胡萝卜种子粒形与千粒重的QTL分析
2022-12-14王雪伟欧承刚黄建新刘星郝卫东赵志伟孔小平庄飞云
王雪伟 欧承刚 黄建新 刘星 郝卫东 赵志伟 孔小平 庄飞云*
〔1 中国农业科学院蔬菜花卉研究所,北京 100081;2 中蔬种业科技(北京)有限公司,北京 100081;3 西宁市蔬菜技术服务中心,青海西宁 810016〕
胡萝卜(DaucuscarrotL.)属于伞形科胡萝卜属两年生草本植物,富含α、β-胡萝卜素,是人类维生素A 的主要来源(方智远,2017)。胡萝卜种子为不规则小粒种子,多为长扁形,外表有细小毛刺,流动性差,种子胚小,顶土能力弱(杨亮等,2021)。胡萝卜种子生产方式主要有两种:一种是种子—种子,即种子前一年播种并露地过冬,翌年抽薹开花收种;另一种是根—种子,即种子前一年播下后收取肉质根越冬贮藏,翌年种植肉质根后抽薹开花收种(Rubatzky et al.,1999;方智远,2017)。目前胡萝卜规模化种植基地基本都采用精量化播种方式,如种绳、丸粒化等,对种子质量提出了较高的要求。除了发芽率,种子形状、大小和千粒重等都会影响胡萝卜播种及出苗的整齐度和一致性,但目前尚无胡萝卜种子粒形和千粒重的遗传研究报道。
在小麦、水稻、玉米中的研究表明种子相关性状为多基因控制的数量性状,已鉴定到多个与之相关的主效QTL 位点。Feng 等(2021)采用F2和F2∶3群体,对水稻粒长、粒宽、粒长宽比、粒厚和千粒重5 个粒形性状进行QTL 定位,共鉴定出分布在1、2、3、6、7、8 号染色体上的12 个相关QTL,在7 号染色体上还存在1 个F2和F2∶3群体重叠区间,表型贡献率在15.09%~16.30%之间;Kumari 等(2018)选择了1 个含有106 个株系的RIL 群体对小麦粒长、粒宽、粒长宽比、千粒重和粒周长进行QTL 定位,发现了13 个分布在1A、2B、4A、5A、6A、7A、7D 染色体上的QTL,表型贡献率在7.66%~63.66%之间;Wang 等(2020)通过不同环境下培育F2及其F2∶3群体,探寻控制玉米粒长(KL)、粒宽(KW)的稳定QTL,最终在3 种不同环境中定位到2 个稳定QTL(qKW-1和qKL-2)。由于粒形不仅影响种子产量,也影响出苗和发育,多项对甘蓝型油菜种子粒形和粒重的QTL 定位研究纷纷展开。Quijada 等(2006)利用种植在两个不同环境中的DH 群体定位到3 个与粒重相关QTL 位点;Zhao 等(2016)对8 个不同环境中种植的DH 群体进行粒形和粒重的QTL 定位,鉴定到25 个相关QTL,并且在A7 和C1 染色体上均存在重叠区间,表型贡献率均大于10%。随着多个QTL 位点被发现,已成功克隆与粒形性状相关的基因逾100 个,如GW2(控制粒宽和千粒重)(Song et al.,2007)、GS3(控制粒长)(Fan et al.,2006)、BSG1(控制粒长和粒厚)(Yan et al.,2013)、GL4(控制粒长)(Wu et al.,2017)和GW5(控制粒宽)(Liu et al.,2017)等。通过对基因结构和分子特性研究发现这些基因主要通过泛素-蛋白酶体途径、IKU 途径、G-蛋白信号途径、MAPK 途径、植物激素途径、转录调控因子途径或者其他途径来调控种子粒形和千粒重(Li &Li,2016)。本试验基于前期构建的回交重组自交系(backcross inbred line,BIL),对胡萝卜种子粒长、粒宽、长宽比和千粒重进行遗传分析,基于胡萝卜全基因组测序构建的高密度遗传图谱进行相关性状QTL 定位,旨在为最终克隆粒形和千粒重的遗传调控基因奠定基础,也为胡萝卜种子性状的改良提供理论依据。
1 材料与方法
1.1 试验材料
