海洋酸化和低氧及其节律性变化对海蜇碟状幼体的影响
2022-12-05吴钟启悦王雷陈立飞李秀保董志军
吴钟启悦, 王雷, 陈立飞, 李秀保, 董志军
1. 海南大学海洋学院, 海南 海口 570228;
2. 海南大学, 南海海洋资源利用国家重点实验室, 海南 海口 570228;
3. 中国科学院烟台海岸带研究所, 牟平海岸带环境综合试验站, 山东 烟台 264003;
4. 江苏省盐城市射阳县金洋水产原种场, 江苏 盐城 224351
化石燃料资源过度开发和利用等人类生产活动导致大气中pCO2升高, 海洋对CO2的吸收使得海洋自身的碳酸盐化学失衡, 造成海水酸度增加。自工业革命以来, 全球表层海水pH 已下降了0.1, 预计2100 年前表层海水pH 将下降0.3~0.4(Orr et al,2005)。20 世纪50 年代后, 河口和近岸海域的低氧问题也愈发严重。陆源营养盐入海通量增加导致的富营养化是近岸海域低氧区扩大的重要原因(Breitburg et al, 2018)。缺氧现象通常发生在夏季,由于水体和沉积物中微生物的矿化分解作用, 导致海水底层缺氧, 并通常伴随着局部pH 降低的酸化过程。目前, 在渤海、黄海、长江口、珠江口和南海部分海域观测到海水酸化与低氧的耦合现象(韦钦胜 等, 2017; 宋金明 等, 2021)。同时, 海水pH值和溶解氧又呈现出明显的昼夜变化特征, 白天光合作用强导致较高的溶解氧水平和pH, 而晚上呼吸作用的增强则导致较低的溶解氧水平和pH(Baumann et al, 2015, 2018; Gobler et al, 2016;Gedan et al, 2017)。
海洋酸化和低氧从多方面影响着不同营养级海洋生物, 是近岸海洋生态系统的重要环境威胁。酸化和低氧对海月水母(Aurelia aurita)螅状幼体的研究显示, 低溶解氧(dissolved oxygen, DO)可使螅状幼体的无性出芽减少~50%, 酸化对无性生殖和有氧呼吸均无显著影响(Treible et al, 2018)。胁迫因素的昼夜节律变化对生物体产生的效应并不完全一致,因波动的周期、幅度和生物体等的不同而异。