番茄潜叶蛾成虫卵巢发育与卵子发生研究
2022-12-19梁永轩郭建洋张毅波张桂芬吕志创刘万学
梁永轩,郭建洋,王 浩,张毅波,周 琼,张桂芬,吕志创,刘万学*
(1.中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193;2.湖南师范大学生命科学学院,长沙 410081)
番茄潜叶蛾Tutaabsoluta(又名Phthorimaeaabsoluta),隶属于鳞翅目Lepidoptera麦蛾科Gelechiidae,起源于南美洲秘鲁,于2006年由原产地入侵欧洲的西班牙后,迅速传播到欧洲中部地区、地中海沿岸等非洲国家和亚洲的部分国家[1-3]。截至2017年,该害虫已入侵80多个国家和地区,成为世界番茄重要入侵害虫,番茄受害面积约占全球番茄种植面积的一半以上,每年直接经济损失约60亿美元。对全球番茄产业造成了严重影响[4]。我国番茄种植面积位居世界第一,是全球番茄产量最大的国家,占全球总产量的32.6%[5]。番茄潜叶蛾自侵入我国西北和西南的多个邻国后[6-7],于2017年8月抵达我国新疆伊犁为害当地露地番茄,次年3月,又在云南临沧发现其为害保护地番茄[8-9]。风险评估表明,该虫在我国大部分番茄种植区都可以存活和定殖,综合风险评估R值为2.64,属于极高风险性农业有害入侵生物,对我国番茄等作物的潜在威胁极大[6]。
番茄潜叶蛾最喜寄主为番茄、茄子[10]等茄科植物,以幼虫在叶片内取食为害,留下窗纸样、不规则的潜道或潜斑,还能直接蛀入果实内部[10-11],导致使用化学农药难以起到理想的防控效果。而研究表明,除茄科外,该虫还为害或取食包括豆科、锦葵科、苋科、旋花科、藜科、菊科、十字花科以及禾本科在内的11个科近50种植物[4,8,12],生存与繁殖能力很强[1,4]。因此,明确其生殖生物学特性并开展早期的预测预报,对于制定有效的防控策略极为重要。
雌性昆虫的生殖系统具有产生和贮存卵子、完成受精和产卵等功能。卵子发生、成熟与受精后产出体外[13]。卵子发生主要通过卵母细胞吸取大量的营养物质,这些物质既有本身合成的,也有从外界摄取的,这些物质沉积在卵内为胚胎发育提供营养[14]。通过分析卵巢发育、卵子发生可以为准确预测预报害虫种群动态、确定害虫防治适期等提供参考,在害虫发生高峰期前采取有效措施,以提高防治效果[15-16]。因此,明确番茄潜叶蛾雌成虫的生殖系统结构,探明该虫的卵巢发育和卵子发生过程,对于野外种群发生期和发生量的预测预报等具有重要的指导意义[17-18],而目前未见有针对番茄潜叶蛾的此类研究。对此,本研究分析了雌成虫的生殖系统结构与组成,明确了成虫发育过程中卵巢的形态变化,补充营养(20%葡萄糖溶液)、交配与否等对其卵巢发育、卵子发生过程的影响,以期为该虫的种群预测预报和防控策略提供支撑。
1 材料与方法
1.1 供试虫源
番茄潜叶蛾于2019年7月采自云南省玉溪市红塔区温室大棚的‘大粉’番茄苗。温室条件:温度(26±1)℃,相对湿度(60±5)%,自然光照。
1.2 成虫的取样及处理