中国农业科学院蔬菜花卉研究所胡萝卜课题组前期以亲缘关系较远的白色胡萝卜品种松滋野生(Ws)和橘色栽培品种Amsterdam(Af)为亲本进行杂交,以Af 为轮回亲本,经2 代回交,再单株自交6 代,得到了包含200 个株系的BIL 群体(BC2S6)。2019 年8 月上旬在中国农业科学院蔬菜花卉研究所南口农场进行露地播种,11 月初采收,每个BIL 株系随机选取20 个肉质根越冬贮藏,2020 年3 月种植春化后的肉质根,5 月进行自交授粉,7 月收获各株系种子进行后续统计分析。
1.2 性状测量与数据分析
从BIL 群体中随机选取110 个株系,每个株系随机选取30 粒种子,使用ImageJ 软件进行粒长和粒宽的测定,并计算长宽比,3 次重复,取平均值作为该性状表型值;每个株系随机选取1 000 粒种子,使用万分之一天平称重,3 次重复,取平均值作为千粒重。使用SPSS 22 软件进行数据和相关性分析。
1.3 遗传图谱构建及QTL 定位
在中国农业科学院蔬菜花卉研究所胡萝卜课题组前期构建BIL 群体的遗传图谱基础上(Ou et al.,2022),利 用Windows QTL Cartographer(WinQTL)v2.5 软件的复合区间作图法(CIM)(Wang et al.,2011),95%置信区间在全基因组范围内检测种子粒长、粒宽、长宽比和千粒重的QTL,QTL 命名遵照McCouch(2008)建议的规则命名。
2 结果与分析
2.1 胡萝卜种子粒形与千粒重的表型分析
从表1 可以看出,亲本Ws 种子粒长为2.37 mm,粒宽为1.35 mm,长宽比为1.76,千粒重为1.10 g;Af 粒长为2.67 mm,粒宽为1.43 mm,长宽比为1.87,千粒重为1.61 g;两亲本种子粒长、粒宽、长宽比和千粒重差值分别为0.30 mm、0.08 mm、0.11 和0.51 g。BIL 群体种子粒长平均值3.05 mm,变异范围为2.09~4.26 mm;粒宽平均值1.54 mm,变异范围为0.97~2.20 mm;长宽比平均值1.99,变异范围为1.30~2.64;千粒重平均值1.96 g,变异范围为0.68~3.63 g。

表1 胡萝卜亲本、BIL 群体种子粒形性状表型分布
从BIL 群体种子粒长、粒宽、长宽比和千粒重的频次分布结果可知(图1),这4 个与胡萝卜种子粒形相关的性状均呈偏正态分布,可进行QTL定位以及作图。

图1 胡萝卜BIL 群体种子粒长、粒宽、长宽比、千粒重频次分布
2.2 胡萝卜种子粒形与千粒重的相关性分析
对胡萝卜BIL 群体种子粒长、粒宽、长宽比和千粒重等4 个性状进一步进行相关性分析。结果表明(表2),千粒重与粒长和粒宽均呈极显著正相关,相关系数分别为0.661 和0.715;粒长与粒宽亦呈极显著正相关,相关系数为0.659;长宽比与粒长呈极显著正相关,与粒宽呈极显著负相关,但与千粒重不相关。

表2 胡萝卜BIL 群体种子粒形性状的相关性分析结果
2.3 胡萝卜种子粒形与千粒重的QTL 分析
对胡萝卜BIL 群体种子粒长、粒宽、长宽比和千粒重进行QTL 分析,共获得6 个QTL 位点(表3)。3 个粒长QTL 分别位于8、9 号染色体上,LOD 值分别为2.61、2.72 和2.89,表型贡献率分别为12.0%、9.7%和9.6%,命名为qGL8、qGL9-1 和qGL9-2;粒宽QTL 有1 个,位于8 号染色体上,LOD 值为2.68,表型贡献率为11.0%,命名为qGW8;千粒重QTL 有2 个,分别位于1、9 号染色体上,LOD 值分别为5.54 和4.02,表型贡献率分别为18.3%和8.8%,命名为qTGW1 和qTGW9;没有得到与长宽比相关的QTL 位点。