研究显示, DO 剧烈波动增加了美洲牡蛎(Crassostrea virginica)对病原体的感染率和致病性, 影响甚至会遗留至次年, 而pH 波动和双因素波动联合对感染都无显著影响, DO 和pH 单独波动及联合波动都会刺激血细胞活性, 但这种活性刺激可能无法有效预防病原体感染, 剧烈波动利于寄生物感染附近种群,扩大感染区阻碍牡蛎恢复(Keppel et al, 2015)。同样,三种双壳类幼体暴露于酸化和低氧实验显示, 海湾扇贝(Argopecten irradians)幼体在恒定酸化或低氧下存活率降低、生长和发育延缓; 硬壳蛤(Mercenaria mercenaria)幼体在恒定酸化下存活率降低、在恒定低氧下发育延缓; 美洲牡蛎(Crassostrea virginica)幼体在恒定酸化下存活率降低、生长减缓, 且复合因素暴露相较单因素暴露表现出更大的负面影响; 该研究中昼夜波动的CO2和DO 未完全减轻双壳类幼体受低氧和酸化的影响,无法为其存活、生长和变态发育提供充分缓和(Clark et al, 2016)。但也有研究表明, 昼夜波动间歇性应激能在一定程度上缓解负面效应, 未来持续性的海洋酸化和缺氧对厚壳贻贝(Mytilus coruscus)的危害比波动性酸化和缺氧更严重。但如果贻贝无法获得外界能量的供应和补偿, 其酸化和缺氧会影响贻贝种群的生存和发育(Shang et al, 2020)。有关月银汉鱼(Menidia menidia)研究显示, 恒定低氧严重降低了胚胎存活率、幼鱼存活率、种群半孵化耗时、孵化时大小和孵化后幼鱼生长速率, 而恒定酸化对大多数性状并无影响。然而, 实验证实酸化和低氧的昼夜波动降低了这些恒定条件对月银汉鱼(Menidia menidia)所有响应性状的负面影响, 而且缓解程度取决于波动的幅度, 表明胁迫因素的波动可能会为沿海生物提供周期性的生理庇护, 从而促进物种对气候变化的适应性(Cross et al, 2019)。此外, 这些研究大多集中在钙化生物上, 而非钙化生物仍未得到充分研究(Davis et al, 2013)。需多因素实验设计来研究同时发生的应激源(如海洋酸化和低氧)涉及的生物响应。
