基于种子发芽率评估长芒苋在我国不同纬度地区的入侵风险
2022-12-19曹晶晶刘万学万方浩郭建英
曹晶晶,王 瑞,刘万学,万方浩,郭建英
(中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193)
随着国际经济贸易的发展,许多外来物种随着人类活动有意或无意地传入多个国家和地区,这些物种一旦成功定殖势必会给当地的生态系统和农业生产造成危害,从而形成外来生物入侵[1]。外来生物入侵是由传入、定殖、扩散和暴发危害等多个阶段构成的一个动态有序的生态学过程[2]。基于生物入侵过程的时序性和阶段性,在入侵的早期阶段及时进行监测预警和预防控制是减少危害的关键,外来生物入侵的防控重点应该集中在防范未传入物种的传入和控制已传入种群的早期定殖两个阶段[3]。由于许多外来物种可通过国际贸易、旅游携带等多种途径传入我国的不同地区,亟须准确评估其在传入地区建立可持续自我繁殖种群的风险,以进行精准防控[4]。
我国地域辽阔,南北跨度大,气候环境复杂多样,具有热带、亚热带、温带等多种气候带。物种,尤其是外来物种通常具有特定的环境适应范围气候带,一旦传入适宜环境就会造成危害。我国气候环境呈纬度地带性分布,导致外来物种的入侵风险也具有纬度地带性差异。外来植物入侵不同纬度地区的风险是由传入种子量、传入后种子发芽率、成苗率、存活率以及繁殖产生的种子量等共同决定的[5]。但是,目前的研究主要是采用物种分布模型,基于已知分布点的气候环境参数,预测适生区等来评估外来植物的广义入侵风险,缺少试验数据支撑的生活史各阶段主导的入侵风险评估[6]。根据外来植物的生活史过程,种子的发芽率是决定外来植物入侵风险的首要因素[7-8]。已有的研究表明,外来植物传入不同地区尤其是不同纬度地区其种子发芽率存在差异[7-8]。因此,系统研究外来植物种子传入不同纬度环境中的发芽率,对定量评估外来物种的入侵风险、进而制定针对性的管理措施具有重要意义。
外来植物种子的发芽率会受到传入地气候环境和种子自身特性的影响[9-10]。通常情况下,气候环境和种子自身生物学特性间是存在互作效应的,如气候环境既可以直接影响入侵植物的发芽率,也可通过改变种子的生物学特性来影响发芽率[10]。因此,气候对外来植物种子发芽率的影响可体现在两个方面:一是温度、水分等直接影响种子尤其是首次传入种子的发芽率。如同一来源的种子传入不同地区后,其发芽率会因气候环境的差异而不同。二是气候会通过影响传入种群在本地繁殖产生种子的千粒重等生物学特性变化间接影响后代种子的发芽率。气候与种子自身生物学特性间的这种互作可导致本地适应性进化,从而提高外来植物的入侵性[11]。如有研究发现已经传入种群产生的种子通常比最初传入种子的发芽率高,说明入侵种群可通过调控种子发芽率来提高其在入侵地的适应能力和入侵性[9]。因此,系统研究外来植物在不同纬度环境下发芽率的变化,明确纬度、种子生物学特性及它们之间的互作效应对种子发芽率的影响,对于评估外来植物在异质环境下的入侵风险具有重要意义。