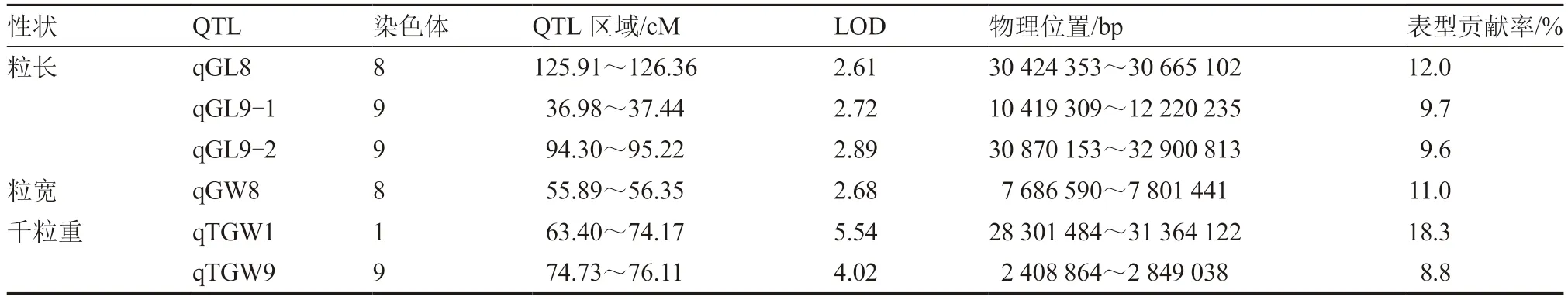
表3 胡萝卜BIL 群体种子粒形相关性状的QTL 定位
2.4 候选基因分析
通过定位共得到6 个QTL 位点,基于胡萝卜参考基因组注释信息(Iorizzo et al.,2016),共在显著候选区间内检索到399 个具有功能注释的基因,其中10 个可能与种子粒长、粒宽和千粒重相关(表4)。qGL8 位点鉴定到2 个可能参与调控粒长的基因,LOC108199762是编码富含丝氨酸/苏氨酸蛋白激酶的基因,作为脱落酸(ABA)信号传导的负调节剂,通过调节ABA 依赖性基因的表达来调控种子长短(Pernas et al.,2007);LOC108198581主要通过引导花粉管的生长来影响种子的大小(Wang et al.,2016)。qGW8 位点鉴定到1 个可能参与调控粒宽的基因,LOC108199012是AP2 基因家族转录因子,AP2 控制胚乳的发育,拟南芥中ap2突变体种子比野生种子更大(Ohto et al.,2009)。qTGW1位点鉴定到7个可能参与调控千粒重的基因,LOC108203830和LOC108226269主要编码E3 泛素连接酶,通过作用于不同受体蛋白调节细胞死亡和防御机制来控制种子大小(Marino et al.,2013;Xia et al.,2013);LOC108226660主要编码G-蛋白,作为调节剂或转导剂参与各种跨膜信号系统,通过促进细胞增殖来调节种子大小(Mudgil et al.,2009);LOC108212357和LOC108226394与ARF 有关,ARF 是一种多效性发育调节剂,通过限制种子珠被中的细胞增殖来控制种子大小(Schruff et al.,2006);LOC108205141编码富含丝氨酸/苏氨酸的蛋白激酶,负调控ABA(Pernas et al.,2007)。LOC108208217编码MYB 转录因子,通过调节雄蕊的发育以响应茉莉酸信号传导,负调节ABA(Cui et al.,2013)。

表4 胡萝卜种子粒形相关性状QTL 位点显著区间内相关预测基因
3 讨论与结论
出苗率高、一致性好是胡萝卜获得高产和优良商品性的前提和基础。为节省劳动力和种子成本,目前规模化胡萝卜种植基地基本都采用精量化播种方式,例如使用种绳或进行丸粒化等,这便对胡萝卜种子的发芽率和萌发一致性提出了更高的要求。因此,明确胡萝卜种子相关性状的遗传调控机理,不仅是对种子性状进行遗传改良的理论基础,更对建立规范化的胡萝卜种子繁育与加工体系具有十分重要的意义。本试验中,亲缘关系较远的2 个亲本松滋野生(Ws)和Amsterdam(Af)种子粒长和粒宽差异较小,但是千粒重存在较大差异。BIL 群体在种子粒长、粒宽、长宽比和千粒重中均表现出较大分离,特别是粒长和千粒重变异较大,分别为2.09~4.26 mm 和0.68~3.63 g;相关性分析表明粒长、粒宽与千粒重两两之间均呈极显著正相关,长宽比与粒长呈极显著正相关、与粒宽呈极显著负相关,说明粒长和粒宽均能够影响千粒重,这与在小麦和水稻中的研究结果相似(Kumari et al.,2018;Xue et al.,2019)。因此,在对胡萝卜种子性状进行改良时,也要注意粒长、粒宽与千粒重之间要协同改良。
种子粒形相关性状均为数量性状,受多基因控制(Xing &Zhang,2010;Huang et al.,2013),目前在胡萝卜种子粒形与千粒重的QTL 分析方面几乎空白。申琪等(2018)利用与本试验相同的遗传图谱,对胡萝卜BIL 群体的始花期、主薹高和单株种子量进行QTL 分析,分别得到2、4、2 个相关QTL 位点,表型贡献率在14.6%~23.8%之间。本试验继续对该群体的种子粒长、粒宽和千粒重进行QTL 分析,获得了3、1、2 个相关QTL 位点,表型贡献率在8.8%~18.3%之间,每个性状之间没有位点重叠,说明胡萝卜种子粒形和千粒重性状是独立遗传的。这是首次对胡萝卜种子粒形和千粒重进行遗传分析和QTL 定位,不仅为后续进行精细定位和挖掘功能基因奠定了理论基础,还可以更加有效的借助分子标记技术对胡萝卜种子粒形和千粒重进行改良,改善种子质量。