海蜇(Rhopilema esculentum)隶属于刺胞动物门(Cnidaria)、钵水母纲(Scyphozoa)、根口水母目(Rhizostomeae)、根口水母科(Hizostomadae)、海蜇属(Rhopilema), 在中国、日本、朝鲜半岛沿海和俄罗斯远东地区均有分布。海蜇为大型暖水性水母,主要栖息于近岸海域, 其食用价值及潜在的药用价值使之成为我国重要的渔业资源之一(李云峰 等,2020)。自20 世纪80 年代以来, 为补充海蜇的种群资源, 辽宁、山东和浙江等多地进行了增殖放流、人工育苗和养殖, 但由于海蜇对环境适应能力较差,其资源日益匮乏的现状并未得到显著改善(王国善等, 2014)。海蜇的生活史具有水母体有性浮游世代和水螅体无性底栖世代交替出现的现象。成熟的海蜇水母体营浮游生活, 通过有性生殖产生受精卵,发育成为浮浪幼虫, 继而附着于附着基上变态为早期的螅状幼体, 螅状幼体可进行无性生殖, 因环境条件作用和生存状况差异, 进行足囊生殖形成新的螅状幼体或横裂生殖产生碟状幼体, 碟状幼体再经变态发育为水母体, 水母体根据其发育状况再细分为稚蜇和成蜇(丁耕芜 等, 1981; 孙婷婷 等,2017)。其中横裂生殖是海蜇实现螅状幼体世代向水母体世代转变的唯一途径, 海蜇早期生活史幼体阶段的种群数量直接决定海蜇成体的渔业资源产量, 海蜇浮浪幼虫的生存附着、螅状幼体的无性生殖、碟状幼体的生长发育均是影响海蜇种群变化的关键阶段(Lucas et al, 2012)。本文旨在对海洋酸化和海洋低氧这两个海洋生态环境正面临的重大变化可能对海蜇碟状幼体的影响进行探讨, 为阐明海蜇对海洋酸化和低氧的响应模式和调控机制奠定基础, 以探究未来海洋中的酸化和低氧水平对海蜇种群发展的影响水平, 对于海蜇资源的预测和评估具有重要意义。
本研究设计两个pH 水平(酸化pH 7.6; 正常pH 8.1)和两个溶解氧(DO)水平(低氧 2mg·L–1; 常氧7mg·L–1), 考虑胁迫因素恒定或昼夜节律性变化,通过测定伞部直径、收缩频率、Ca2+-ATP 酶、Na+K+-ATP 酶、酸性磷酸酶(acid phosphatase, ACP)、碱性磷酸酶(alkaline phosphatase, AKP)、过氧化氢酶(catalase, CAT)活力和丙二醛(malondialdehyde,MDA)含量等生理指标, 从生长和运动及酸碱平衡、免疫和抗氧化相关酶活力方面, 分析海蜇(R.esculentum)碟状幼体在海洋酸化和低氧及其昼夜节律变化胁迫下的生理响应。