基于三色堇全长转录组的MYB基因家族鉴定
2022-12-13王童欣潘庆龙李霆格
谷 佳, 王童欣, 饶 英, 潘庆龙, 李霆格, 赵 莹, 周 扬, 彭 婷, 王 健
(1.热带特色林木花卉遗传与种质创新教育部重点实验室/海南省热带特色花木资源生物学重点实验室/海南大学林学院,海南海口 570228;2.贵州大学农学院,贵州贵阳 550025)
植物在生长发育过程中受到多种转录因子调控,如WRKY、MYB、AP2/ERF转录因子等[1],其中MYB家族具有特定的保守结构域,根据其结构域重复的次数可分为4R-MYB、3R-MYB、1R-MYB、R2R3-MYB 等4类,其中R2R3-MYB 转录因子是MYB家族中最重要的、目前研究最广泛的一类转录因子,在植物体内广泛存在[2]。MYB转录因子对植物具有重要影响,可以参与植物根、茎、叶和花的生长[3],也可调控植物代谢,如调节类黄酮、花青素等物质合成[4]。目前,通过基因组以及转录组数据对物种进行MYB基因家族进行鉴定,从而对物种生长发育进一步进行研究探索,已经在拟南芥(Arabidopsisthaliana)、彩叶杨(Populusdeltoids)、甘蔗(Saccharumsp.)等物种进行MYB基因家族研究,如拟南芥中发现含有126个R2R3-MYB、64个1R-MYB、5个3R-MYB、1个4R-MYB[2],在彩叶杨中已被鉴定出183 个R2R3-MYB、 5个3R-MYB、1个4R-MYB和 113个1R-MYB[5],在甘蔗中鉴定出57个R2R3-MYB、13个1R-MYB[6],进行基因家族的鉴定已经成为MYB功能挖掘的重要研究手段。
三色堇(Viola×wittrikiananGams.)为堇菜科堇菜属植物,是常见的花卉植物,因其花斑形状多样,颜色多彩而具有较高的观赏价值和经济价值,国内外应用十分广泛。目前,对三色堇MYB基因家族分析未见报道,其在三色堇中基因功能尚不清楚。本研究通过对三色堇3代全长转录组测序,在三色堇3代全长转录组数据的基础上鉴定出三色堇MYB转录因子并对其序列特征、蛋白质保守基序、进化关系等进行分析,对挖掘三色堇中具有特殊功能的MYB基因具有重要的意义。
1 材料与方法
1.1 材料与序列来源
材料使用花瓣为黄底紫斑的三色堇(Viola×wittrockianaGams.)梦蝶,2020年1月种植于海南大学林学院基地,播种3个月后采摘三色堇植株的根、茎、叶、花并均匀混合,置于液氮中速冻,送广州基迪奥生物科技有限公司利用单分子实时测序技术(SMRT)进行三色堇3代全长转录组测序。蛋白序列源于本测序上传的所有转录组数据库,拟南芥MYB家族氨基酸序列下载于Plant TFDB 3.0。
1.2 三色堇MYB转录因子鉴定与序列特征分析
