烟草甲气味受体基因的鉴定和组织表达谱
2022-12-20王桂瑶杨萌萌常延斌苗晨琳赵国庆李玉娥宋纪真
王桂瑶,杨萌萌,常延斌,许 强,苗晨琳,赵国庆,李玉娥,宋纪真*
1. 中国烟草总公司郑州烟草研究院,郑州高新技术产业开发区枫杨街2 号 450001
2. 江苏中烟工业有限责任公司,南京市建邺区兴隆大街29 号 210000
3. 吉林烟草工业有限责任公司,长春市经开区世纪大街99 号 130022
烟草甲Lasioderma serricorne(Fabricius)属鞘翅目(Coleoptera)窃蠹科(Anobiidae),是世界性仓储害虫,具有食性杂、繁殖力强和分布范围广的特点,主要为害贮藏的烟叶及烟草制品、粮食和中草药等[1-2]。已有调查表明烟草甲为害给卷烟企业造成的直接损失率约为0.215%[3],每年给国家带来巨大的经济损失。目前,磷化铝熏蒸是防治烟草甲的主要手段,但长期应用化学防治已引起害虫抗药性和环境污染等问题[2,4-5]。嗅觉对昆虫搜寻食物、寻找配偶和产卵等至关重要[6]。随着分子生物学和基因组学技术的发展,可利用昆虫嗅觉基因高通量筛选对昆虫嗅觉行为具有调节作用的小分子化合物来开发昆虫行为调节剂,此方法相比使用昆虫嗅觉仪筛选昆虫行为调控剂的传统方法[7-10]效率更高,对于烟草甲的绿色防控具有重要意义。
昆虫气味受体是具有反向拓扑结构(N 端位于细胞内,C 端位于细胞外)的G 蛋白偶联受体[11],能够特异性地识别气味分子,是昆虫嗅觉感受中最关键的嗅觉蛋白[11-13]。通过鉴定昆虫气味受体基因,进而研究气味受体基因功能,可以解释昆虫行为产生的分子基础,有助于昆虫行为调控剂的开发[13-14]。已有研究表明,昆虫气味受体主要分为两种:一种是传统气味受体(ORx),它们在不同昆虫间同源性较低且数量各异[15];另一种是气味受体辅助受体Orco(Odorant receptor co-receptor),每种昆虫只有1个,在不同昆虫间保守,能够与ORx 形成配体门控性离子通道(异源二聚体)[12]。例如,利用星天牛(Anoplophora chinensis)触角转录组数据,Wang 等[16]鉴定出43 个ORs基因和1 个Orco基因。Cheng 等[17]从黄野螟(Heortia vitessoides)触角转录组数据中鉴定出27 个ORs基因和1 个Orco基因。