长芒苋AmaranthuspalmeriS.Watson是苋科苋属异株苋亚属一年生草本植物,竞争能力强、危害性大,是具有多种除草剂抗性的检疫性有害“超级杂草”[12-13]。长芒苋原产于北美洲,最早于20世纪80年代传入我国北京市丰台区[14]。目前,已在我国从南到北不同纬度的多个地区定殖,部分传入点并不在预测的适生区范围内[15]。长芒苋能否在我国的不同纬度地区形成入侵并造成危害是当前亟须解决的问题。理论上,不同纬度下,气候环境条件的差异会影响长芒苋种子传入后的成功定殖与可持续种群的建立,种子发芽率又是决定种群定殖与繁殖增长的入侵风险的首要条件。此外,在高纬度地区,由于温度等因素的影响,存在外来植物传入后种子虽能发芽,但不能完成生活史的情况。我们野外调查时发现长芒苋在我国东北地区盛夏发芽的种子大多数不能完成生活史。因此,对于不同纬度地区,长芒苋能够传入并形成种群的入侵窗口期存在差异。基于此,本研究通过对中国11个不同纬度入侵的长芒苋种群和原产地美国种群的种子发芽率进行比较,通过分析纬度环境与发芽率的关系,明确种子发芽率是否随纬度发生快速适应,评估长芒苋在不同纬度地区的入侵窗口期和入侵风险,为制定区域化的精准防控措施提供指导。
1 材料与方法
1.1 种子样品采集
从2012年至2018年,在我国11个省市采集了长芒苋的成熟种子样品(表1),2019年初又在这11个省市重新采集了种子。室内发芽率测定表明,同一地点两个年份采集的种子的发芽率没有差异,均在44.00%以上。因此选用2019年采集的样品分析不同纬度种群种子发芽率的差异。所有采集的种子带回实验室自然干燥后均测定千粒重,重复4次[16]。同时,还获得了原产地美国的种子作为供试样品。

表1 不同长芒苋种群的样品采集信息Table 1 Sampled information of Amaranthus palmeri populations
1.2 纸皿法和盆栽法测定不同纬度长芒苋种群的种子发芽率
采用纸皿法在人工气候箱完全控制的最优环境下测定种子的发芽率。在直径为9 cm的培养皿中铺两层滤纸,每个培养皿中随机均匀放置50粒种子,用移液枪在每个培养皿中加入等量的水,保证水分条件一致。每个长芒苋种群设置3个重复。将培养皿置于25℃、L∥D=12 h∥12 h、RH 70%的恒温人工气候箱中[15]。从第1天起每天统计发芽数,直至连续3 d不再发芽停止计数,计算每个培养皿中的种子发芽率[17]。