本文初步探讨了海洋酸化和低氧及其昼夜节律变化对海蜇碟状幼体生理代谢和生长发育等过程的差异影响, 为进一步研究水母类群适应气候变化及未来环境条件下生物多样性和生态进程的变化趋势提供理论依据, 为渔业发展和海洋环境监测提供参考。
1 材料与方法
1.1 实验动物
海蜇碟状幼体采集自盐城金洋水产原种场, 选择由螅状幼体释放的1 日龄碟状幼体, 当天运送至中国科学院烟台海岸带研究所牟平海岸带环境综合试验站的水母养殖实验室, 将海蜇碟状幼体置于玻璃水缸(200L)中, 加入经砂滤处理的天然海水, 连通供气装置暂养3d, 暂养期间海水温度18.0±0.5℃、盐 度(32±0.5)‰ 、 pH 8.10±0.05 、 溶 氧 浓 度(7.0±0.3)mg·L–1。暂养期内每日投喂足量卤虫无节幼体(Artemia nauplius)两次。选择状态良好、结构完整、大小一致(平均伞径4mm)的碟状幼体进行实验。
1.2 实验设计与海水化学
实验采用正交设计, 两个pH 水平(酸化pH 7.6;正常pH 8.1)和两个溶解氧(DO)水平(低氧2mg·L–1;常氧7mg·L–1)恒定处理, 以及仅pH 波动酸化处理(pH: 8.0~7.2; DO: 7mg·L–1)、仅DO 波动低氧处理(pH:8.1; DO: 3~1mg·L–1)、pH 和DO 联合波动的酸化和低氧复合处理(pH: 8.0~7.2; DO: 3~1mg·L–1), 共七种处理。暴露实验为期7d, 暴露期间7:00—19:00 为波动处理的白天条件, 19:00—7:00 为波动处理的夜间条件。每种处理设置3 个重复, 每个重复包含100只海蜇碟状幼体, 饲养于21 个20L U 型水母缸。使用气体流量控制系统调节通入空气-CO2-N2混合气体的进气量与曝气周期, 以达到目标pH 和DO 水平实现酸化或/和低氧处理(刘辉 等, 2018)。
