基于GAM 模型的浙江近海曼氏无针乌贼时空分布研究
2022-12-12徐晓萱刘姝含周永东刘连为隋宥珍张洪亮徐开达孙浩奇
徐晓萱,谢 玉,刘姝含,周永东,刘连为,隋宥珍,张洪亮,徐开达,孙浩奇
(浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,农村农业部重点渔场渔业资源科学观测试验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021)
曼氏无针乌贼Sepiella maindroni属软体动物门Mollusca、头足纲Cephalopoda、乌贼目Sepiida、乌贼科Sepiidae、无针乌贼属Sepiella,俗称墨鱼、乌贼等,广泛分布于印度东海岸至西太平洋,在我国主要分布于浙江近海与闽东海域,历史上黄海、南海有少量集群[1-2]。浙江省曼氏无针乌贼渔获量曾占我国海洋捕捞渔获量的前列,历史最高年产量达6 万多t,占浙江省海洋捕捞总产量的9.3%[3],并成为东海渔业资源开发的重要研究对象。
该种为典型的短距离洄游性生物,从4 月中旬开始从东海近海向沿岸作生殖洄游,进入渔山、韭山、中街山渔场和嵊泗渔场等,随后深入岛礁周围进行产卵,11 月份作越冬洄游,开始返回越冬场[4-5]。国内外关于曼氏无针乌贼的研究主要涉及其繁殖生物学[6-10]、群体遗传学[11-12]、摄食生态学[13]等方面,但是关于环境因子变化对曼氏无针乌贼时空分布影响机理的研究偏少,且缺乏大范围和长时间的调查研究。由于GAM模型可直接处理响应变量与多个解释变量之间的非线性关系,对不同形式的函数进行加和,找出其中的规律,适用于不同类型分布的函数分析[14-17]。因此,本研究根据2014-2021 年连续年份的底拖网数据结合CTD 获得的环境数据,采用GAM 模型分析影响其资源分布的主要环境因子,探讨浙江近海曼氏无针乌贼的时空分布随环境变化的响应,以期为曼氏无针乌贼渔业资源评估和管理提供数据基础。
1 材料与方法
1.1 数据来源
采用的渔业数据来源于2014-2021 年4 月和11月浙江省海洋水产研究所在浙江近海进行的底拖网渔业资源调查,调查范围为27°N-31°N、120.5°E-124°E,其调查位置分布如图1 所示。数据内容包括作业时间、经度、纬度、渔获种类和生物量等信息。调查网具网口拉紧周长为25 m,囊网网目尺寸为3.0 cm,拖速为3 kn,每个站位拖网时间为1 h,每一个调查站位的水深、水温、盐度等环境数据通过CTD 同步测定。依据《海洋渔业资源调查规范》[18]采集、保存样品,在实验室对采集的曼氏无针乌贼进行生物学测定。

图1 调查站位示意图Fig.1 Sketch of survey station
1.2 数据处理
1.2.1 生物量密度估算
曼氏无针乌贼的生物量密度估算公式[19]如下:

式中:D为现存生物量密度,kg·km-2;C为每小时取样面积内的渔获量,kg;q为网具渔获率,由于曼氏无针乌贼属于沿岸暖水底栖种,q取0.5;A为网具每小时扫海面积,km2。
1.2.2 模型分析方法
建立模型之前,先对各影响因子进行Pearson 相关性分析[16],将2 个具有显著相关性的因子保留其中1 个,分析后初步选择年份、月份、纬度、水深、底层水温、表层盐度和底层盐度这几个因子。利用GAM 模型对曼氏无针乌贼资源密度与时空分布及环境因子进行回归分析。其一般表达式为[20]:

式中:函数Y 为连接函数;α 为常数截距项;x 为解释变量;s(x)为连接解释变量的样条平滑函数(spline smoothing);ε 为随机残差项。
将所选择的因子带入模型,用AIC 准则(akaike information criterion)检验模型的拟合程度,以AIC 最小值的模型作为最优模型[21]。将GAM 模型中上下95%的置信区间虚实线最接近的部分定义为最适环境范围[22]。GAM 模型的构建过程通过R 语言(Vision4.0.3)中的mgcv 软件包实现,假设模型的误差分布为Gaussian 分布,连接函数选择自然对数[23]。使用ArcGIS 10.2 软件绘制调查站位图以及生物量密度分布与环境因子的关系图。
2 结果与分析
2.1 生物量密度年际变化
2014-2021 年研究区域曼氏无针乌贼平均生物量密度最大值为0.34 kg·km-2,出现在2020 年,最小值为0.16 kg·km-2,出现在2014 年。2014 年以来,曼氏无针乌贼平均生物量密度总体上呈波动上升的趋势,且2020 年为峰值(表1)。