采用盆栽法在温室控制条件下测定发芽率,目的是模拟野外适宜环境下种子在土壤中的发芽率。每个育苗盘中放入营养土并种植50粒种子,播种深度0.5 cm,保持浇水量一致。每个长芒苋种群设置3个重复。温室的温湿度和光照条件与纸皿法保持一致。每天记录种子发芽数,连续3 d不再发芽停止计数,最终确定每个育苗盘中的种子发芽率。该试验在中国农业科学院植物保护研究所廊坊科研中试基地开展。
1.3 交互种植条件下不同纬度长芒苋种群的种子发芽率测定
为了进一步分析不同纬度长芒苋种群的种子发芽率差异并判断其是否会在入侵地发生本地适应,从采集的11个种群中选择了4个具纬度代表性的种群,包括低纬度的浙江宁波种群(ZJ)、中低纬度的河南安阳种群(HN)、中高纬度的河北石家庄种群(HB)和高纬度的吉林长春种群(JL)及美国种群(US)(表1),连续1年分别在吉林省(高纬度)、河北省(中高纬度)、河南省(中低纬度)和广东省(低纬度)4个纬度环境下多次多地交互种植,检测纬度对种子发芽率的影响。
为了确定长芒苋种子在不同纬度地区的入侵窗口期,明确其在野外的发育起点温度,自2019年4月起,每个月将这4个代表种群分别在 4个纬度地区的室外环境下各开展1次种子发芽测定的盆栽试验。每个月在每个种植地点,将长芒苋种群的种子播种到盛有营养土的育苗盘中,每个育苗盘种植50粒种子,种植深度0.5 cm,每个种群设置3个重复。不定时浇水保持水分充足。同时记录每日最低温度和最高温度。以月为单位持续观测不同种群在每个纬度种植点的种子发芽率,直至种子不再萌发为止。记录长芒苋种子能发芽的起始月份和能发芽且能完成生活史的终止月份。将种子能发芽的起始月份到发芽且能够完成生活史的月份这段时间段定义为长芒苋在该地区的入侵窗口期。
1.4 数据分析
计算不同长芒苋种群的种子在不同种植条件下发芽率的平均值和标准差,通过HSD(Tukey’s honestly significant difference)法进行显著性检验和多重比较,确定95%置信区间的种群间发芽率的差异[18]。通过单因素方差分析比较不同检测方法和不同种植地点所测定的种子发芽率之间是否具有显著性差异。对种子千粒重和纬度与种子发芽率分别进行Pearson相关性分析[19]。基于发芽率的差异对种群、种植纬度和种植时间对发芽率的影响进行三因素方差分析,确定种群、种植纬度和种植时间及它们之间的互作对种子发芽率的影响是否显著。所有数据统计分析和作图在Excel 2010和R 3.5.1软件中完成。
2 结果与分析
2.1 同质园条件下不同长芒苋种群的种子发芽率
