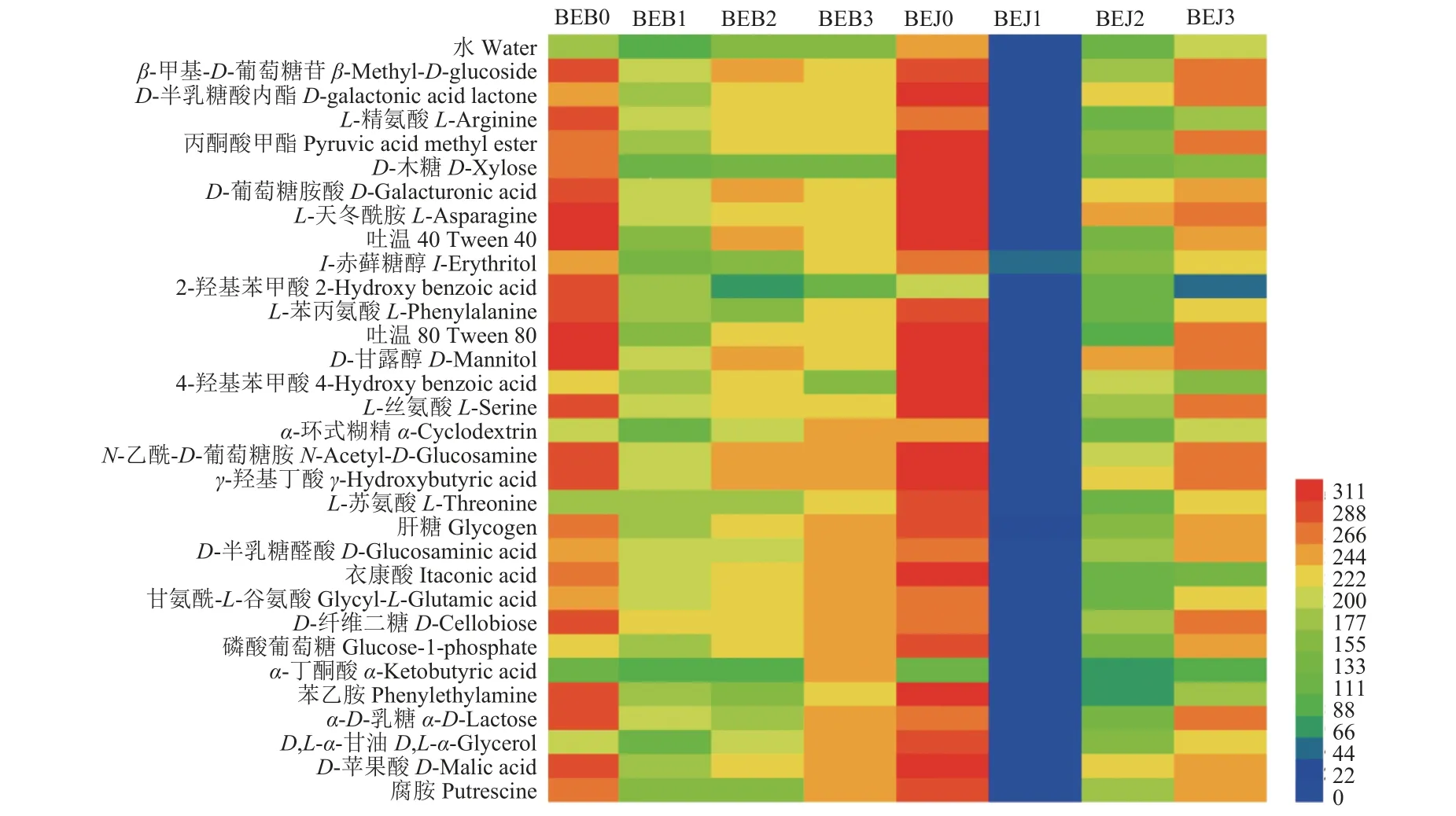
图4 波尔多液处理烟叶叶际微生物代谢功能的聚类热图Fig.4 Cluster heat map of the effect of Bordeaux mixture on the metabolism of tobacco phyllosphere microorganism
喷施波尔多液5、10 和15 d 后,健康与感病烟叶叶际微生物代谢功能均被不同程度抑制。健康烟叶叶际微生物的代谢功能表明,波尔多液处理5 d 时,叶际微生物对31 种碳源的代谢均被抑制,无高效代谢碳源;处理10 d 时,叶际微生物的代谢功能逐渐恢复,但仍有23 种碳源的代谢被抑制,包括β-甲基-D-葡萄糖苷、L-精氨酸、丙酮酸甲酯等,代谢高的碳源有8 种碳源,包括D-半乳糖酸内酯、D-葡萄糖胺酸、L-天冬酰胺等;处理15 d 时,叶际微生物的代谢功能进一步加强,6 种碳源的代谢仍被抑制,包括2-羟基苯甲酸、L-精氨酸、D-木糖等,24 种碳源能被高效代谢,包括β-甲基-D-葡萄糖苷、丙酮酸甲酯和吐温40 等。
相比而言,感病烟叶叶际微生物的代谢功能表明,波尔多液处理5 d 时,叶际微生物对17 种碳源的代谢被抑制,包括D-半乳糖酸内酯、丙酮酸甲酯、D-木糖等,但可高效代谢13 种碳源,包括β-甲基-D-葡萄糖苷、L-精氨酸、D-葡萄糖胺酸等;处理10 d 时,叶际微生物对7 种碳源的代谢被抑制,包括D-木糖、I-赤藓糖醇、2-羟基苯甲酸等,可高效代谢22 种碳源,包括β-甲基-D-葡萄糖苷、D-半乳糖酸内酯、丙酮酸甲酯等;处理15 d 时,叶际微生物对3 种碳源的代谢被抑制,为D-木糖、2-羟基苯甲酸和4-羟基苯甲酸,但可高效代谢28 种碳源,包括β-甲基-D-葡萄糖苷、D-半乳糖酸内酯、L-精氨酸等。
3 结论与讨论
本研究中,波尔多液对烟草赤星病菌菌丝生长和孢子萌发的EC50值均大于450 mg/L,活性较低。这可能与波尔多液的作用机理有关,它主要通过释放的铜离子来杀菌,铜离子的作用靶标虽然较多,但活性较弱。为此,该药剂适合做保护性药剂,喷施后在叶片形成一层保护膜,起到抑制病菌萌发的作用。
碳源是微生物生命活动的基础,Biolog ECO代谢板含有大部分微生物生存所需的31 种常见碳源,可以反映烟叶叶际微生物的营养需求特性。本研究测定了感赤星病烟叶叶际微生物对糖类、氨基酸、羧酸类、双亲化合物和聚合物以及胺/氨基化合物碳源的代谢利用情况,发现羧酸类的代谢利用程度低于其他5 类碳源。该结果与张文艳[19]及周运来等[20]的研究结果类似。