过表达三七PnSS 基因对烟草萜类物质含量及生长发育的影响
2022-12-20罗崇玉张利娜李作森杜云龙
罗崇玉,张利娜,姜 茜,周 璇,韩 丽,梁 燕,彭 晟,李作森,杜云龙*
1. 云南农业大学植物保护学院,昆明市盘龙区沣源路452 号650201
2. 云南农业大学烟草学院,昆明市盘龙区沣源路452 号 650201
3. 云南省蒙自市人民政府雨过铺街道办事处,云南省蒙自市尼苏小镇火把路1 号 661111
烟叶的香气和香味是衡量烟叶及其制品质量的重要指标。萜类化合物是香味物质的重要组成部分[1]。萜类化合物通过参与合成次生代谢产物新植二烯、类胡萝卜素和希柏烷类等香气前体物质,可以提高烟叶香气品质[2-4]。植物萜类化合物主要由甲羟戊酸途径(Mevalonic acidpathway,MVA)合成,其代谢途径利用MVA 生成萜类化合物中心前体异戊二烯焦磷酸(Isopentenyl pyrophosphate,IPP),并在IPP异构酶的作用下生成二甲基丙烯基二磷酸(Dimethylallyl Pyrophosphate,DMAPP),IPP 和DMAPP 以头尾缩合的形式合成香叶基焦磷酸(Geranylpyrophosphate,GPP),在单萜环化酶的作用下形成单萜及其衍生物,GPP也可以与1分子IPP在法尼基焦磷酸合酶(Farnesyl pyrophasphate synthase,FPS)催化下反应生成法尼基焦磷酸(Farnesyl diphosphate,FPP),FPP是多萜化合物合成的原料,FPP 也可以在角鲨烯合酶(Squalene synthase,SS)的作用下经过一系列的氧化还原反应得到三萜化合物及其衍生物[3,5]。薄荷中的萜类化合物合成关键基因FPS在烟草中异源过表达后,烟草中8种类胡萝卜素降解产物及茄酮、新植二烯含量均有不同程度的提高[6]。酵母中的FPS基因转入烟草后,烟草中的致香成分甾醇和类胡萝卜素含量显著增加[7]。SS所催化的反应是三萜化合物、甾醇、胆固醇等萜烯类物质合成分支点,其活性高低决定着这些物质的产量[8-9]。角鲨烯合成酶抑制剂Terbinafine处理烟草细胞会抑制烟草中的甾醇的合成[10]。角鲨烯合酶在不同物种间具有较高的同源性,酶活性中心高度保守,外源基因表达可提高相应的酶活性[11-12]。人参SS1的过表达可以增加转基因刺五加[13]和人参[14]中植物甾醇和三萜类物质的含量。烟草中异源表达反义的青蒿SS基因可降低植物甾醇的生物合成,并促进类异戊二烯途径中以FPS为前体的其他代谢支路的生物合成[15]。三七(Panax notoginseng(Burk.)F.H.Chen)属于五加科人参属植物,是中国传统的名贵中药材。过表达三七的角鲨烯合成酶(Squalene synthase fromPanax notoginseng,PnSS)会增加三七中的三萜皂苷含量[16-17],但PnSS 基因对烟草品质的影响,以及能否在香料烟巴斯玛14 号中表达PnSS基因来增加萜类物质含量进而改善烟叶品质尚不清楚。为此,克隆了PnSS基因,并转化烟草品种巴斯玛14 号,旨在为利用三七皂苷合成相关基因调节烟草萜类香味物质含量并改善烟叶品质提供依据。
1 材料与方法
1.1 植物材料
烟草品种巴斯玛14号(Basma No. 14)由云南省烟草农业科学研究院提供,含有PnSS基因的中间载体为本实验室保存。
1.2 生物信息学分析
用 DNAMAN8 进 行PnSS(GeneBank no.KC953032)和NtSS(GeneBank no. NM_001325508)核酸序列和氨基酸序列的比对,用在线网站https://prosite.expasy.org/prosite.html进行功能区域的预测。
1.3 植物表达载体构建