幼虫化蛹后雌、雄分装于1.5 mL透气离心管中继续生长发育,离心管置于人工气候箱,饲养条件为:温度(25±1)℃,相对湿度(60±5)%,L∥D=16 h∥8 h,取每天晚上(20:00-22:00)羽化雌成虫用于试验,初羽化成虫转移到50 mL的特制离心管。分别设置交配+清水、交配+20%葡萄糖溶液、未交配+清水和未交配+20%葡萄糖溶液4个处理,其中交配处理选与雌成虫同一天羽化的雄成虫按1∶1配对,观察到雌雄交配的作为有效处理组。成虫一羽化立即用脱脂棉球蘸取清水或20%葡萄糖溶液并包裹番茄叶片叶柄,放入离心管中。各处理离心管置于人工气候箱中,每天下午16:00补充清水或20%葡萄糖溶液。
1.3 雌成虫生殖系统的解剖与观察
分别随机选取初羽化雌成虫、1~6日龄取食20%葡萄糖溶液的交配或未交配雌成虫解剖并观察卵巢结构。首先将雌虫在-20°C冰箱中冷冻处理10~15 min,然后将晕厥的成虫置于滴有磷酸盐缓冲液(PBS)的载玻片上,于体视显微镜(型号:Stemi 508,ZEISS)下去除体壁仅留下腹内组织,去除消化道、腹神经索、气管等组织,保留生殖系统与脂肪体。
根据脂肪体数量与颜色、卵黄沉积水平、卵子发生状况、卵巢管发育水平等[19-21],并参考张万娜等对棉铃虫Helicoverpaarmigera[21]、刘宁等对苹果蠹蛾Cydiapomonella[22]的卵巢分级方法,对番茄潜叶蛾的卵巢发育状况进行分级,以评价其发育程度。最后去除脂肪体,采用荧光体视显微镜(型号:SZX 16,Olympus)观察和拍摄生殖系统图片。
1.4 不同处理对雌成虫卵子发生的影响
按1.2小节设置处理。分别选取6头初羽化,以及每种处理羽化24、48、72、96 h的雌成虫在体视显微镜(型号:Stemi 508,ZEISS)下解剖,并记录卵巢内外的卵子数。
此外,以未交配+清水条件下羽化96 h的雌成虫为试虫,进一步设置2种处理,即分别继续提供清水或补充20%葡萄糖溶液,于羽化120、144 h时再取6~8头进行解剖,并记录卵子数。
1.5 数据处理与统计分析
采用SPSS Statistics 20软件进行数据分析。用双因素方差分析(Two-way ANOVA analysis,LSD test)检验羽化后相同时间,不同处理对番茄潜叶蛾卵子发生数量影响的显著性;用单因素方差分析(One-way ANOVA analysis)和最小显著差数法(LSD test)检验羽化后不同时间之间,卵子动态变化的差异显著性;未交配+清水条件下羽化后96 h继续提供清水或补充20%葡萄糖溶液2种处理对卵子发生数量影响的显著性采用t测验。
2 结果与分析
2.1 雌成虫生殖系统结构与卵巢发育分级
对不同日龄雌成虫生殖系统的观察结果表明,其生殖系统主要包括:1对卵巢、1对侧输卵管(图1e:A)、1根中输卵管(图1e:B)、1对附腺(图1e:C)、1个交配囊(图1e:D)、1个阴道支囊(图1e:E)、1个受精囊(图1e:F)及其受精囊腺(图1e:G)。卵巢整体呈乳黄色,左右对称排列。每侧的卵巢管与同侧的侧输卵管相接,侧输卵管汇合成一条中输卵管,中输卵管末端开口于生殖腔,其侧向分支形成的管道末端附着有受精囊、交配囊等(图1e)。大多数雌成虫卵巢两侧的卵巢管数目相等,均为4根(图1);仅个别成虫两侧的卵巢管数不等,出现一侧为4根,另一侧为5根的现象,占总计观察数量的1.5%(图1f)。
