降尘对阿克苏地区主栽果树叶片光合特性及叶绿素荧光特性的影响
2022-12-24孙守霞朴涵琪周光辉王贺松张述斌郝金莲
孙守霞,陈 虹,吕 威,朴涵琪,周光辉,王贺松,张述斌,郝金莲
(新疆农业大学 林学与风景园林学院/新疆教育厅干旱区林业生态与产业技术重点试验室,新疆 乌鲁木齐830052)
浮尘天气是全球干旱、半干旱地区极其普遍的一种风沙灾害性天气现象并对植物有很大的危害[1-2]。新疆地处欧亚大陆腹地塔里木盆地边缘,气候干燥、多风少雨,复杂的地理环境导致沙尘天气频发[3]。新疆南疆特色林果资源丰富,林果种植已成为当地支柱产业及农民增收的主要来源之一,由于南疆盆地沙尘天气高发时段集中在3—9月[4],伴随果树的整个生长期并对果树生长造成不良影响。前人研究表明,沙尘覆盖在叶片表面会阻碍叶片对光能的吸收,堵塞植物叶片表面的气孔,使蛋白质变性[5],抗氧化酶含量增加[6],造成叶片角质层损坏,影响植物营养元素的吸收和代谢[7]、开花授粉、坐果等过程[8],最终影响果树果实的产量和品质。目前关于降尘的研究主要集中在短时间内对果树叶片的危害[9-12],为了更好地了解降尘对果树生长发育的危害,必须深入了解长时间降尘对果树叶片生理生态机理的影响。本实验以新疆阿克苏地区的苹果(Maluspumila)、核桃(Juglansregia)、枣(Ziziphusjujube)3种主栽果树为材料,进行叶片滞尘量及叶片响应自然降尘的光合特性及叶绿素荧光特性的研究,以期探索果树对降尘的生理代谢响应机制,并为阿克苏地区主栽果树的栽培管理提供一定的科学理论依据。
1 材料与方法
1.1 材料
实验园位于新疆阿克苏地区温宿县新疆农业大学林果试验基地(40°52′~42°15′N,79°28′~81°30′E)。实验样地2 hm2,实验材料为新红1号苹果、温185核桃、灰枣3个果树品种。新红1号苹果树高4 m,温185核桃树高4.5 m,灰枣树高3 m,树龄均为10 a,行间距为4 m×6 m,南北行向栽植。自然状态下接受降尘影响的样树5株为降尘组,以每7 d用清水冲洗一次叶片的样树5株为对照组,于2020年6—8月进行滞尘量、光合参数、荧光参数和叶绿素含量测定。
1.2 测定项目与方法
1.2.1 滞尘量的测定
参考路艳等[13]的方法,6—8月每个月每7 d(即7、14、21、28 d)采集一次叶片,在树体四周同一高度处均匀采样,根据叶片大小,采20~30片,将采集的叶片放入信封袋中带回实验室。首先用万分之一的电子天平称质量(W1),W1即为叶片及叶片上的降尘颗粒物的质量,然后将叶片表面灰尘用流水冲洗干净,待阴干后再称重(W2),2次质量之差即为叶片滞尘量。用万深叶面积测量仪LA-S测定叶表面积(S),则单位叶面积滞尘量Q=(W1-W2)/S,结果取均值作为单月平均滞尘量。
1.2.2 光合参数和荧光参数的测定
分别于6—8月的每月下旬,选择晴朗静风天气,选取对照组与降尘组树体中上部外围光照充足、生长健康的叶片进行测定。采用美国产LI-6800便携式光合仪测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等光合参数;采用英国 Hansatech便携式调制荧光仪(FMS-2)测定荧光参数,用叶夹将要测定的叶片夹住暗适应15 min,在低强度的测量红光照射下,得到最小荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)。