在烟草甲中,Wang 等[18]对烟草甲触角转录组进行了测序分析,鉴定出14 个气味结合蛋白、5 个化学感受蛋白和2 个Niemann-Pick C2 蛋白。然而,目前有关烟草甲气味受体基因的鉴定研究还鲜见报道。因此,基于烟草甲触角转录组数据,利用生物信息学手段挖掘鉴定气味受体基因,并分析烟草甲气味受体与其他鞘翅目昆虫气味受体之间的系统进化关系,进而通过荧光定量PCR 分析气味受体基因在烟草甲雌虫和雄虫触角中的相对表达量,旨在增加对烟草甲嗅觉感受系统的认识,一方面为烟草甲气味受体的功能研究奠定基础,另一方面为烟草甲行为调控剂的开发提供理论依据,从而加快引诱剂和趋避剂等行为调控剂在烟草甲绿色防控中的应用步伐。
1 材料与方法
1.1 供试虫源和组织解剖
烟草甲从中国烟草总公司郑州烟草研究院贮存的原烟上采集并在培养箱内用人工饲料(全麦粉和燕麦粉的质量比为7∶3)饲养多代。培养箱的温度为29 ℃±1 ℃,相对湿度为75%±5%,光照条件为全暗。刚羽化的烟草甲被分成雌虫和雄虫,收集大约500 对羽化2 d 的烟草甲成虫用来解剖其触角、腹部和足[19-20]。解剖的组织样品液氮冷冻后保存在-80 ℃冰箱中备用。
1.2 烟草甲ORs基因的挖掘
实验室前期通过高通量测序构建了烟草甲触角转录组数据库,利用关键词odorant receptor 对转录组注释结果进行搜索。基因的开放阅读框通过网 站https://www.ncbi.nlm.nih.gov/orffinder/进 行 预测。搜索到的序列在NCBI(National Center for Biotechnology Information)的Nr 数据库中进行BLASTX 分析,从而确认序列注释的正确性。
1.3 烟草甲ORs基因的系统进化分析
选择烟草甲的9 个气味受体的氨基酸序列,与下载自NCBI 数据库的暗黑鳃金龟(Holotrichia parallela)、绿豆象(Callosobruchus chinensis)、光肩星天 牛(Anoplophora glabripennis)和 赤 拟 谷 盗(Tribolium castaneum)等鞘翅目昆虫氨基酸序列进行系统进化分析。利用Clustal Omega(https://www.ebi.ac.uk/Tools/msa/clustalo/)对这些嗅觉基因进行多序列联配。使用最大似然法,在MEGA 7(JTT model,1 000 bootstrap)中 构 建 系 统 进 化树。使用Figtree 1.4.0 软件对进化树进行可视化编辑。