根据脂肪体的数量与颜色、卵黄沉积水平、卵子发生状况、卵巢管发育水平等情况,将卵巢发育划分为5个级别[21-22]。Ⅰ级:卵黄沉积前期。为初羽化时的卵巢,此时卵巢内卵子偶有卵黄沉积,基本为未成熟卵子,总卵子数少;卵巢管细而短,卵巢管及输卵管乳白色,输卵管及中输卵管较细且明显。卵巢周围脂肪体乳黄色,量多而饱满,充满腹腔(图1a)。Ⅱ级:卵黄沉积期。卵巢管端部可见未成熟卵子,成熟卵子开始出现,在卵巢管基部成堆,卵巢管膨大,总卵子数较少,中输卵管较细,侧输卵管内充满卵子而膨大;脂肪体乳黄色,量多(图1b)。Ⅲ级:成熟待产期。卵子数量迅速增多,排列整齐,成熟卵在卵巢管基部成堆,由于卵量的增加和卵的膨大,卵巢管膨胀,卵巢体积明显增大;侧输卵管内充满卵子,中输卵管内亦开始出现成熟卵子;卵巢周围脂肪体黄色,量仍较多(图1c)。Ⅳ级:产卵盛期。卵子量继续增多,达到卵巢最大抱卵量,卵巢管膨大、变长,卵巢达到“饱满”的状态;成熟卵子充满卵巢端部、侧输卵管、中输卵管,排列紧密;脂肪体减少,呈黄色(图1d)。Ⅴ级(产卵末期):卵巢管内成熟卵子和未成熟卵子数量明显减少;卵巢管明显变短,卵巢体积减小;卵巢周围脂肪体剩余量少(图1e)。
2.2 不同处理条件下雌成虫的卵子发生动态
未交配+清水条件下,成虫初羽化(0 h)至羽化后24 h,卵巢内成熟卵子逐渐增加,24~72 h,成熟卵子、总卵子数迅速增加,未成熟卵子数增长平缓,72 h时各类卵子数均达到高峰;72~96 h,观察到部分卵子有重吸收现象,羽化后96 h,成熟卵子(F=11.50,P=0.006 9)、未成熟卵子(F=28.12,P=0.000 3)及总卵子数(F=33.54,P=0.000 2)相比72 h均显著减少(图2a)。
未交配+20%葡萄糖溶液条件下,0~72 h,卵巢内成熟卵子、总卵子数持续增加,而未成熟卵子数在0~48 h持续减少,48~72 h略有增加。在72~96 h,成熟卵子数仍略有增加(图2b)。成熟卵子和总卵子数均于96 h时达到峰值,数量分别为(96.3±6.9)粒和(242.7±10.4)粒(表1)。
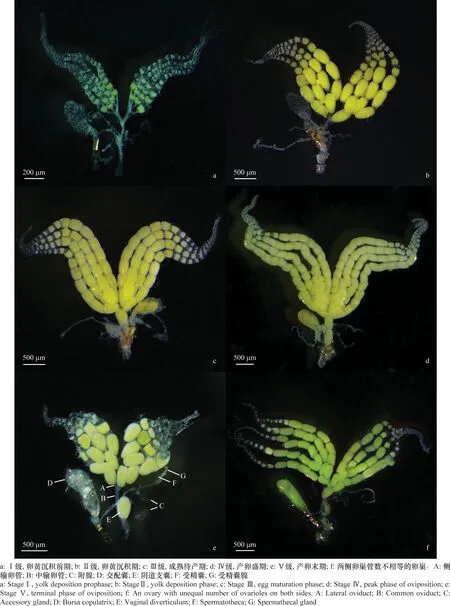
图1 番茄潜叶蛾卵巢不同发育程度的形态和级别划分Fig.1 Morphology and grading criteria of the ovaries of Tuta absoluta at different developmental stages

图2 不同交配状态及补充营养处理的番茄潜叶蛾雌成虫卵子发生动态Fig.2 Oogenesis dynamics of Tuta absoluta female adults under different mating conditions and nutritional supplements

表1 不同交配状态及补充营养处理的番茄潜叶蛾雌成虫的卵子数量比较1)Table 1 Comparison of eggs number of Tuta absoluta female adults under different mating conditions and nutritional supplements
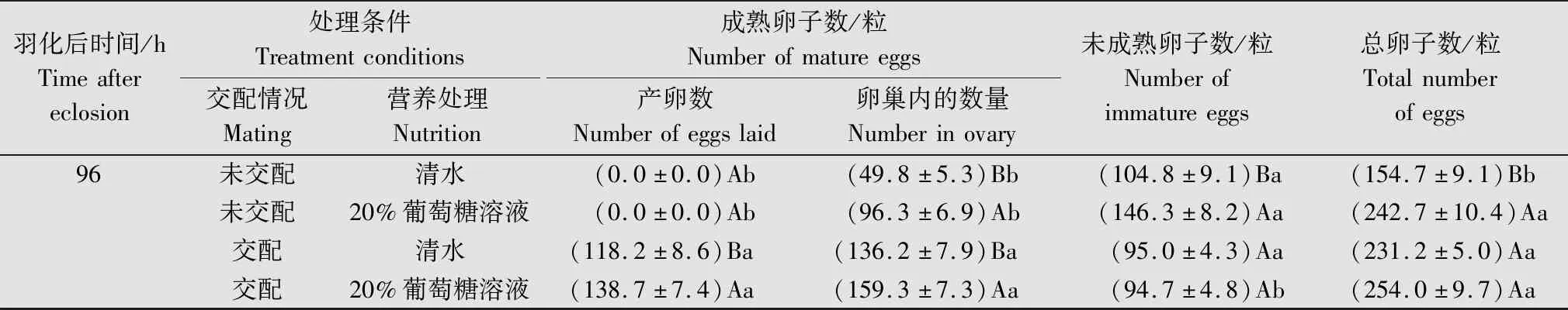
续表1 Table 1(Continued)
交配+清水条件下,成虫羽化后0~96 h,成熟卵子和总卵子数呈较快的增加趋势,而未成熟卵子数则持续减少(图2c)。雌虫与雄虫交配后,在羽化后逐渐将成熟卵子产到体外,至96 h时,成熟卵子、总卵子及产卵数均达最大值,分别为(136.2±7.9)、(231.2±5.0)和(118.2±8.6)粒(表1);而未成熟卵子数减少到最小值。