在同质园条件下,通过纸皿法和盆栽法测定的同一种群长芒苋种子的发芽率之间的差异不显著(P=0.59)(表2)。两种检测方法均表明不同纬度种群的发芽率间存在显著差异(P<0.05)。US种群的种子发芽率低于我国采集的长芒苋种群的种子发芽率。在我国采集种群中,低纬度的福建福州种群(FJ)的种子发芽率显著低于其他种群的种子发芽率,而高纬度的吉林长春(JL)、辽宁锦州(LN)种群和中高纬度的北京(BJ)、天津(TJ)和山东滨州(SD)种群的种子平均发芽率均达到80%以上(表2)。通过纸皿法,所有长芒苋种群种子的平均发芽率为51.30%~92.00%。LN种群的种子平均发芽率最高(92.00%),US种群的平均发芽率最低(51.30%)(表2)。通过盆栽法,所有长芒苋种群种子的平均发芽率为46.20%~92.67%。BJ种群的种子平均发芽率最高(92.67%),US种群的平均发芽率最低(46.20%)(表2)。

表2 同质园条件下用纸皿法和盆栽法测定的不同长芒苋种群的种子发芽率1)Table 2 Seed germination rates of different Amaranthus palmeri populations by paper-dish and pot-planting methods in common gardens
2.2 不同纬度种群种子发芽率的影响因子分析
11个长芒苋种群的种子千粒重在0.25~0.45 g,种群间的种子千粒重差异不显著(图1)。但是Pearson相关性分析表明,通过纸皿法和盆栽法检测的不同长芒苋种群的种子发芽率与种子千粒重之间均存在线性正相关关系(图2)。
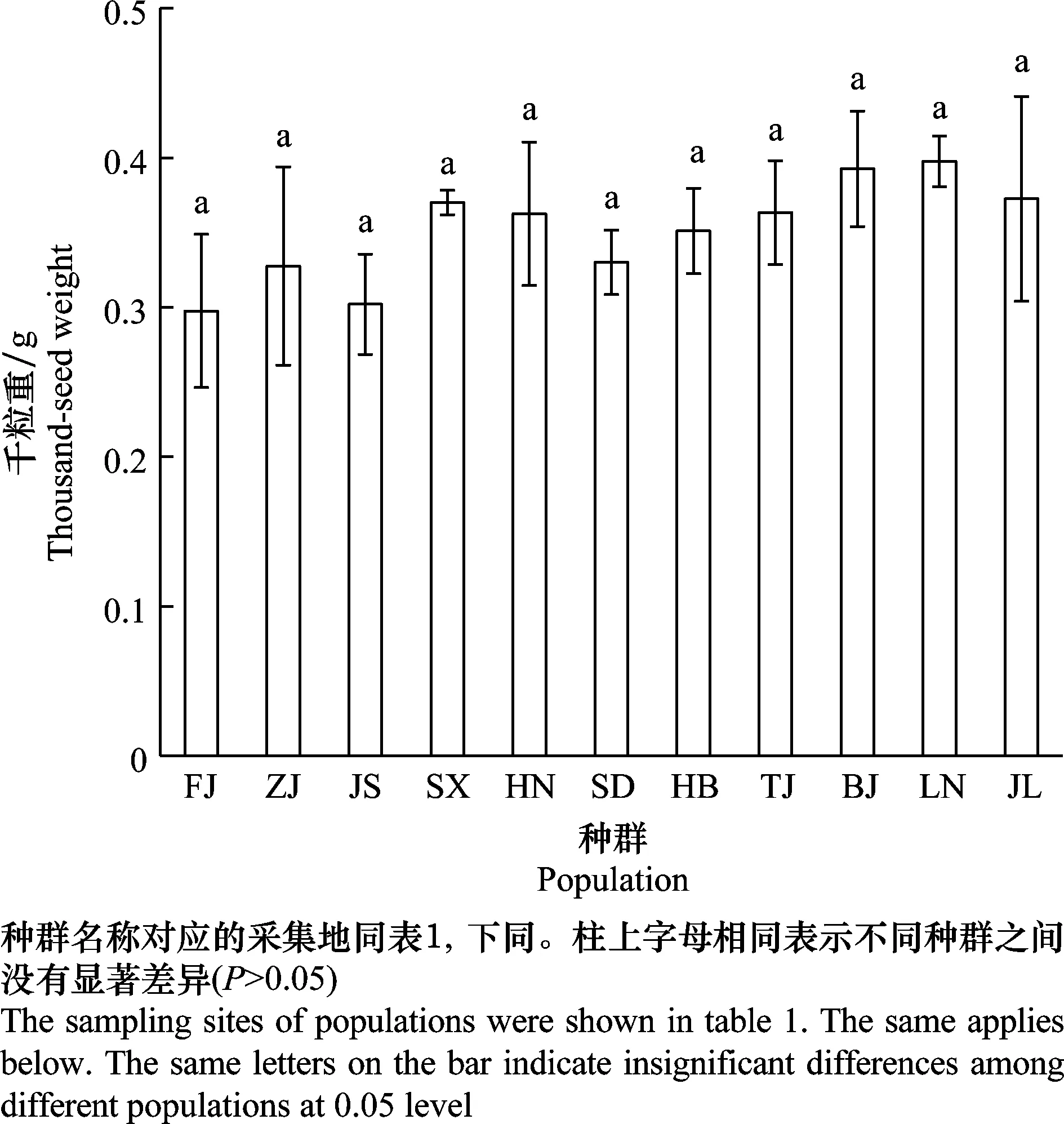
图1 不同长芒苋种群的种子千粒重Fig.1 Thousand-seed weights of different Amaranthus palmeri populations

图2 通过纸皿法和盆栽法测定的不同长芒苋种群的种子发芽率与千粒重的关系Fig.2 The relationships between seed germination rates of different Amaranthus palmeri populations and thousand-seed weights measured by paper-dish and pot-planting methods
为了确定纬度环境对种子发芽率的影响。通过Pearson相关性分析,纸皿法和盆栽法测定的不同长芒苋种群的种子发芽率与种群所处纬度之间均存在显著的线性正相关关系(图3)。高纬度种群的种子发芽率高于低纬度种群的种子发芽率(P<0.05)。
2.3 交互种植条件下长芒苋的入侵窗口期
根据对长芒苋种子发芽和完成生活史数据的统计分析,当日最低温度达到10℃,长芒苋种子开始萌发。由于长芒苋种子发芽后需要达到一定的有效积温才能完成生活史,这就导致了种群在不同纬度地区的入侵窗口期存在差异。长芒苋在高纬度中高纬度、中低纬度和低纬度的入侵窗口期分别为2个月(5月-6月)、3个月(5月-7月)、4个月(4月-7月)和5个月(4月-8月)(图4)。