推测烟草叶际微生物不喜偏酸性的碳源,今后可以尝试采用羧酸类碳源的营养调控来防控烟草赤星病,相关假设有待下一步验证。喷施波尔多液后,健康与感病烟叶叶际微生物的代谢功能与施药前相比有所变化,可抑制对β-甲基-D-葡萄糖苷、D-半乳糖酸内酯等绝大多数碳源代谢,抑制作用可达5 d,之后微生物对碳源的代谢程度均有不同程度的上升,且对健康叶际微生物的影响大于感病组织,表明波尔多液适合用作保护性药剂,且持效期相对较短。
叶际微生物的多样性与宿主植物的健康生长息息相关,施药前健康烟叶的真菌、细菌群落的多样性指数和丰富度指数均高于感病烟叶,与Chen 等[21]研究发现拟南芥发病叶片内微生物多样性降低的结果一致,表明植株可通过调节叶际微生物菌群平衡来保证自身健康。施药5 d 后,感病和健康烟叶的真菌、细菌的多样性指数 (Shannon)和丰富度指数 (Chao1、ACE) 大多数高于施药前,与韩永琴[22]发现施用化学药剂后根际土壤菌群丰富度及多样性增加的结果一致,表明药剂的使用对微生物种群存在影响,其原因可能是施用药剂后刺激了某些菌群 (如:细菌的假单胞菌属、劳尔氏菌属和马赛菌属等;真菌的亚隔孢壳属、绿僵菌属和木耳属等) 的大量繁殖。而某些植物致病菌的大量繁殖,则有可能会引起病原菌产生复合侵染,导致烟株发病更严重。
高通量测序结果发现,感病与健康烟叶优势细菌属为Kosakonia和假单胞菌属,这与刘畅等[15]的研究结果类似,本研究发现烟叶叶际同时也存在劳尔氏菌属[23]、鞘脂单胞菌属[24]等菌属细菌。感病与健康烟叶优势真菌属为链格孢属、Symmetrospora和枝孢霉属,与刘畅等[25]、向立刚等[26]发现感赤星病鲜烟叶和烤后烟叶优势真菌属的研究结果类似。同时,本研究还发现亚隔孢壳属[27]、镰孢属[28]等致病真菌的存在。施用药剂后,植物叶际或根际土壤微生物的群落结构会发生改变。本研究发现,喷施波尔多液后,烟草叶际真菌群落中链格孢属和枝孢霉属等相对丰度降低,该结果与前人研究结果[29-31]一致。在本研究中,波尔多液不仅作用于链格孢属等植物致病菌,对鞘脂单胞菌属、Kosakonia等有益菌群也产生了抑制作用[32],Kosakonia是一种固氮菌,可以帮助植物有效进行氮利用吸收[33],而这类菌群的改变可能会直接影响植物的各种生理代谢活动,进而影响植物的正常生长。目前,有关杀菌剂对烟叶叶际微生态影响的研究报道较少,本研究结果可为杀菌剂防控植物病害的微生态机制提供参考。由于烟草赤星病为叶部真菌性病害,喷施药剂时,主要喷施在叶片上,散落在地面的不多,因此本次试验重点研究其对叶际微生物的影响,下一步将继续开展波尔多液对烟草根际土壤微生物群落结构与代谢功能影响的研究。