暴露实验期间海水温度18 ℃, 每日投饵卤虫无节幼体两次, 每两天更换约2/3 经相应预处理的新鲜海水并吸底去污, 所用新鲜海水已分别按照相应处理组的海水化学要求进行预处理。每天使用YSI水质监测仪测定温度、盐度和溶解氧, 使用NBS 标准溶液校准的pH 电极监测实验水体pH。每两天于白天时段和夜间时段, 在昼夜模式切换前1h 内, 从实验水体中采集水样, 采用滴定法测定水体总碱度(total alkalinity, TA)。根据TA 值和测量参数(温度、盐度和pH), 利用CO2 SYS 软件计算得海水碳酸盐化学的溶解无机碳(dissolved inorganic carbon,DIC)、方解石饱和度、文石饱和度。
1.3 伞部直径
暴露实验开始前和结束时, 从每种处理的水母缸内随机取15 只实验个体, 放置在含有相应处理海水的培养皿中, 使用体视显微镜连接电脑拍摄软件OPT Pro,拍照测量碟状幼体的伞部直径(Heins et al, 2018)。
1.4 收缩频率
暴露实验结束后, 从每种处理的水母缸内随机取15 只实验个体, 每只碟状幼体单独放置在含有相应处理海水的500mL 烧杯内, 待稳定3min 后, 记录1min 内碟状幼体的收缩次数, 作为收缩频率(pulsation rates)(Algueró-Muñiz et al, 2016)。
1.5 酶活力
暴露实验结束后, 用相应处理的无菌海水浸泡清洗碟状幼体, 吸取其表面水分后收集在无菌离心管中, 液氮速冻并转移至–80℃冰箱保存待用。将待检测的组织样品在冰上解冻, 称取约0.10g 碟状幼体组织, 按照质量(g):体积(mL)=1:9 的比例加入0.9%生理盐水, 低温匀浆后离心10min (2500r·min–1,4 ℃), 收集上清液分装待测。组织样的Ca2+-ATP 酶、Na+K+-ATP 酶、酸性磷酸酶(acid phosphatese, ACP)、碱性磷酸酶(alkaline phosphatase, AKP)、过氧化氢酶(catalse, CAT)活力和丙二醛(malondialdehyde, MDA)含量以及组织总蛋白浓度均采用南京建成生物工程研究所试剂盒并按照说明书进行测定。
1.6 数据分析