交配+20%葡萄糖溶液条件下,0~96 h,成熟卵子、总卵子和产卵数变化趋势与交配+清水处理基本一致,于96 h达最大值,分别为(159.3±7.3)、(254.0±9.7) 粒和(138.7±7.4)粒(表1);而未成熟卵子数在0~24 h相对稳定,24~48 h之间减少,于96 h减少到最小值(图2d)。
在未交配+清水条件下雌虫发生卵子重吸收现象(96 h)后,若继续提供清水,则直到羽化后120 h,卵巢内成熟卵子、未成熟卵子、总卵子数变化相对稳定;144 h后均减少到最小值,分别为(45.3±6.0)、(99.3±12.0)粒和(144.7±13.7)粒(表2)。而当卵子重吸收后,给雌虫持续补充取食20%葡萄糖溶液,则羽化120 h时,各类卵子的数量均显著增加,到144 h时略有减少(图3)。

图3 未交配+清水处理下番茄潜叶蛾雌成虫卵重吸收后提供清水或补充葡萄糖后的卵子发生动态Fig.3 Ovogenesis dynamics of Tuta absoluta female adults after oosorption under unmated + water treatment after water supply or 20% glucose solution supplementation
2.3 补充营养及交配与否对卵子发生的影响
取食20%葡萄糖溶液对雌虫卵子发生有明显的促进作用。羽化后24 h,未交配雌虫形成的成熟卵子(F=21.61,P=0.000 2)、未成熟卵子(F=4.92,P=0.038 3)、总卵子数(F=9.73,P=0.005 4),及交配雌虫的成熟卵子(F=19.94,P=0.000 2)、总卵子数(F=6.84,P=0.016 6)均显著高于清水对照组;羽化后72 h,交配雌虫的未成熟卵子数(F=5.33,P=0.031 8)显著高于清水组;而96 h,未交配雌虫的成熟卵子(F=22.59,P=0.000 1)、未成熟卵子(F=17.91,P=0.000 4)、总卵子数(F=49.59,P<0.000 1),及交配雌虫的成熟卵子数(F=5.61,P=0.028 1)均高于清水组(表1)。
在未交配且仅取食清水的条件下,发生卵子重吸收(96 h)后给雌虫补充20%葡萄糖溶液,于120 h时成熟卵子(F=11.40,P=0.007 1)、未成熟卵子(F=5.92,P=0.035 2)和总卵子数(F=20.12,P=0.001 2)均显著增加;120~144 h各类卵子的数量相对稳定。另外,发生卵子重吸收后雌虫补充取食20%葡萄糖溶液与继续取食清水相比,120 h时成熟卵子(t=-3.88,P=0.003 1)、未成熟卵子(t=-3.23,P=0.009 0)和总卵子数(t=-6.04,P=0.000 1),及144 h时的成熟卵子(t=-3.26,P=0.008 6)、未成熟卵子(t=-2.24,P=0.049 0)和总卵子数(t=-3.79,P=0.003 5)均显著多于继续取食清水的同日龄雌虫(表2)。而未交配取食20%葡萄糖溶液的雌虫卵巢内成熟卵子的数量在羽化后72~96 h仍有增加的趋势;羽化后96 h,取食20%葡萄糖溶液雌虫卵巢内成熟卵子(F=22.59,P=0.000 1)、未成熟卵子(F=17.91,P=0.000 4)、总卵子(F=49.59,P<0.000 1)的数量也均显著多于取食清水的对照组(表1)。