1.4 烟草甲ORs基因的组织表达谱分析
利用Trizol 试剂(南京诺唯赞生物科技股份有限公司)提取烟草甲雌雄虫触角、腹部和足的总RNA。利用Nanodrop 微量核酸蛋白检测仪(美国赛默飞公司)测定RNA 浓度。利用荧光定量PCR反转录试剂盒(南京诺唯赞生物科技股份有限公司)反转录1 µg RNA 为cDNA。以烟草甲的18SrRNA 和RPS15基因为内参基因,在线设计(http://bioinfo.ut.ee/primer3-0.4.0yi/)所有基因的引物,引物序列见表1。用Light Cycler 480 II 荧光定量PCR 仪(瑞士罗氏公司)进行扩增,使用SYBR 荧光定量专用染料(南京诺唯赞生物科技股份有限公司),PCR反应程序:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火和延伸共30 s,40 个循环;95 ℃15 s,60 ℃60 s,95 ℃15 s 形成融解曲线,通过融解曲线判断引物的特异性。利用2-ΔΔCt方法计算ORs基因在烟草甲雌虫和雄虫触角、腹部和足中的相对表达量,以足为对照,RPS15基因为内参基因,进行3 次生物学重复。用DPS 19.05 软件对数据进行ANOVA(One-way Analysis of Variance)方差分析,利用LSD(Least Significant Difference)法对差异显著性(P<0.05)进行检验。

表1 荧光定量PCR 使用的引物Tab.1 Primers for qRT-PCR
2 结果与分析
2.1 烟草甲ORs基因的挖掘
基于烟草甲触角转录组数据,共鉴定出9 个ORs基因(LserOR1 ~LserOR8,LserOrco)(表2)。这些ORs基因序列具有完整的开放阅读框,氨基酸长度范围为132 ~479 个氨基酸。烟草甲ORs基因在NCBI 进行BLASTX 比对的结果显示,烟草甲的Orco 序列与光肩星天牛的Orco 序列相似度最高(78.29%),而烟草甲其他的ORs 序列与其他昆虫ORs 的匹配度为24.06% ~55.36%。