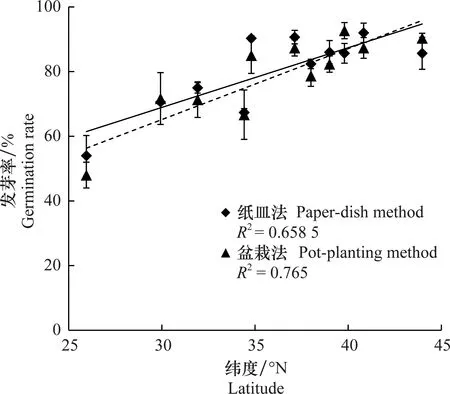
图3 通过纸皿法和盆栽法测定的不同长芒苋种群的种子发芽率与纬度的关系Fig.3 The relationships between seed germination rates of different Amaranthus palmeri populations and latitudes measured by paper-dish and pot-planting methods
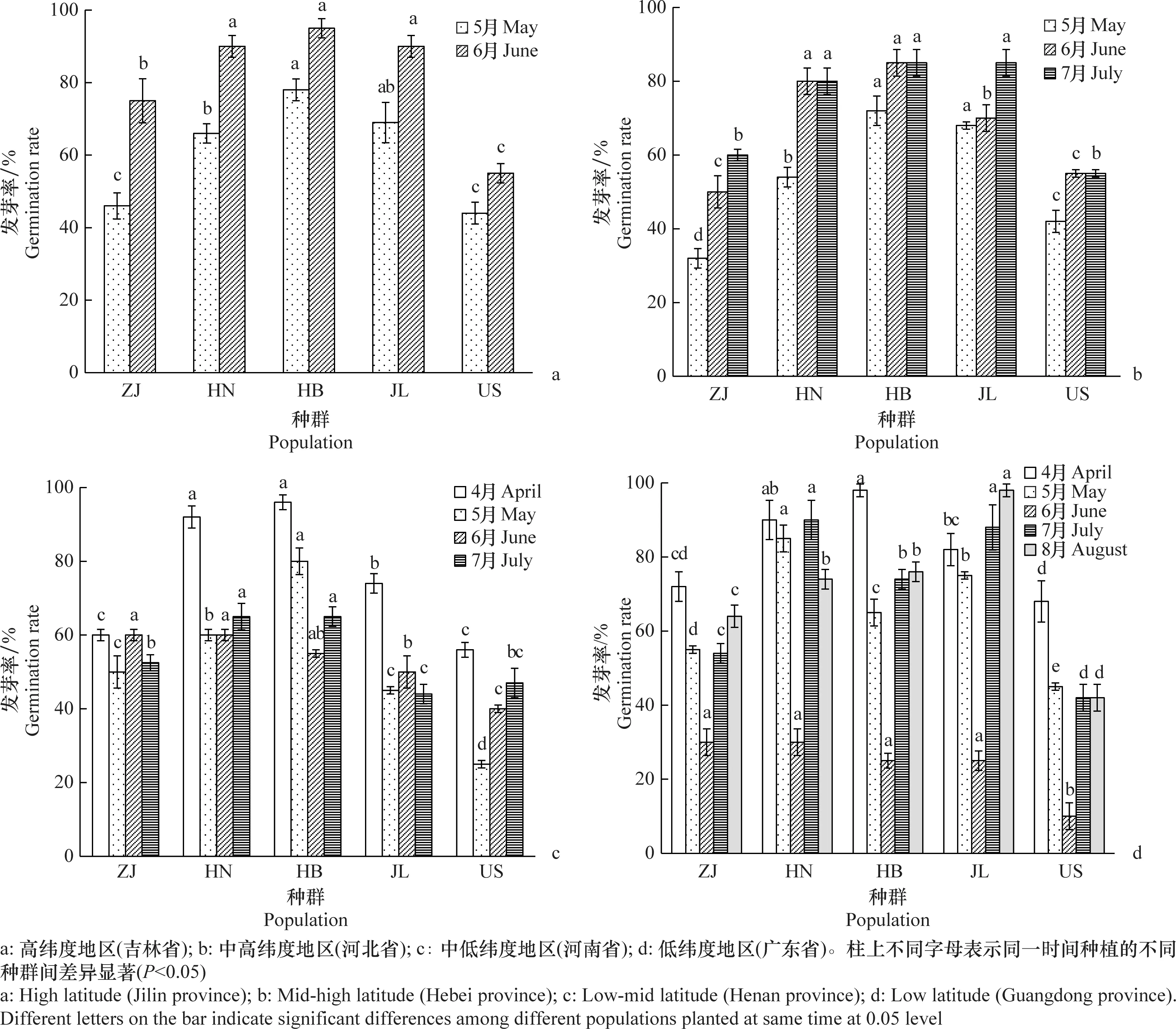
图4 不同长芒苋种群的种子在不同纬度地区种植的发芽率Fig.4 Seed germination rates of different Amaranthus palmeri populations planted at different latitudinal areas
2.4 交互种植条件下不同长芒苋种群种子发芽率比较
在入侵窗口期内,4个纬度交互种植地点的发芽率分析表明,美国种群的种子发芽率显著低于我国HN、HB和JL种群的种子发芽率(图4)。在高纬度的吉林省试验地,5个供试种群种子的平均发芽率为(70.80±14.43)%(图4a)。6月种植的种子发芽率较5月高。5月和6月的两次种植均表明,高纬度JL种群的发芽率显著高于低纬度的ZJ种群,表现出了本地适应(图4a)。在中高纬度的河北省试验地,种群的平均发芽率为(64.87±10.07)%(图4b)。5月-7月的3次种植均表明,中高纬度HB种群的发芽率高于其他种群的发芽率,且显著高于低纬度ZJ种群的发芽率,表现出了本地适应(图4b)。在中低纬度的河南省试验地,种群的平均发芽率为(58.83±11.24)%(图4c)。中低纬度HN种群4、5、7月的发芽率显著高于高纬度JL种群和低纬度ZJ种群的发芽率,表现出了本地适应(图4c)。在低纬度的广东省试验地,种群的平均发芽率为(62.28±22.29)%(图4d)。低纬度ZJ种群的发芽率低于其他种群的发芽率,没有表现出本地适应。在不同时间种植的种子的发芽率存在显著差异,6月份种子的发芽率最低,小于30%(图4d)。
种群、种植纬度和种植时间对种子发芽率的三因素方差分析结果表明,种子的发芽率受到种群、种植纬度和种植时间的共同显著性影响(P<0.01)(表3)。