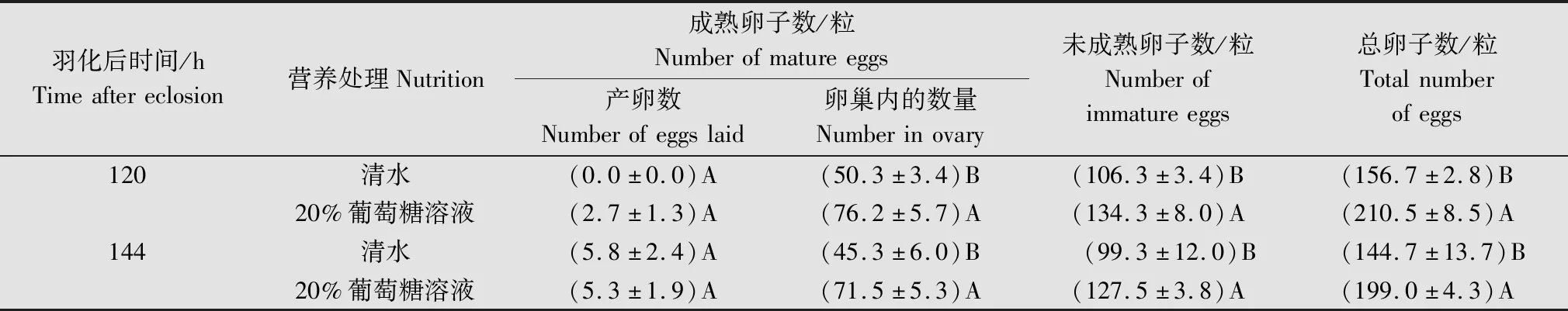
表2 未交配+清水处理下番茄潜叶蛾雌成虫卵重吸收后提供清水或补充葡萄糖后的卵子数量比较1)Table 2 Comparison of eggs number of Tuta absoluta female adults after oosorption under unmated + water treatment after water supply or 20% glucose solution supplementation
从交配与否对卵子发生的影响来看,羽化后96 h,交配雌虫取食清水形成的成熟卵子(F=77.88,P<0.000 1)、总卵子数(F=37.48,P<0.000 1)和取食20%葡萄糖溶液时的成熟卵子(F=41.47,P<0.000 1)均显著多于未交配雌虫相应的对照组。交配雌虫羽化后24~48 h开始产卵,在96 h时,交配雌虫取食清水、20%葡萄糖溶液产卵数分别已达(118.2±8.6)、(138.7±7.4)粒(表1);而未交配雌虫取食清水96 h后补充取食20%葡萄糖溶液,在羽化后120、144 h才有少量产卵,数量分别为(2.7±1.3)粒和(5.3±1.9)粒;未交配持续取食清水的雌虫在144 h才开始产卵,数量为(5.8±2.4)粒(表2)。
3 结论与讨论
昆虫物种卵巢结构相似,主要由一对卵巢、中输卵管、侧输卵管等组成,但昆虫雌虫卵巢内卵巢管数量差异很大,如鞘翅目的莲草直胸跳甲Agasicleshygrophila每侧卵巢管数多为12~16根[22],异色瓢虫Harmoniaaxyridis每侧卵巢管数为25~33根[24],而膜翅目的红颈常室茧蜂Peristenusspretus的卵巢主要由6~10对卵巢管组成[25]。卵巢管含有不同数量、不同排列方式的卵子,前人根据卵巢管中卵子的滋养细胞有无和排列方式,将卵巢管分为无滋式(panoistic type)、端滋式(telotrophic type)和多滋式(polytrophic type)等3种模式[26]。本研究发现,番茄潜叶蛾卵巢管的数目基本为4对,与棉铃虫、苹果蠹蛾、甜菜夜蛾Spodopteraexigua等鳞翅目昆虫相似[27,20-22];体视镜下可见卵巢管内卵母细胞与滋养细胞交替出现,卵室内一个卵母细胞可伴随着多个滋养细胞,卵巢管类型与鳞翅目昆虫的相符,为多滋式[21]。虽然观察发现个别雌虫卵巢两侧的卵巢管数不等,但这一现象可能与雌虫个体大小或其在幼虫期所摄取营养量多寡有关,在其他昆虫中也存在类似现象,如美洲斑潜蝇Liriomyzasativae两侧卵巢管总数多为13~15根,且左右两侧卵巢管数通常不等[28];孟氏隐唇瓢虫Cryptolaemusmontrouzieri每侧卵巢分别含11~14根卵巢管,两侧卵巢管数量也多不相等[29]。
卵巢发育级别在害虫预报中是一项重要指标,可以预测害虫的发生期和发生量。在鳞翅目的重要农业害虫如玉米螟、棉铃虫、苹果蠹蛾、草地贪夜蛾Spodopterafrugiperda等卵巢发育的研究中,常将卵巢发育分为5个级别[19,21-22,30]。本研究在以往卵巢发育分级理论的基础上,根据番茄潜叶蛾卵巢自身特点,将其雌成虫的卵巢分为卵黄沉积前期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)和产卵末期(Ⅴ级)5个级别(图1),并对各发育时期卵巢的脂肪体数量与颜色、卵黄沉积、卵子发育及卵巢管发育情况等加以描述,可为快速、准确判断其卵巢的发育级别提供依据。