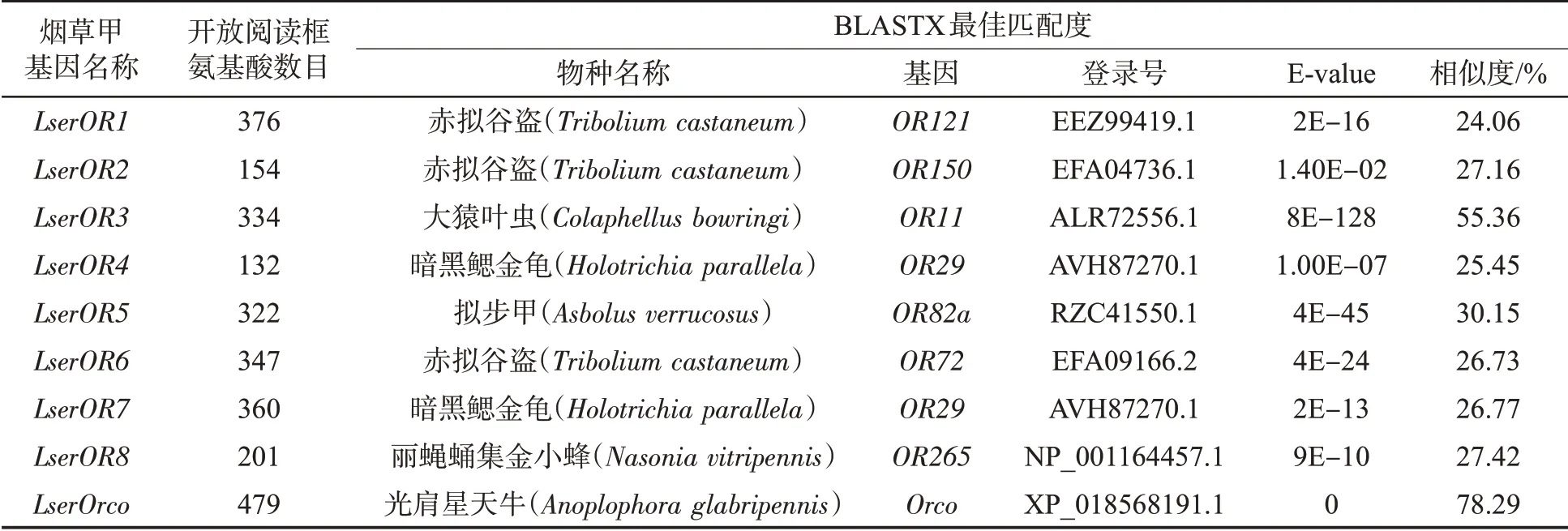
表2 烟草甲触角转录组数据中鉴定的气味受体基因Tab.2 OR genes identified in antennal transcriptome data of L. serricorne
2.2 烟草甲ORs基因的系统进化分析
系统进化分析结果(图1)表明,烟草甲的LserOR6 与光肩星天牛AglaOR22 聚在同一个分支,表明它们之间进化关系更接近。另外,烟草甲的LserOrco 与绿豆象等其他鞘翅目昆虫的Orco 也聚在同一个分支。
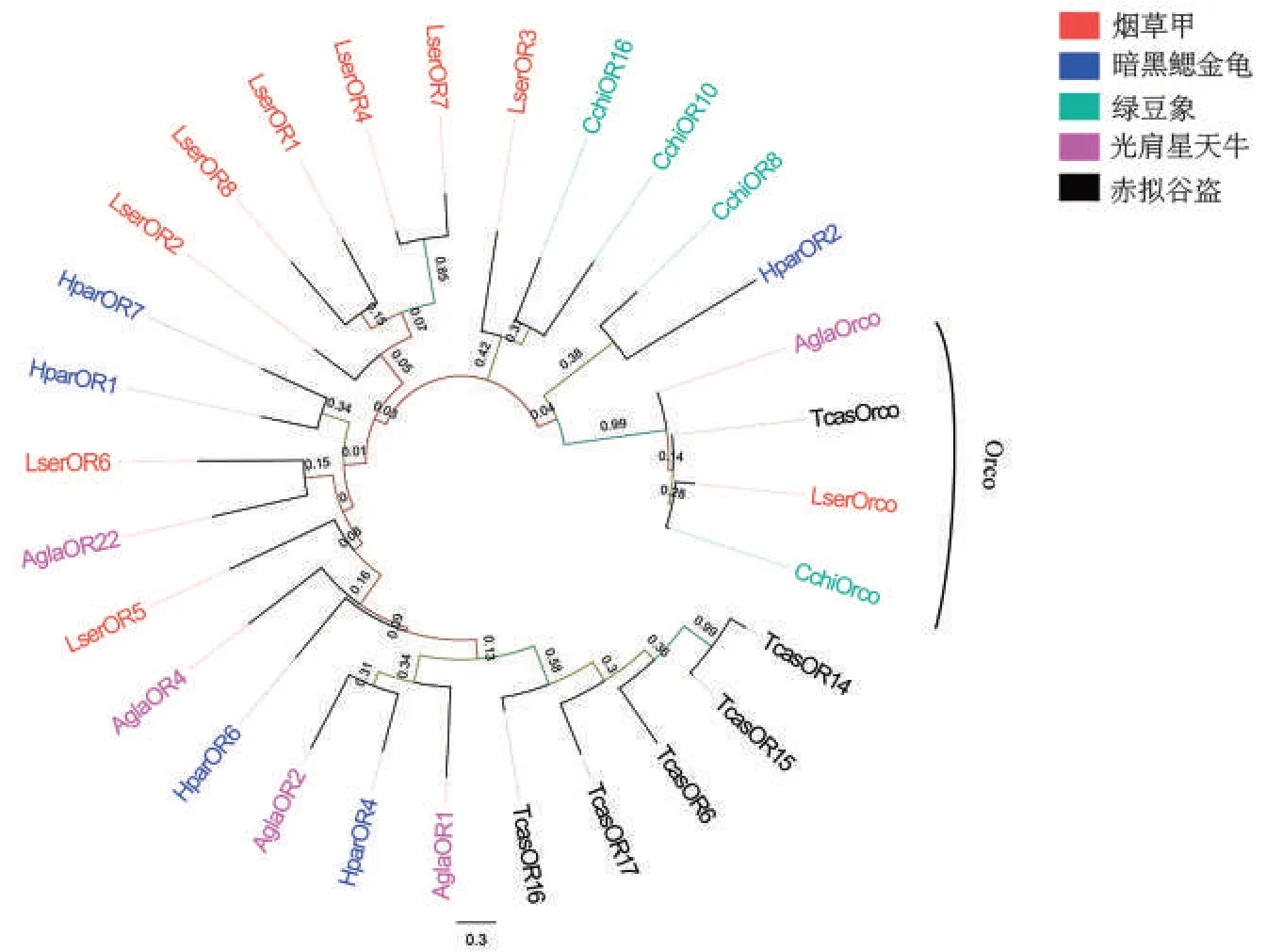
图1 不同鞘翅目昆虫气味受体蛋白的系统进化分析Fig.1 Phylogenetic analysis of ORs from various coleopteran insects
2.3 烟草甲ORs基因的组织表达谱分析
实时荧光定量PCR 结果显示,两个LserORs基因(LserOR5和LserOrco)在烟草甲雌虫和雄虫触角中表达量相对较高,是对照转录本量的3~5 倍(图2)。其中,LserOR5在雄虫触角中的表达量显著高于雌虫,而LserOrco在雌虫和雄虫触角中的表达量无显著差异。另外,LserOR1和LserOR4在雄虫触角中表达量是对照的1.7 倍,而其他气味受体基因(LserOR2~LserOR3,LserOR6~LserOR8)在触角和腹部低表达。
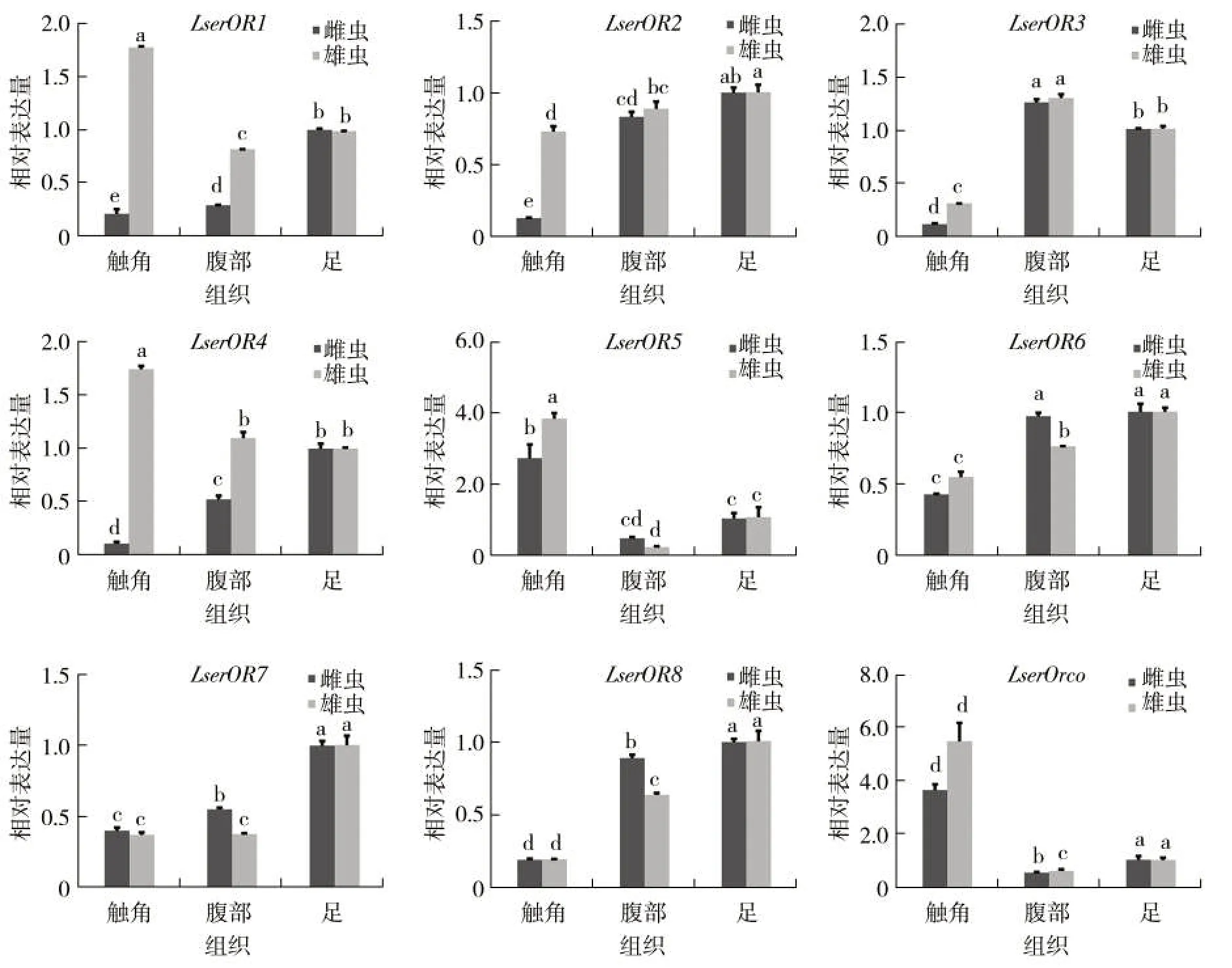
图2 烟草甲气味受体基因在雌雄虫的组织表达谱Fig.2 Tissue expression profiles of L. serricorne OR genes
3 讨论
目前,研究人员已经在多种鞘翅目昆虫中发现了ORs基因[21]。本研究中从烟草甲触角转录组中共鉴定出9 个ORs基因,相比星天牛(44 个ORs)[16]和甘薯小象鼻虫(Cylas formicarius)(54 个ORs)[22]较少。在不同昆虫中鉴定出的ORs基因数目各异,一方面可能是由于昆虫本身的生理学差异[23],另一方面可能是昆虫在特定条件下表达的基因数目有限,因为转录组数据呈现的只是特定时间细胞中表达的嗅觉基因,而缺少一些低丰度或不表达的气味受体基因[24]。下一步可通过烟草甲全基因组测序,更全面地揭示烟草甲气味受体基因信息[25]。