表3 种群、种植纬度和种植时间对长芒苋种子发芽率影响的三因素方差分析Table 3 ANOVA analysis of three factors,population,latitude and planting time influencing seed germination rate of Amaranthus palmeri
3 结论与讨论
3.1 长芒苋在入侵地的本地适应导致了不同纬度种群的种子发芽率差异
受全球经济贸易的影响,外来植物长芒苋的种子可以随着人类活动传入不同纬度的环境中。根据本试验结果,传入我国的长芒苋种群的种子发芽率高于或者显著高于美国种群种子的发芽率(表2),表明长芒苋传入后能通过提高种子发芽率来促进种群的定殖[9]。但是,传入不同地区种群的种子发芽率存在差异,传入种群所在纬度不同程度地影响种子发芽率。同质园试验表明,长芒苋种群所在纬度与种子发芽率之间存在显著的线性正相关关系,高纬度种群的种子具有更高的发芽率,而低纬度种群的发芽率较低(表2,图3)。种子发芽率随纬度的变化表明纬度环境可能导致种群发生适应性进化,使得高纬度种群采集的种子具有更高的发芽率[20]。种群对纬度环境的快速适应可能受到表型可塑性或本地适应性进化的调控[21]。交互种植试验的结果表明,在入侵窗口期内,我国高纬度、中高纬度和中低纬度的种群都发生了本地适应(图4)。而低纬度种群的种子发芽率显著低于其他纬度的种群(6月份种植除外),且发芽率在低纬度地区也没有表现出本地适应(图4)。由于温湿度、光照等环境条件的差异,不同纬度地区适合种群生长并完成生活史的时期的长短以及在适合生长期内温湿度、光照强度不一致,可能导致不同种群后代产生的种子的发芽率存在差异。在高纬度地区,适合种群生长的时期相对较短,但是植株的生长期一直处于较为合适的温湿度环境中,适宜的环境条件有利于植株和种子更好地完成发育,种群能够产生更高的种子量且种子千粒重较大,有利于提高种子的发芽率。因此,高纬度的长芒苋种群可能通过提高种子的发芽率来增加种群在该地区的繁殖体压力和繁殖量,增强其在高纬度环境中的适应与成功入侵[22-23]。在低纬度地区,虽然适合长芒苋的生长期长,但是植株完成生活史所需的有效积温等能量积累是固定的,因此该区域持续性高温和降水导致的湿热过剩可能不利于长芒苋植株的生长发育,使得植株快速达到完成生活史所需的能量,导致最终繁殖产生的种子量较少且种子千粒重较低(图1)。千粒重越大,种子的发芽率越高(图2),种子达到一定的千粒重才能保证其有足够的营养供其繁殖生长[24-25]。因此,长芒苋种子千粒重和发芽率均随纬度环境表现出梯度变化,纬度环境和种子千粒重又导致了种子发芽率的差异。纬度和种子千粒重的互作效应导致的长芒苋在入侵窗口期内种子发芽率的差异,可能使其在不同纬度地区具有不同的入侵风险。
3.2 基于长芒苋的种子发芽率和入侵窗口期评估其入侵不同纬度地区的风险