使用SPSS 22.0 软件分析实验数据, 分别对数据进行了方差齐性检验以及正态分布检验。使用two-way ANOVA 来确定pH 和DO 之间是否存在相互作用影响。使用student-t 检验分析每个实验组和对照组之间, 以及每个对应波动处理组和恒定处理组之间差异性是否显著, 显著性水平设定为P<0.05。实验数据以平均值±标准差(Mean±SD)表示。
2 结果与分析
2.1 海水化学
本实验通过向实验海水充入CO2气体模拟海洋酸化环境, 充入N2气体模拟海洋低氧环境。根据实验测得的数据, 温度、盐度、pH、DO 均保持在实验目的环境条件范围内, 根据计算得出其他碳酸盐化学参数 (表1)。
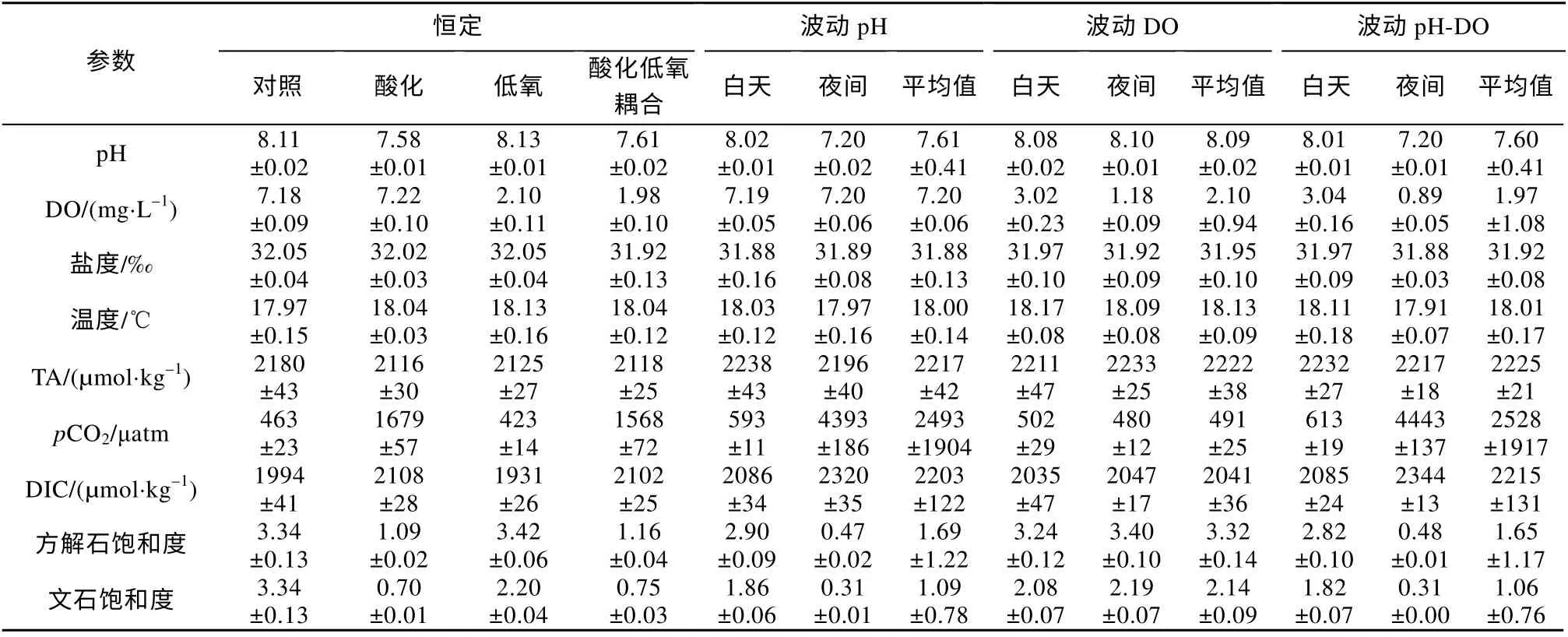
表1 本实验中相关海水化学参数Tab. 1 Chemical parameters of seawater in this experiment (mean±SD)
2.2 伞部直径
与对照组相比, 所有波动处理组的碟状幼体伞部直径均显著降低(P<0.05), 恒定和波动处理组中, 均没有观察到pH 和DO 之间的交互作用(P>0.05; 表2、3)。碟状幼体的伞部直径在酸化、低氧、酸化与低氧联合的恒定和波动处理组之间均无显著差异(P>0.05, 图1)。
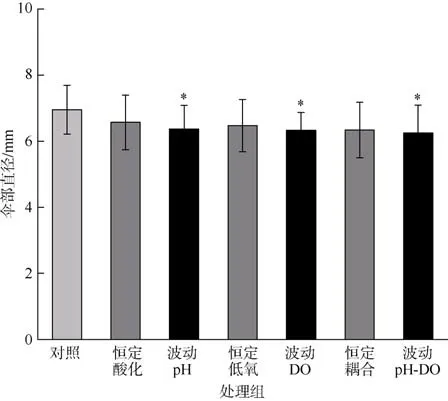
图1 海蜇碟状幼体暴露于恒定/波动的酸化和/或低氧处理后的伞部直径*表示和对照组有显著差异(P<0.05)Fig. 1 Bell diameter of R. esculentum ephyrae after constant/fluctuating acidification and/or hypoxia exposure.The * error bar indicates significant difference (P<0.05) as compared to control group

表2 双因素方差分析结果: 恒定酸化和低氧胁迫对海蜇碟状幼体的影响Tab. 2 Two-way ANOVA summary: Effects of constant acidification and hypoxia on the R. esculentum ephyrae