然后打开暗适应夹,叶片在自然光条件下适应30 min后,在内源 300 μmol·m-2·s-1光化光照射下测得稳态荧光(Fs),在1 300 μmol·m-2·s-1的饱和脉冲白光下照射30 s后获得光下最大荧光(Fm),最大光化学效率(Fv/Fm)可从FMS-2脉冲调制式荧光分析仪上直接读取。每处理5个重复,结果取平均值。计算非光化学淬灭系数(NPQ)、光化学淬灭系数(qP)。
1.2.3 叶绿素含量的测定
参考董斌等[14]的方法,采用80%丙酮法测定叶绿素含量。将测量完光合及荧光参数的苹果、核桃、枣叶片采集后带回实验室,避开叶脉分别称取0.02、0.04、0.03 g,用5 mL 80%丙酮研成匀浆,放置在试管中暗处理20 min后进行过滤,然后用80%丙酮定容至10 mL,摇匀。以80%丙酮为空白对照,在波长663、646 nm下测定吸光度,计算叶绿素总量。每处理5个重复,结果取平均值。
1.3 数据处理
数据以“平均数±标准误”表示。采用Microsoft Excel 2010软件进行数据整理和绘图,采用SPSS 20.0 软件用LSD法对所测数据进行显著性检验和多重比较。采用Pearson相关性分析法分析3种果树滞尘能力与叶片光合特征及叶绿素荧光参数之间的关系,相关系数矩阵图采用Origin软件制作。
2 结果与分析
2.1 三种果树单位叶面积滞尘量
3种果树单位叶面积滞尘量见表1,6—8月3种果树单位叶面积滞尘量呈一定的累积效应。随着时间的推移,3种果树单位叶面积滞尘量之间存在显著(P<0.05)差异。苹果、核桃、枣单位叶面积平均滞尘量分别为 8.78、7.42、8.01 mg·cm-2,滞尘能力为苹果>枣>核桃。

表1 三种果树单位叶面积滞尘量
2.2 降尘对3种果树叶片叶绿素含量的影响
降尘对3种果树叶片叶绿素含量(Chl)的影响见图1。受降尘影响,3种果树叶片叶绿素含量在不同时间段均显著(P<0.05)低于对照植株。叶片叶绿素含量为苹果>核桃>枣,3种果树叶片叶绿素含量存在显著(P<0.05)差异。6—8月苹果降尘处理的植株叶片叶绿素含量分别比对照植株显著(P<0.05)降低14.30%、15.55%、17.56%;核桃降尘处理的植株叶片叶绿素含量分别比对照植株显著(P<0.05)降低9.54%、22.47%、29.70%;枣降尘处理的植株叶片叶绿素含量分别比对照植株显著(P<0.05)降低10.16%、13.40%、17.19%。由此可以看出,果树叶片受降尘影响时间越长,叶片叶绿素含量下降的程度越大。

不同大写字母表示同一处理的不同树种间差异显著(P<0.05);不同小写字母表示同一时间段两种处理间差异显著(P<0.05);图中ck(A)为苹果对照处理、da(A)为苹果降尘处理、ck(W)为核桃对照处理、da(W)为核桃降尘处理、ck(J)为枣对照处理、da(J)为枣降尘处理。下同。
2.3 降尘对3种果树叶片光合特性的影响
2.3.1 降尘对3种果树叶片净光合速率及胞间CO2浓度的影响
如图2-A所示,受降尘影响,3种果树叶片Pn均显著(P<0.05)低于对照植株,并且随着处理时间的推移,3种果树之间Pn在7月出现显著(P<0.05)差异。6—8月苹果降尘处理的植株叶片Pn分别比对照植株显著(P<0.05)降低13.83%、26.11%、25.08%;核桃降尘处理的植株叶片Pn分别比对照植株显著(P<0.05)降低8.70%、19.77%、26.71%;枣降尘处理的植株叶片Pn分别比对照植株显著(P<0.05)降低25.30%、31.16%、27.42%。