Li 和Li(2016)将已克隆出来的与种子性状相关基因划分为6 种途径:IKU 途径、泛素-蛋白酶体途径、G-蛋白信号途径、MAPK 途径、植物激素途径和转录调控因子途径,包括控制水稻粒宽和千粒重的基因GW2(Song et al.,2007)、控制水稻粒长的基因GS3(Fan et al.,2006)、控制水稻粒长和粒厚的基因BSG1(Yan et al.,2013)、控制玉米千粒重的基因GRMZM2G039934(Chen et al.,2013)、控制小麦粒宽和千粒重的基因TaDA1(Liu et al.,2020)等。本试验通过对显著区间进行候选基因筛选,检索到参与其中4 种调控途径上的9 个基因和其他因素引起的1 个基因。泛素化在影响种子大小方面起着重要的作用(Song et al.,2007;申琪 等,2018)。在水稻中,已经被克隆的GW5、GW2均是编码E3 泛素连接酶的基因,通过正向和反向调节种子粒宽和粒重来调控种子大小(Xia et al.,2013;Choi et al.,2018)。本试验鉴定到的LOC108203830和LOC108226269为主要编码E3 泛素连接酶的基因,推测可能与种子千粒重有关。LOC108226660是一个编码G-蛋白β 亚基的基因,G-蛋白信号是一个保守的分子信号,参与多个信号调控通路,包括G-蛋白α 亚基、β 亚基和γ 亚基(Mudgil et al.,2009)。在拟南芥中,α 亚基和β 亚基参与花发育的调控,γ 亚基控制花芽、角果的发育及籽粒大小(Chakravorty et al.,2011;Li et al.,2012)。在水稻中发现β 亚基表达量降低也会引起籽粒变小(Urano et al.,2016)。因此LOC108226660可能与胡萝卜种子大小有关。LOC108199762和LOC108205141编码富含丝氨酸/苏氨酸的蛋白激酶,是ABA 信号传导的负调节剂,但LOC108199762通过调节ABA 依赖性基因的表达来控制种子粒长,而LOC108205141通过负调控ABA,影响花粉发育来控制种子千粒 重(Pernas et al.,2007);LOC108212357和LOC108226394与多效性发育调节剂ARF 有关,通过限制种子珠被中的细胞增殖来控制种子大小(Schruff et al.,2006)。ABA 是通过影响细胞增殖和扩张来调节籽粒大小的重要因素(Krishnan &Dayanandan,2003)。LOC108199012为AP2基因家族转录因子,AP2 控制胚乳的发育,在拟南芥中ap2突变体的种子比野生种子更大(Ohto et al.,2009);LOC108208217编码MYB 转录因子,通过调节雄蕊的发育以响应茉莉酸信号传导,负调节ABA(Cui et al.,2013)。转录因子也被称作基因表达调控因子,不仅参与植物细胞中的反应,在控制种子大小方面也有一定影响(Xiao et al.,2006;Chae et al.,2008;Ramamoorthy et al.,2008;Sun et al.,2010)。此外,本试验还得到1 个主要通过引导花粉管的生长来影响种子大小的基因LOC108198581。这些基因通过不同途径对胡萝卜种子粒形和千粒重起着不同程度的作用,是后续基因功能研究的候选基因。