用引物对SSFP1和SSRP1(SSFP1:5'-ACGCGTC GACATGGGAAGTTTGGGGGCAATTC-3',SSRP1:5'-TGGTTCTGCAGTCACTGTTTGTTCGGTAGTA G-3')扩增PnSS基因的目的片段,并进行电泳验证,然后胶回收DNA,通过测序检测再次确认获得的扩增目的片段。 将35S-GFP-terminator插入在pCAMBIA2300载体上的多克隆位点5'-EcoRI /HindIII-3'之间,获得pCAMBIA2300-35S-GFP载体,之后用SalI及PstI切下GFP并插入PnSS基因替换GFP基因,用T4 DNA Ligase 进行连接,获得表达载体pCAMBLA2300-35S::SS,并转化农杆菌GV3101 菌株。载体构建及菌株转化由武汉双螺旋生物科技有限公司完成。
1.4 烟草的转化和阳性株系的鉴定
农杆菌培养至对数生长期(OD600= 0.5),通过叶盘法转化烟草[18]。转化PnSS的烟草在含有500 mg/L羧苄青霉素和50 mg/L 卡那霉素的MS 选择培养基上培养。在筛选培养基上生长的苗长至0.5 cm 时,转移到含有相应抗生素的1/2 MS 固体培养基上诱导生根。
取烟草叶片,利用CTAB法提取烟草DNA,用引物对SSFP2 和SSRP2(SSFP2:5'-ATGGGAAGTTTG GGGGCAATTCTGA-3',SSRP2:5'-TCACTGTTTG TTCGGTAGTAGGTTT-3')扩增PnSS基因。 PCR 程序:95 ℃预变性3 min;95 ℃变性50 s,52 ℃退火30 s,72 ℃延伸90 s,共30 个循环;最后72 ℃延伸10 min。PCR产物用0.8%的琼脂糖凝胶电泳检测。
1.5 RNA提取、逆转录及real-time PCR分析
用EasyPure Plant RNA Kit(TransGen Biotech)试剂盒(北京全式金生物技术有限公司)提取烟草根、茎、叶中的总RNA。对提取的总RNA 进行逆转录合成cDNA,逆转录反应体系:以5 μL总RNA为模板,加入4 μL 5 × TransScript All-in-One SuperMix for qPCR(北京全式金生物技术有限公司)和1 μL gDNA Remover,并加入10 μL RNase-free Water 配置成20 μL的反应体系。该反应体系在42 ℃金属浴上孵育5 min,然后50 ℃反应15 min,75 ℃反应5 min 停止反应,获得cDNA。通过ABI QuantStudio7 Flex(美国Applied Biosystems 公司)进行Real-time PCR 检测烟草植株中PnSS基因的表达水平,Real-time PCR反应体系:以0.5 μL的cDNA为模板,加入5 μL 的Power Up TM SYBRTM Green Master Mix[赛默飞世尔科技(中国)有限公司],0.4 μL的正向引物(PnSS-rFP:5'-TTGCTGAAGTCCAAGGTTG ACA-3')和0.4 μ L 的反向引 物(PnSS-rRP:5'-GGCTGAATTGTGTCCTGACTC-3'),并加入3.7 μL ddH2O 配置成10 μL 的反应体系。Real-time PCR 反应程序:95 ℃预变性2 min;95 ℃变性45 s,56 ℃退火30 s,40 次循环;72 ℃延伸1 min。以NtActin1基因作为内参基因(NaActin-rFP:5'-GGTCGTACCAC CGGTATTGTG-3',NaAtin-rRP:5'-GTCAAGACGGA GAATGGCATG-3')。进行3次生物学重复,采用2-ΔΔCt计算基因的相对表达量。使用SPSS 26.0 软件分析基因的表达量差异。
1.6 香味物质含量检测
巴斯玛14 号植株(野生型,OT5)和过表达PnSS基因的巴斯玛14号植株(转基因植株,PnSS)在生长4个月后,从植株顶部依次往下取第3~6片叶,单个株系所取样品混合后在40 ℃烘箱中烘干,研磨成粉末,用0.42 mm筛子过滤残渣,根据文献[19]中的方法,采用456GC-TQ 固相微萃取-气相色谱-串联质谱联用仪(美国Bruker公司)检测烟叶中的香味物质含量(质量分数)。
1.7 数据分析
对烟草植株株高和叶片大小的差异进行t检验。采用2-ΔΔCt分析转基因烟草根、茎和叶中的PnSS基因的表达量,使用软件SPSS 26.0分析基因的表达量差异。