表1 2014-2021 年曼氏无针乌贼生物量密度Tab.1 Biomass density of S.maindroni from 2014 to 2021(kg·km-2)
2.2 资源密度时空分布及变化
浙江近海曼氏无针乌贼生物量年间和季间变化明显,如图2 所示,曼氏无针乌贼主要集中分布在27°N-29°N,121°E-123°E 海域。从4 月(春季)来看,2014 年仅有5 个站位点有曼氏无针乌贼出现,最高生物量密度为30.62 kg·km-2,2015 年和2016 年其资源密度增大,主要分布在30°N 以南海域,到2017 年开始逐渐向北向南大范围增加,舟山渔场中部海域的资源密度较往年大幅增长,最高生物量密度达86.86 kg·km-2,2018 年资源密度较2017 年减少,集中分布在28°N-30.5°N 海域,2019-2021 年资源分布范围明显扩大,资源密度显著增长,最高生物量密度可达113.85 kg·km-2。

图2 2014-2021 年曼氏无针乌贼生物量密度的时空分布Fig.2 Temporal and spatial distribution of S.maindroni by biomass density from 2014 to 2021
从11 月(秋季)来看,2014-2016 年资源密度向北逐渐增加,主要分布在30°N 附近海域,其中2015 年生物量密度最大,最高可达122.50 kg·km-2,2017 年开始分布范围向南拓展,主要分布在浙江南部海域,2018 年主要分布在28°N-30°N,122°E-123°E,最高生物量密度为71.36 kg·km-2,2019 年集中分布在28°N附近,2020 年主要分布在浙江中部、浙江南部海域,最高生物量密度达155.97 kg·km-2,2021 年主要分布在浙江北部和浙江中部海域,且29°N-31°N 海域资源密度显著增加,最高生物量密度达155.49 kg·km-2。
4 月,平均资源密度最高值出现在2020 年,为0.39 kg·km-2;11 月,平均资源密度最高值出现在2021年,为0.27 kg·km-2,生物量密度最大值出现在2020 年11 月,达155.97 kg·km-2。4 月平均生物量密度和曼氏无针乌贼出现站位均高于11 月,且4 月多分布在近禁渔区线以西的近海,而11 月多分布在靠近禁渔区线附近的外侧海域。
2.3 模型选择
通过AIC 准则进行因子的筛选,根据AIC 最小原则获取最优GAM 模型拟合曼氏无针乌贼资源密度分布与各影响因子间的关系(表2、3)。结果显示,年份、月份、底层温度和底层盐度4 个因子与生物量密度呈显著相关,故模型表达式如下:


表2 GAM 模型选择Tab.2 GAM selection

表3 GAM 模型检验结果Tab.3 Test results of GAM
2.4 不同因子对资源密度的影响
根据GAM 模型拟合结果及不同因子对曼氏无针乌贼生物量密度分布的影响可得其资源分布与年份、月份、底层水温和底层盐度的相关性显著(图3),加入底层水温和底层盐度2 个环境因子后的最终拟合模型AIC 值最小。由年际效应图可看出曼氏无针乌贼生物量密度年际变化较大,2020 年比2021 年稍高且显著高于其他年份,且2014 年生物量密度最小。从季节效应的生物量密度来看,4 月(春季)生物量最高,明显高于11 月(秋季),其生物量密度具有明显的季节变化。由底层水温与生物量密度的关系可知,其适宜底层水温为14~22 ℃,14~19 ℃之间保持基本平衡,其生物量密度在19~21 ℃范围内随着底温的上升而增加,最高值出现在21 ℃附近;由底层盐度与生物量密度的关系可知,底层盐度对曼氏无针乌贼生物量密度的影响范围为16.8~35.0,盐度在25~30 范围内其生物量密度随着盐度的增高而增加,当盐度在30~35 时,生物量密度较高。
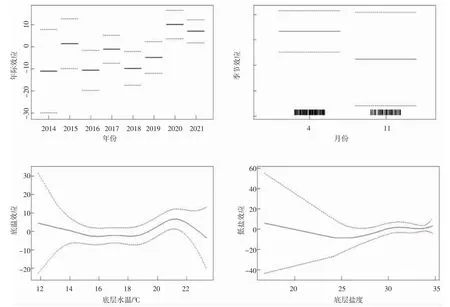
图3 2014-2021 年曼氏无针乌贼生物量密度与环境因子的GAM 模型拟合结果Fig.3 GAM simulation results of biomass density and environment factors of S.maindroni,2014-2021
3 讨论
3.1 曼氏无针乌贼季节性分布变化与其洄游生态习性密切相关
浙江近海曼氏无针乌贼资源密度年际和季节变化较大(图3)。根据曼氏无针乌贼的洄游特性可知其为一种区域性的近海活动生物,由于短距离的迁徙活动使得曼氏无针乌贼对生活海域的生态环境依赖性较强,因此乌贼的生长分布与近海人类活动密切相关,故其资源密度年际变化可能与增殖放流活动相关联[4]。20 世纪80 年代以来,由于春、夏季未产卵的“亲本乌贼”被围捕,秋、冬季幼乌贼和越冬乌贼被过度捕捞,导致乌贼群体数量骤减,使乌贼资源逐渐枯竭[24],而曼氏无针乌贼生长速度快、生命周期短,资源补充、恢复快,是理想的增殖放流种类[3,25-27]。因此,浙江沿岸进行的曼氏无针乌贼受精卵增殖放流活动[28-30],为补充和恢复其资源群体起到了重要作用;曼氏无针乌贼在浙江近海海域存在季节性时空分布,可能与其生活习性(产卵、索饵和越冬洄游)有关。有研究表明,饥饿对乌贼幼体的存活和游动行为影响显著[31],4 月(春季)近海靠岸水温升高快,陆地上江河流入的营养盐和潮汐、上升流等水流交汇为乌贼生存繁衍提供了丰富的饵料,可将其吸引到近岸[32]。李星颉等[33]研究表明,乌贼一般产卵在附着物上,集中在10 m 水深内最多,其次是10~20 m 水深,沿岸地带较外海地区更窄更浅,为雌雄个体相遇交配创造了条件,且更适合乌贼产卵,有利于提高其繁殖力。11 月(秋季)随着气温的下降,水温逐渐降低,而外海水层较近海厚,热散发慢,降温的速度也慢,乌贼为了越冬从近岸向外海深水层进行洄游[4,34-35]。唐逸民等[36]研究发现,在一定温度范围内,温度与曼氏无针乌贼卵的孵化速度呈正相关,近岸水温升高,缩短了乌贼卵和幼乌贼的发育期,丰富的浮游生物也为幼乌贼的生长创造了条件。因此,曼氏无针乌贼4 月(春季)在近海资源密度大。
3.2 环境因子对曼氏无针乌贼资源分布的影响
模型拟合结果显示,曼氏无针乌贼的数量分布主要受底层水温和底层盐度2 个环境因子的影响。水温是影响乌贼生态习性的重要环境因子,对其生存、繁殖以及集群和洄游分布等都起着举足轻重的作用,且能通过影响饵料生物的资源分布对乌贼的分布产生间接影响[37-40]。李星颉等[41]研究表明人工养殖条件下乌贼生长最适温度为16~25 ℃,而自然海域乌贼生长最适温度与其相比偏低,唐逸民等[36]研究表明自然海区乌贼孵化的平均水温范围为17.48~17.87 ℃。李成林等[42]研究发现11 月胶州湾内水温降至9.4~14.0 ℃后,曼氏无针乌贼群体开始越冬洄游,这与本研究得出的适宜底层水温为14~22 ℃的结论基本一致;盐度也对乌贼的生长发育有重要影响,在不同的发育阶段其生长速度的快慢和盐度的高低相关联[43-46]。
本研究通过GAM 模型对浙江近海曼氏无针乌贼的年间变动与时间(年份和月份)、海洋环境变量(底层水温和底层盐度)的分析表明:2014-2021 年间,2020 年生物量密度显著高于其他年份,且2014 年最小;4 月(春季)生物量最高,明显高于11 月(秋季)。水温在19~21 ℃范围内,曼氏无针乌贼资源密度随底层水温升高而增加,底层水温在21 ℃附近其资源密度较大;底层盐度在30~35 范围内曼氏无针乌贼资源密度较高。曼氏无针乌贼的数量分布代表着对其生存环境的适宜程度,了解其分布对于曼氏无针乌贼的资源管理、保护以及可持续利用具有重要意义。