降尘对3种果树叶片胞间CO2浓度的影响见图2-B。随着处理时间的推移,3种果树Ci在8月出现显著性差异,且降尘处理的植株Ci均显著(P<0.05)高于对照植株。6—8月苹果降尘处理的植株叶片Ci分别比对照植株显著(P<0.05)升高9.87%、10.15%、11.01%;核桃降尘处理的植株叶片Ci分别比对照植株显著(P<0.05)升高9.11%、5.29%、3.40%;枣降尘处理的植株叶片Ci分别比对照植株显著(P<0.05)升高9.07%、7.67%、1.45%。

图2 降尘对3种果树叶片净光合速率及胞间CO2浓度的影响
2.3.2 降尘对3种果树叶片气孔导度及蒸腾速率的影响
如图 3-A所示,降尘处理的苹果和核桃叶片Gs呈先升后降的变化趋势,而枣则相反。6月未受降尘影响的3种果树Gs无显著差异,受降尘影响后出现显著(P<0.05)差异。6—8月苹果降尘处理的植株叶片Gs分别比对照植株显著(P<0.05)降低13.13%、10.93%、29.11%;核桃降尘处理的植株叶片分别比对照植株显著(P<0.05)降低5.67%、9.69%、21.80%;枣降尘处理的植株叶片分别比对照植株显著(P<0.05)降低9.16%、28.93%、31.69%。
如图3-B所示,3种果树叶片Tr变化规律与Gs一致。7月两种处理下3种果树叶片Tr之间均出现显著(P<0.05)差异,Tr值为核桃>苹果>枣。6—8月苹果降尘处理的植株叶片Tr分别比对照植株显著(P<0.05)降低16.17%、12.80%、8.72%;核桃降尘处理的植株叶片Tr分别比对照植株显著(P<0.05)降低19.53%、13.09%、10.83%;枣降尘处理的植株叶片Tr分别比对照植株显著(P<0.05)降低1.06%、20.89%、22.55%。随着处理时间的推移,枣Tr受降尘的影响增大,而苹果和核桃Tr受降尘影响程度越来越小,但两种处理之间仍存在显著(P<0.05)差异。

图3 降尘对3种果树叶片气孔导度及蒸腾速率的影响
2.4 降尘对叶片叶绿素荧光特性的影响
2.4.1 降尘对3种果树叶片Fv/Fm及Fo的影响
如图4-A所示,未接受降尘影响的3种果树叶片Fv/Fm基本不变,而受降尘影响后3种果树叶片Fv/Fm均显著(P<0.05)低于对照植株,3种果树Fv/Fm之间无显著差异。随着处理时间的推移,苹果和核桃Fv/Fm呈下降趋势,而枣呈先降后升的变化趋势。6—8月苹果降尘处理的植株Fv/Fm分别比对照植株显著(P<0.05)降低1.54%、1.91%、2.23%;核桃降尘处理的植株Fv/Fm分别比对照植株显著(P<0.05)降低1.54%、2.42%、3.52%;枣降尘处理的植株Fv/Fm分别比对照植株显著(P<0.05)降低1.89%、3.99%、2.98%。
如图4-B所示,受降尘影响后,核桃Fo逐渐上升,苹果和枣Fo呈先升后降的变化趋势。3种果树处理植株Fo均显著(P<0.05)高于对照植株。6—8月苹果降尘处理的植株Fo分别比对照植株显著(P<0.05)升高11.53%、15.58%、8.24%;核桃降尘处理的植株Fo分别比对照植株显著(P<0.05)升高6.01%、11.37%、13.51%;枣降尘处理的植株Fo分别比对照植株显著(P<0.05)升高15.53%、26.59%、15.64%。