基于初羽化雌虫卵巢中成熟卵的数量可将昆虫分为卵熟型(proovigenic)和卵育型(synovigenic)两种类型。卵熟型指雌虫羽化时卵巢中的卵子已全部成熟,成虫阶段不需要再获取用于卵熟的营养物质[31]。而卵育型是指雌虫羽化时卵巢中没有或只有部分成熟卵,雌虫需要继续取食用于满足卵子成熟及持续产卵的营养[32],如轩景丽等研究发现美洲斑潜蝇初羽化雌成虫体内无卵子,且饥饿条件下几乎不能形成成熟卵子[28]。本研究显示,番茄潜叶蛾成虫初羽化时,卵巢未发育成熟,卵巢内仅有未成熟卵;成虫仅取食清水时成熟卵子、总卵子数也可达到较高水平,不需要通过摄入外界营养用于卵子发生、成熟与维持生命,属于卵育型昆虫。
虽然番茄潜叶蛾的卵子发生和卵子成熟无需依赖外界营养,但取食20%葡萄糖溶液对其卵子发生有明显的促进作用,且即便缺乏额外营养的成虫在发生卵子重吸收后更换为取食20%葡萄糖溶液,与继续取食清水的个体相比,可观察到其腹内有更多的脂肪体。可能是因为葡萄糖直接提供了维持生命的能量,减少了雌虫本身的资本营养如脂类物质的消耗,从而使雌虫将资本营养优先用于卵子发生和卵子成熟,且暂不需要重吸收卵子以转化为维持生命的能量[33-34]。因此,在实验室养殖中可通过补充葡萄糖来促进其种群增殖。但葡萄糖是否还直接参与了该虫卵子发生所需脂类物质等的合成还有待明确。李栋等对番茄潜叶蛾的研究结果显示,在实验室条件下,与雄虫两两配对的雌虫在25°C和30°C时,寿命平均约9.24 d和7.87 d[35]。而本次试验中时间梯度到144 h,此时雌虫仍存活且腹内还剩有脂肪体,当羽化时间更长,雌虫资本营养完全消耗且仍存活时,葡萄糖的补充能否合成用于卵子发生的脂类物质从而使雌虫可继续形成卵子值得进一步探究。
研究结果显示,交配处理组,番茄潜叶蛾雌成虫羽化后24~48 h开始大量产卵,而未交配时最早在120 h才见有少量产卵。且未交配+清水处理组,72 h时雌成虫卵巢内成熟卵数量(抱卵量)达到高峰,96 h时由于部分卵子发生了重吸收而显著减少。可能与鳞翅目昆虫的生存策略有关,雌虫羽化时将营养优先用于繁衍,形成一定数量的成熟卵子,交配受精后将可成熟的卵子大量产出;而未受精卵不能正常孵化。卵子的重吸收现象被认为是昆虫在面临环境胁迫时,为了继续存活以便在胁迫解除时能继续繁衍后代的一种生存策略[32]。番茄潜叶蛾雌成虫也存在卵子重吸收现象。因此,雌虫短时间内没有选择把卵子产出,而是将其转化为维持生命的备用营养物质并保存于体内,根据实际需求,如一段时间出现资源缺失或不足或缺乏合适的产卵场所、无法交配等情况时,通过对其卵子的重吸收来继续存活以便将来完成有效繁衍。类似的现象在其他物种的生殖发育研究中也有报道,如潜蝇姬小蜂Diglyphusisaea、印巴黄蚜小蜂Aphytismelinus等[33-34]。可以明确的是,雌虫羽化后越早获得交配机会,其卵子发生越早、成熟更快且产卵累计量更大。因此,在田间及时采取有效的防控措施以减少雌雄虫交配的机会尤为重要,如诱捕法、迷向法等措施的应用[36-38]。