本研究中,烟草甲的ORs 与其他昆虫的ORs 相似度为24% ~55%,这与黑肩绿盲蝽(Cyrtorhinus lividipennis)(28% ~77%)[26]等其他昆虫的研究结果具有一致性,表明ORs在不同昆虫间同源性较低,可能与不同昆虫具有不同的气味偏好有关[12-13]。气味受体蛋白的系统进化分析表明,烟草甲与其他鞘翅目昆虫的Orco 聚在一起,这与甘薯小象鼻虫Orco 蛋白聚类分析结果一致[22],进一步说明Orco序列在不同昆虫间的保守性。另外,烟草甲的LserOR6 与光肩星天牛AglaOR22 聚在同一个分支,说明它们在进化上具有相似性[26-27]。
烟草甲LserOR5和LserOrco基因在触角中表达量相对较高,已有研究表明昆虫触角中一些高表达的ORs和Orco基因与感受植物挥发物和昆虫性信息素密切相关[28-29]。例如,暗黑鳃金龟触角中特异性表达的气味受体HparOR27 能够感受3 种植物挥发物[29]。利用RNAi 技术降低Orco基因在黑肩绿盲蝽[26]和红棕象甲(Rhynchophorus ferrugineus)[30]触角中的表达量,可以使这些昆虫对植物挥发物等多种气味的敏感性下降。在烟草甲中,LserOR5和LserOrco识别气味分子的功能有待通过蛋白体外表达和RNAi等技术进一步研究。另外,本研究中发现一些气味受体基因(LserOR2~LserOR3,LserOR6~LserOR8)在烟草甲成虫触角和腹部低表达,这可能是由于这些基因在烟草甲其他生长发育阶段高表达或具有其他生理功能[13,21],仍需进一步研究。
4 结论
基于烟草甲触角转录组数据,共鉴定出9 个ORs基因(LerOR1~LerOR8,LserOrco)。系统进化树分析表明,烟草甲的LserOR6 与光肩星天牛AglaOR22 聚在同一个分支,它们的进化关系更接近。实时荧光定量PCR 结果显示,烟草甲的两个气味受体基因(LserOR5和LserOrco)在触角中表达量相对较高。上述结果可为烟草甲气味受体基因的功能研究奠定基础,从而促进其引诱剂和趋避剂等行为调控剂的开发。