2 结果与讨论
2.1 PnSS基因的生物信息学分析
PnSS基因与烟草中的同源基因NtSS序列分析结果(图1)显示,PnSS与NtSS的核苷酸序列具有66.1%的一致性,氨基酸序列具有81.9%的一致性,并且都具有两个角鲨烯和植烯合成酶(Squalene and phytoene synthases)结构域。

图1 PnSS与NtSS氨基酸序列比对和功能区分析Fig.1 Amino acid sequence alignment and functional region analysis of PnSS and NtSS
2.2 过表达PnSS基因对烟株生长发育的影响
PnSS与NtSS具有较高的一致性(图1),将含有PnSS基因的植物表达载体(图2)通过农杆菌介导的方法转化到烟草中,获得5株在筛选培养基上生长的转化PnSS基因的烟草。随机挑选4株经抗生素筛选成功的转化PnSS基因的烟株,通过PCR检测转基因植株中的PnSS基因,获得2株含有PnSS基因的烟草植株(图3A)。
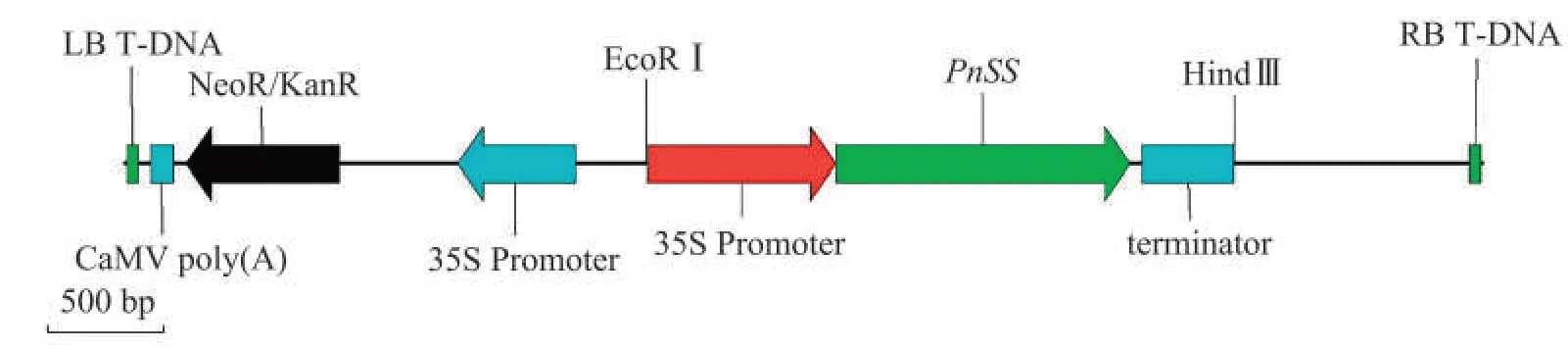
图2 含有PnSS基因的表达载体示意图Fig.2 Schematic diagram of expression vector containing PnSS gene
对含有PnSS基因的转基因烟草进一步进行植株表型观察。表型结果(图3A~图3D)显示,与野生型植株(n=11)相比,转基因烟草植株(n=14)变矮,叶片变小(图3A~图3D)。同时,在转基因烟草植株的根、茎和叶中,PnSS基因在叶片中的表达量最高(图3E)。表明过表达PnSS基因会抑制烟草植株的生长发育。
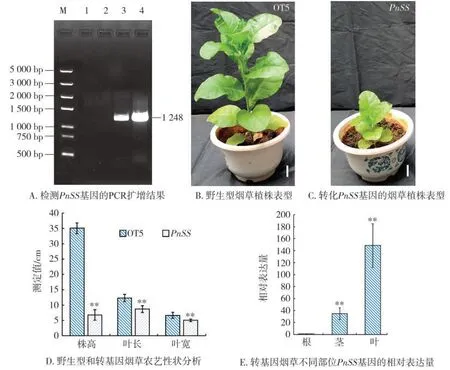
图3 PnSS基因的过表达对烟草植株生长发育的影响Fig.3 Effects of overexpression of PnSS gene on growth and development of tobacco plant
2.3 过表达PnSS基因对烟草中香味物质含量的影响
转基因烟草叶片中香味物质含量检测结果(表1)表明,PnSS基因的过表达使烟叶中乙酸、糠醇、4,6-二甲基嘧啶、苯乙醛、3,5-二甲基苯甲醛、二烯烟碱、二氢猕猴桃内酯、2-甲基吡嗪、壬酸、异戊醛、2-甲基丁醛、西松烯和烟碱13种香味物质含量增加,2,3-二氢-3,5二羟基-6-甲基-4(H)-吡喃-4-酮、2,3-联吡啶、植醇、苯乙醇、新植二烯、苯甲醛和麦斯明7种香味物质含量降低。表明PnSS基因的过表达影响了烟叶中香味物质含量。

表1 烟叶样品中香味物质的含量Tab.1 Relative contents of aroma components in tobacco leaves
3 讨论
萜类香味物质在烟草品质评价中具有重要作用。植物中SS基因和FPS基因是萜类物质合成途径中的关键基因,FPS基因位于SS基因上游[3,5,20]。在本研究中发现,过表达PnSS基因会抑制烟草植株的生长发育,这与前人的研究结果一致,茶树FPS基因在烟草中异源过表达也会抑制烟草植株的生长发育[21],这显示萜类合成基因调控了植株的生长发育。但萜类物质如二烯烟碱、植醇、新植二烯的含量变化如何影响过表达PnSS烟草植株的表型还需进一步试验。
本研究中对烟草香味物质含量变化的分析发现,在过表达PnSS基因的烟草中新植二烯含量减少,这与前人的研究结果不一致。薄荷FPS基因在烟草中过表达会导致烟叶中新植二烯含量增加[6],这暗示在烟草中过表达PnSS基因时PnSS基因可能参与了不同代谢途径,从而导致新植二烯含量发生变化,但烟草中FPS基因是否与PnSS基因共同调控烟草中新植二烯等萜类物质的合成还需进一步深入研究。
4 结论
三七PnSS 的氨基酸序列与烟草中同源物具有高度一致性,在烟草中过表达PnSS基因后,PnSS基因的表达量在叶片中最高,同时过表达PnSS基因的烟草植株变矮、叶片变小、叶片中与香味有关的萜类物质含量发生改变。因此,PnSS基因可影响烟叶中萜类物质等香味物质的合成并参与调控烟草植株的生长发育。