从Pfam 31.0数据库下载MYB结构域种子文件(PF00249),之后利用HMMER 3.2.1 软件创立Profile HMM(数值表格型隐马可夫模型)[7],检索三色堇转录数据,对检索结果去冗余,得到候选VwMYB蛋白序列。候选蛋白序列用SMART软件预测[8],再通过NCBI的Blastp分析,去除重复序列,得到三色堇MYB序列蛋白。利用在线工具 ExPAS软件对三色堇 MYB转录因子家族成员的蛋白分子量(pW)、蛋白长度以及等电点(pI)等基本属性进行分析[9];利用CELLO v.2.5软件进行亚细胞定位预测[10]。
1.3 三色堇MYB家族保守结构域分析
使用软件MEGA-X中的ClustalW 对三色堇R2R3-MYB进行多序列比对,再利用 WebLogo软件获取序列标识。利用在线网站MEME和TBtools软件分析三色堇MYB转录因子蛋白保守基序序列[11]。
1.4 三色堇R2R3-MYB转录因子系统发育分析
使用 MEGA-X软件,利用邻域连接(NJ)法,设置bootstrap(n=1 000)进行系统发育分析[12]。
2 结果与分析
2.1 三色堇MYB转录因子基因鉴定与分类
以MYB结构域序列作为靶序列,利用三色堇全长转录组数据进行筛选,得到141个MYB 转录因子,后通过SMART与Blastp 2次筛选最终得到121个MYB,分别编号VwMYB1~VwMYB121,其中共包含61个1R-MYB、57个R2R3-MYB以及3个 3R-MYB 类转录因子。
2.2 三色堇MYB类转录因子基本信息分析
对三色堇MYB转录因子家族成员的氨基酸序列、分子量、等电点进行分析,结果表明,三色堇MYB基因家族中,序列最长的为VwMYB61(Isoform0000184),有1 996个氨基酸残基,最短的为VwMYB91(Isoform0026081),有119个氨基酸残基;等电点从10.16 (VwMYB43,Isoform0045501)到4.44 (VwMYB75,Isoform0008562);分子量从13.9 ku(VwMYB91,Isoform0026081)到216.3 ku (VwMYB61,Isoform0000184)。蛋白亚细胞定位预测显示,多数MYB定位到细胞核,还有部分定位到细胞质、线粒体等(表1)。
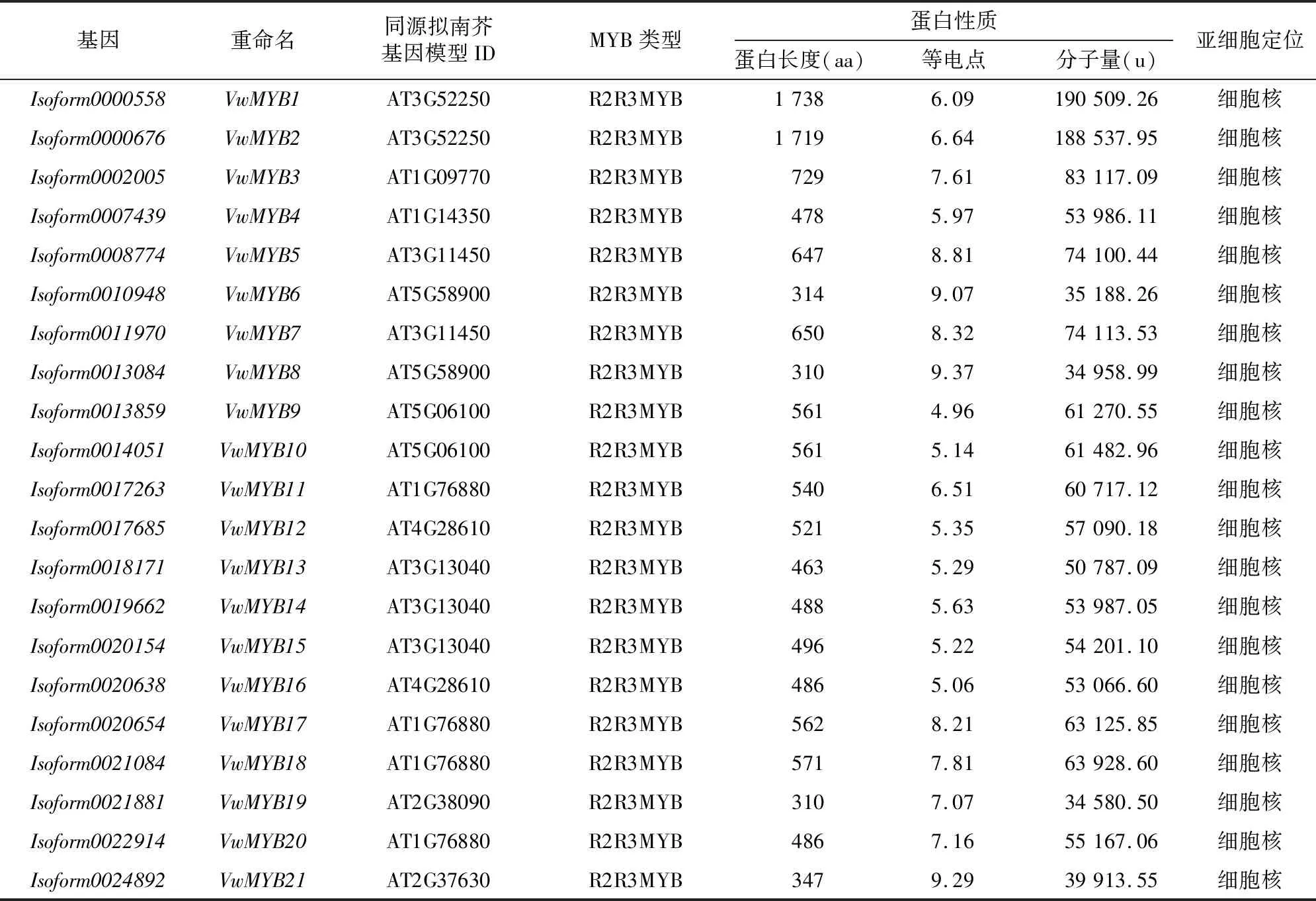
表1 三色堇MYB转录因子基本信息

表1(续)

表1(续)

表1(续)
2.3 三色堇MYB保守基序分析