长芒苋在不同纬度环境中存在能够发芽并完成生活史的入侵窗口期,但是不同纬度地区温湿度、光照等环境条件的差异导致各地区入侵窗口期长短不一致(图4)。长芒苋在不同纬度地区入侵窗口期的长短和种子在窗口期内发芽率的高低共同决定了长芒苋入侵该地区的风险。基于全年不同纬度多次多地的交互种植试验,长芒苋种子在高纬度地区要在5月-6月发芽才能形成入侵,在该地区的平均发芽率为(70.80±14.43)%。在中高纬度和中低纬度地区,长芒苋种子在入侵窗口期5月-7月和4月-7月的平均发芽率分别为(64.87±10.07)%和(58.83±11.24)%。在低纬度地区,长芒苋的种子在入侵窗口期4月-8月的平均发芽率为(62.28±22.29)%,在6月份的种子发芽率低于30%(图4)。通过对试验数据和纬度环境的结合分析,长芒苋在低纬度地区较低的发芽率可能是由于降水过多导致的。根据长芒苋在不同纬度地区表现出种子发芽率和入侵窗口期的梯度变化,可以对长芒苋入侵不同纬度地区的风险进行评估。中高纬度和中低纬度地区种子发芽率较高且入侵窗口期较长,导致长芒苋在中低纬度至中高纬度地区具有较高的入侵风险。高纬度地区种子发芽率高但是入侵窗口期短,低纬度地区虽然入侵窗口期长但发芽率低,因此长芒苋在高纬度和低纬度地区的入侵风险相对较低。
根据基于种子发芽率和入侵窗口期对长芒苋入侵风险的评估结果,建议在不同纬度地区开展区域化精准防控。长芒苋在中低纬度的华中地区至中高纬度的华北地区入侵风险较高,应该重点加强防控,建议对所有可能的入侵地进行监测预警,在7月底种子成熟前,对已经定殖的种群通过农业、物理、人工、化学防治等多种防治技术相结合的方式进行综合防除,阻止其种群扩张蔓延。此外,长芒苋的种子在野外成熟后可以形成土壤种子库,第一年未能发芽的种子还可以在接下来的几年中不断发芽,有利于种群的维持[26-27]。因此,对区域内已经定殖的长芒苋种群,由于其产生的种子量大,种子发芽率高,应该在每年7月底前开展一次综合防除工作,进行连续3~5年的持续防控,以达到彻底清除土壤种子库和消除种群定殖风险的目的。长芒苋在高纬度的东北地区由于入侵窗口期短,建议在6月底前集中对营养生长期的植株开展物理或者人工全面灭除,可以达到早期根除的目的。长芒苋在低纬度的华南地区种子发芽率低,但是入侵窗口期长,建议应该全年加强监测预警以防止种子的不断传入。
综上所述,不同纬度长芒苋种群的种子发芽率随纬度升高而增高,而入侵窗口期随纬度升高而缩短。根据不同纬度长芒苋种群的种子发芽率和入侵窗口期的差异,长芒苋在中低纬度至中高纬度地区入侵风险较高,应该重点防控。长芒苋在高纬度和低纬度地区的入侵风险相对较低,但是长芒苋种子发芽率的本地适应有利于增加其在高、低纬度地区的入侵风险。因此应该加强对我国已传入长芒苋种群监测与防控力度以抑制其进一步扩散蔓延。