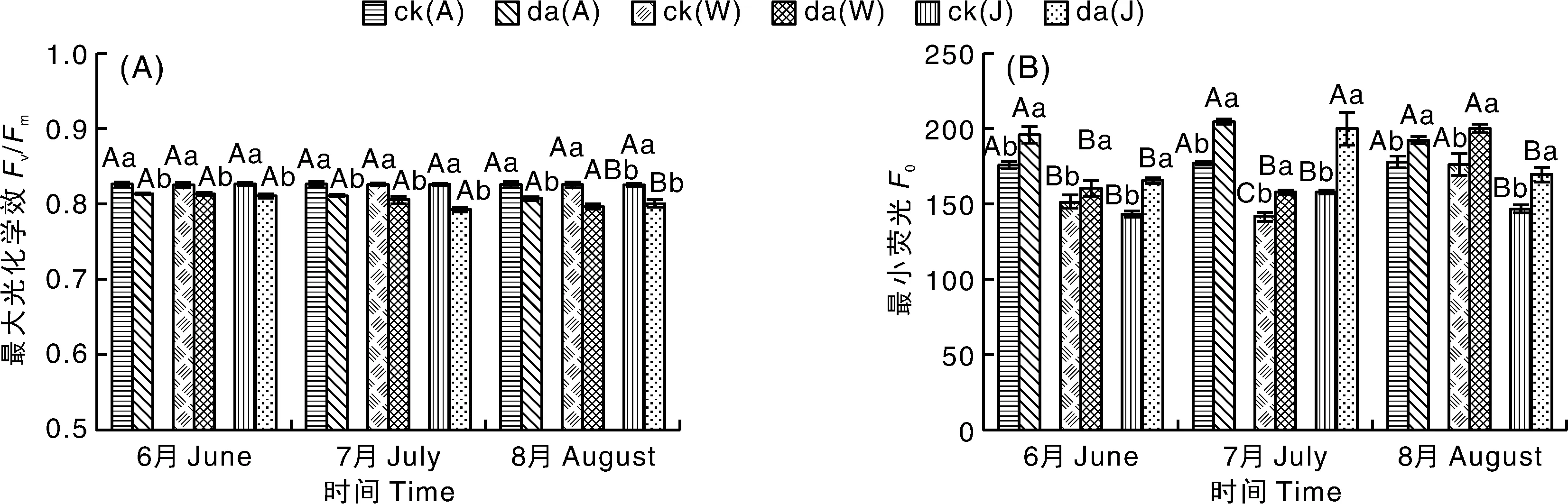
图4 降尘对三种果树叶片Fv/Fm及Fo的影响
2.4.2 降尘对3种果树叶片qP及NPQ的影响
如图5-A所示,受降尘影响的苹果和核桃NPQ呈先降后升的变化趋势,枣NPQ则呈下降趋势。随着处理时间的推移,降尘处理的植株叶片NPQ均显著(P<0.05)高于对照植株。6—8月苹果降尘处理的植株NPQ分别比对照植株显著(P<0.05)升高42.33%、32.75%、26.35%;核桃降尘处理的植株NPQ分别比对照植株显著(P<0.05)升高27.20%、52.95%、27.79%;枣降尘处理的植株NPQ分别比对照植株显著(P<0.05)升高23.30%、3.19%、17.65%。
如图5-B所示,两种处理下3种果树qP存在显著(P<0.05)差异。受降尘影响后3种果树降尘处理的植株叶片qP显著(P<0.05)低于对照植株。6—8月苹果降尘处理的植株qP分别比对照植株显著(P<0.05)降低4.48%、5.26%、5.08%;核桃降尘处理的植株qP分别比对照植株显著(P<0.05)降低3.32%、6.90%、13.74%;枣降尘处理的植株qP分别比对照植株显著(P<0.05)降低19.31%、7.76%、24.11%。
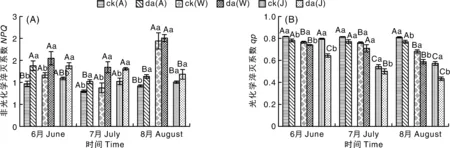
图5 降尘对三种果树叶片qP及NPQ的影响
2.5 三种果树滞尘量与光合参数及叶绿素荧光参数的相关性分析
如图6所示,通过对6—8月3种果树单位叶面积滞尘量与光合参数及叶绿素荧光参数的相关性分析显示,3种果树叶片滞尘量与Pn、Chl及Fv/Fm之间显著(P<0.05)负相关或极显著(P<0.01)负相关,与Ci显著(P<0.05)正相关。此外苹果滞尘量还与Gs之间极显著正相关,枣滞尘量与Tr极显著(P<0.01)负相关。核桃滞尘量与qP极显著(P<0.01)负相关,与NPQ显著(P<0.05)正相关。

*表示在0.05水平上显著相关;**表示在0.01水平上显著相关。
3 讨论
大量研究表明,果树滞尘能力与叶片表面粗糙程度[15]、有无绒毛[16]以及叶柄长短等[17]多种因素有关。本研究发现,3种果树滞尘能力为苹果>枣>核桃。滞尘能力最大的苹果因其叶片表面粗糙且具有沟状组织使其能够接受较多的降尘。而核桃叶片表面较为光滑,两面无毛,叶柄长,其滞尘能力最弱。表明果树滞尘能力与叶面形态结构密切相关,叶面粗糙程度是决定果树叶片滞尘能力强弱的重要因素,这在杏(Armeniacavulgaris)、樱桃(Prunuspseudocerasus)、石榴(Punicagranatum)等[18-19]多种果树的相关研究中,也得到了相似结论。
植物叶片叶绿素含量是光合能力及生理活性的重要指标,环境胁迫会引起植物体内叶绿素酶活性升高,加快叶绿素的降解[20-22]。李海波等[23]研究表明,环境胁迫会加速水稻(Oryzasativa)叶绿素的降解,类囊体膜的稳定性降低,导致光合速率下降。本研究发现,降尘导致3种果树叶片叶绿素含量显著降低,并且叶面滞尘时间越长,叶绿素含量下降程度越大。这在欧洲李(Prunusdomestica)[24],阿月浑子(Pistaciavera)[25]的相关研究中,也得到了进一步证实,表明降尘引发植物体内产生大量活性氧,叶绿体结构被破坏,从而抑制植物叶片叶绿素的形成。