对121个三色堇MYB转录因子进行保守基序分析,获得 20种保守motif元件(图1),其中分布频次最高的是motif2(15.2%),其次是motif4 (12.6%),最低的是motif20 (0.3%)(图2)。不同MYB蛋白的motif数量与分布存在差异,多数motif 元件分布在 N 端,少数(如 motif9)分布在MYB的C端,同一个亚组的基因具有相似基序,根据亲缘性说明其具有相似功能,同时序列所含motif数量、类型与分布存在不同,表明不同基因功能之间具有差异,也存在一些高频率基序,如motif 2几乎存在于每一个亚组中,反映这个基序对VwMYB蛋白功能具有重要性。
R2和R3的保守结构域是识别MYB序列所必需的,通过WebLogo对三色堇的57个R2R3MYB序列位点信息进行分析,从图3中可看出,三色堇R2R3-MYB类转录因子结构域存在R2R3MYB特有的W型保守氨基酸,R2 MYB与R3 MYB的结构域均含有52个氨基酸残基,R2 MYB包含3个高度保守的色氨酸残基(W)(图3-A),R3 MYB第1个色氨酸(第925位)被亮氨酸(L)所取代(图3-B),只有2个色氨酸残基(W)。三色堇保守结构域特点与已报道拟南芥、辣椒、梨等物种一致[13]。
2.4 三色堇R2R3型MYB家族进化分析
将三色堇R2R3-MYB和拟南芥MYB蛋白家族进行系统发育分析,根据樊锦涛等对拟南芥R2R3-MYB族群的划分依据,对三色堇R2R3-MYB类转录因子划分亚群,结果见图4[14]。三色堇R2R3-MYB类转录因子可划分为30个亚群(A1~A30),有8个亚群不能与拟南芥聚类,其他22个亚群可以聚类到拟南芥S1、S4、S6、S7、S9、S16、S18、S22、S23亚群中。聚类在同一个亚群的基因具有较高的同源性与序列相似性,并可能具有类似的功能[15]。
3 讨论与结论
三色堇没有参考基因组,这限制了对三色堇的深入研究,但利用3代测序技术可以快速得到全长转录组,获得植物大部分有效的基因表达信息,是对非模式植物研究的新方法。本研究利用3代测序得到三色堇全长转录本,分析得到121个MYB转录因子,接近已报道玉米基因组分析发现的132个ZmMYB基因[16],明显高于亚洲百合基于2代转录组测序结果发现的69个LaMYB基因[17],表明3代测序获得高质量的cDNA全长序列与基因信息,为三色堇MYB转录因子研究提供有效参考。
MYB转录因子N端包含保守DNA 结合域,该结合域由1~4个不完全重复序列(R)构成,每个重复序列约52个氨基酸,构成3个α-螺旋[18],其中第2和第3螺旋(R2和R3)形成 HLH(Helix-Loop-Helix)结构与目标基因结合[19]。本研究获得的三色堇MYB转录因子全部包含上述的R序列,共有61个1R-MYB、57个R2R3-MYB、3个3R-MYB类转录因子,符合MYB转录因子基因家族的基本结构特征。在大部分MYB转录因子中,N端保守性更高,C端序列具有可变性,负责蛋白质活性调节,MYB转录因子具有通过其保守的N端结构域转录调节C端结构域参与复杂的生理过程[20],这与本研究中三色堇MYB转录因子的大部分motif 元件集中、规律地分布在N端,少数无规律地分布在 MYB 的C端的保守基序结构特点一致,表明MYB基因家族可以通过其特殊的保守结构域进行识别以及对基因功能产生影响。