光合作用是植物生长发育的基础,外界环境改变后,植物的净光合速率Pn随之改变。Farquhar等[26]指出,因Ci值升高造成的植物Pn下降,主要与叶片叶肉细胞光合能力的下降有关;而Ci值降低造成的植物Pn下降,则由气孔受限引发。本研究发现,受降尘影响的3种果树叶片Pn显著降低,而Ci显著升高,非气孔因素是导致其Pn下降的主要原因,这在西梅(Prunusdomestica)[27]、榅桲(Cydoniaoblonga)[28]和苹果[29]等研究中也得出一致结论,表明降尘导致植物叶片光合机构受损,CO2的固定和溶解能力下降[30]。但李巧云等[31]则认为浮尘使冬小麦(Triticumaestivum)Pn下降系由气孔受限引发,降尘阻碍了CO2进入叶片的能力,胞间CO2浓度随之降低,进而影响到净光合速率,使光合速率下降。表明降尘会通过气孔或者非气孔的因素造成植物叶片净光合速率下降,但因植物类型、种类的不同存在差异。秦璐等[32]、杨茂生等[33]通过对梭梭(Haloxylonammodendron)、侧柏(Platycladusorientalis)的研究发现,植物叶片降尘量越大,气孔导度和蒸腾速率越小,表明叶面降尘引发气孔堵塞,水汽扩散受阻,从而导致蒸腾速率降低。本研究发现,降尘导致果树叶片气孔导度和蒸腾速率均降低。这与莫治新等[34]研究结果一致,说明叶面长期滞尘导致气孔堵塞严重,进而加剧蒸腾速率降低。
植物叶绿素荧光能够直观地反映出环境胁迫对植物叶片光合作用的伤害机理[35]。Fv/Fm是判断植物是否受到光抑制的重要标准[36],Fo表示黑暗条件下PSⅡ反应中心全部开放的荧光水平,PSⅡ反应中心受到破坏Fo就会增加[37]。本研究发现,降尘使3种果树叶片Fv/Fm显著降低,Fo显著升高,这在杏[38]的相关研究中,也得到了进一步证实,说明降尘使果树叶片PSⅡ反应中心产生一定程度的伤害或可逆性失活,果树叶片原初光能转换效率降低,出现光抑制现象。王兆等[39]认为低温胁迫使彩叶草(Solenostemonscutellarioides)qP降低,NPQ上升,表明环境胁迫使植物叶片PSⅡ反应中心的开放程度降低,参与CO2固定的电子减少。本研究发现,降尘导致3种果树qP显著降低,NPQ显著上升。这与付春霞等[40]对苹果的研究结果一致,说明降尘胁迫使植物用于光化学反应的能量减少,植物通过加强非化学反应的能量耗散来避免过量的光伤害。
吕东蓬[41]研究发现,植物光合特征参数损失率与叶片滞尘量之间呈显著的正相关关系,即滞尘量越大,植物光合特征参数损失率越大。本研究发现,3种果树滞尘量与Pn、Fv/Fm以及Chl之间呈显著负相关关系或极显著负相关关系。这与吴天忠等[42]研究结果一致,说明植物叶片的光合作用与其所处的环境紧密相关,植物滞尘量增大会减少叶片的光合有效面积,严重阻碍植物的光合作用。
4 结论
三种果树滞尘能力为苹果>枣>核桃,叶面滞尘量越大,对果树叶片光合作用的影响越大。降尘导致3种果树叶片净光合速率下降的主要因素是非气孔因素。降尘使果树叶片PSⅡ反应中心产生一定程度的伤害或可逆性失活,降低了果树叶片的原初光能转换效率,从而影响光合作用的正常进行。