R2R3-MYB是目前研究最广的转录因子,参与植株生长发育、代谢调节、表皮细胞形态分化等功能调控[21]。本研究中将三色堇57个R2R3-MYB与拟南芥共同进行系统发育进化分析,其中一些三色堇转录因子与拟南芥处于同一分支, 表明这2个物种的MYB基因家族具有部分相同的进化过程。一般来说,聚集在同一个分支的基因亲缘关系相近,其在功能上也具有相似性,如居利香等利用该方法认证多个与辣椒相关的转录因子,其中CaMYB141、CaMYB71与拟南芥亚组20中的AtMYB2聚为一类,认为CaMYB141、CaMYB71与AtMYB2功能相似,均可在干旱条件下增加辣椒素含量[22]。本研究中,三色堇中的VwMYB48、VwMYB53、VwMYB47、VwMYB52与拟南芥亚组4中的AtMYB3、AtMYB4、AtMYB7、AtMYB32聚为一类,表明三色堇中VwMYB48、VwMYB53、VwMYB47、VwMYB52可能参与植物生长过程中的生物协迫与非生物胁迫调节过程[23]。VwMYB40、VwMYB35与拟南芥亚组1中的AtMYB30、AtMYB96关系最近,AtMYB30、AtMYB96与激素调节有关,且主要关于脱落酸与水杨酸,而脱落酸影响花青素的合成[24],推测三色堇中VwMYB40、VwMYB35通过调节激素的合成从而影响花青素的合成。同理,三色堇中VwMYB39、VwMYB36、VwMYB43、VwMYB41与拟南芥第七亚组中的AtMYB11、AtMYB12和AtMYB111聚为一类,已有研究表明AtMYB11、AtMYB12、AtMYB111与黄酮醇生物合成相关[25],因此VwMYB39、VwMYB36、VwMYB43、VwMYB41也可能参与三色堇中黄酮醇生物合成,调节植物代谢活动。VwMYB51、VwMYB54与拟南芥亚组6中的AtMYB75、AtMYB90、AtMYB113、AtMYB114最相近,而AtMYB75、AtMYB90、AtMYB113、AtMYB114是控制植物花青素合成的基因[26],表明三色堇的花色形成可能受到VwMYB51、VwMYB54调节,这2个转录因子可能参与三色堇花青素合成[27]。但是,有一部分三色堇MYB与拟南芥分支不同,如A1分支不含有任何AtMYB,这种转录因子的特异性可能是物种后期进化分离导致[28]。当然,上述论述只是基于基因结构相似性的推测,是否具有相应的功能及其作用机制,还需要进一步研究阐明。