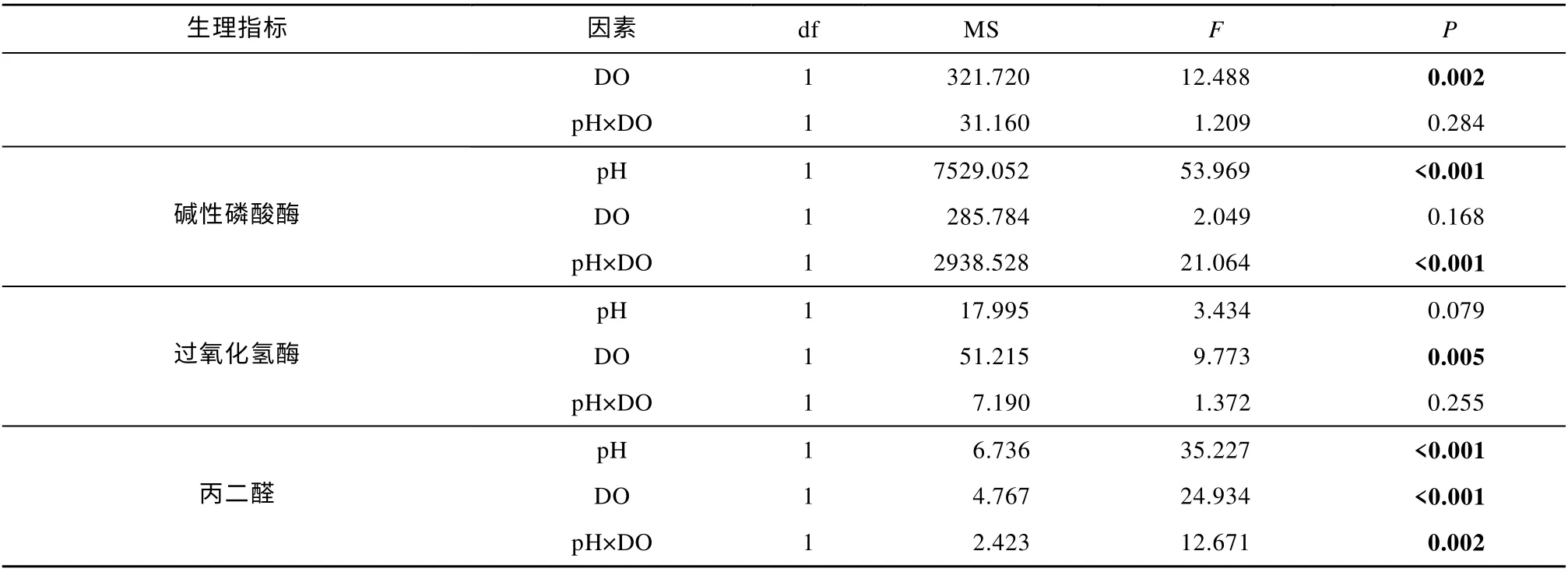
续表
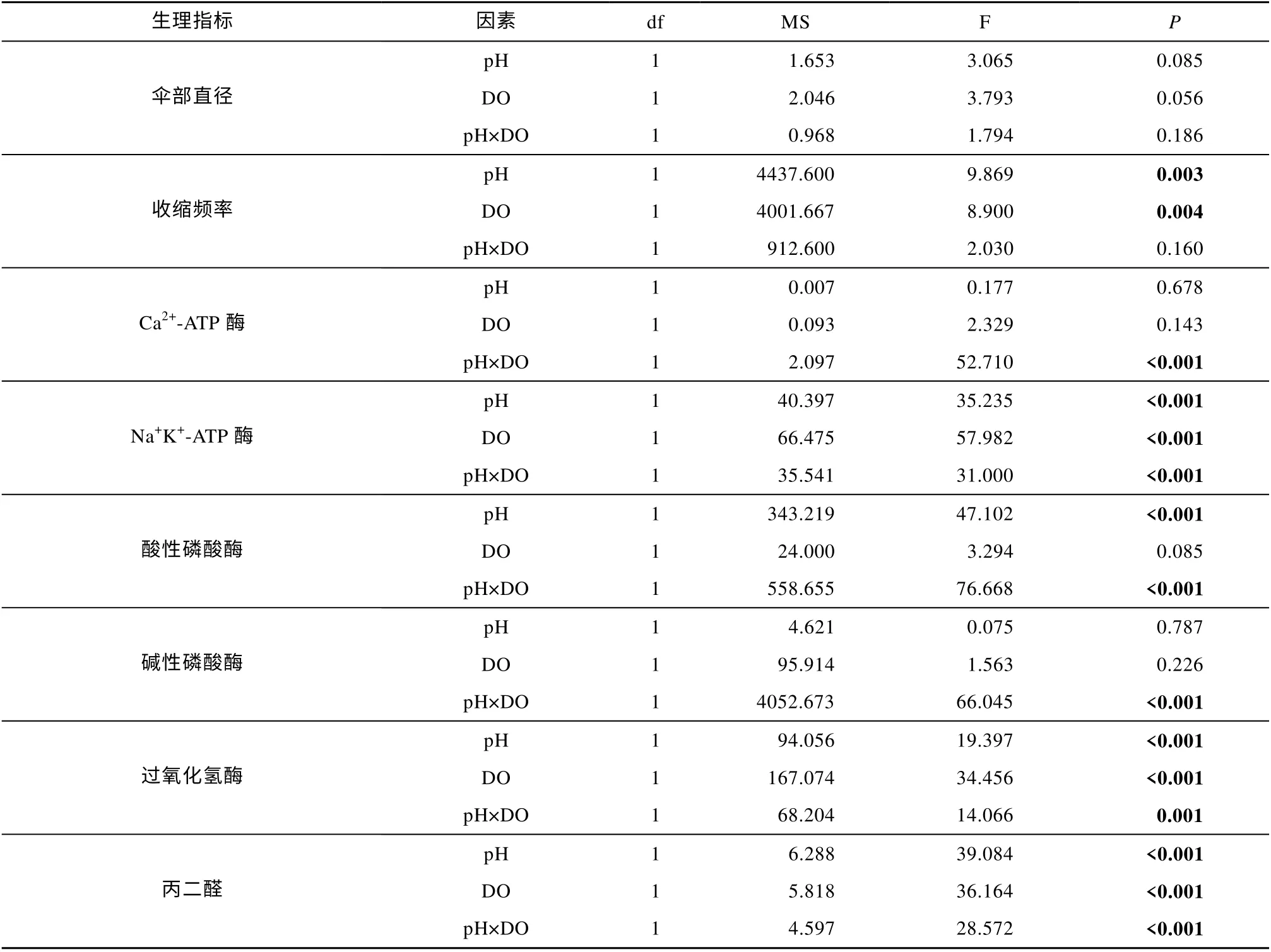
表3 双因素方差分析结果: 波动酸化和低氧胁迫对海蜇碟状幼体的影响Tab. 3 Two-way ANOVA summary: Effects of fluctuating acidification and hypoxia on the R. esculentum ephyrae
2.3 收缩频率
与对照组相比, 恒定和波动的酸化与低氧联合处理组的海蜇碟状幼体收缩频率显著降低(P<0.05)。恒定和波动处理组中, 均没有观察到pH 和DO 之间的交互作用(P>0.05; 表2、3)。碟状幼体的收缩频率在酸化、低氧、酸化与低氧联合的恒定和波动处理组之间均无显著差异(P>0.05; 图2)。
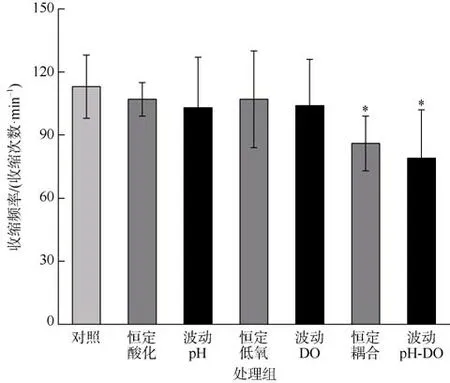
图2 海蜇碟状幼体暴露于恒定/波动的酸化和/或低氧处理后的收缩频率*表示和对照组有显著差异(P<0.05)Fig. 2 Pulsation rates of R. esculentum ephyrae after constant/fluctuating acidification and/or hypoxia exposure.The * error bar indicates significant difference (P<0.05) as compared to control group
2.4 酸碱平衡、免疫和抗氧化相关酶活力
2.4.1 Ca2+-ATP 酶和Na+K+-ATP 酶活力
与对照组相比, 恒定的酸化、低氧、酸化与低氧联合处理的海蜇碟状幼体Ca2+-ATP 酶活力均显著升高(P<0.05), 但波动的酸化、低氧处理的海蜇碟状幼体Ca2+-ATP 酶活力则显著降低(P<0.05)。在恒定和波动处理组中, pH 和DO 之间都存在显著的交互作用(P<0.001; 表2、3)。碟状幼体的Ca2+-ATP 酶活力在暴露于酸化、低氧、酸化与低氧联合的恒定处理后, 均显著高于对应的波动处理组(P<0.05, 图3a)。
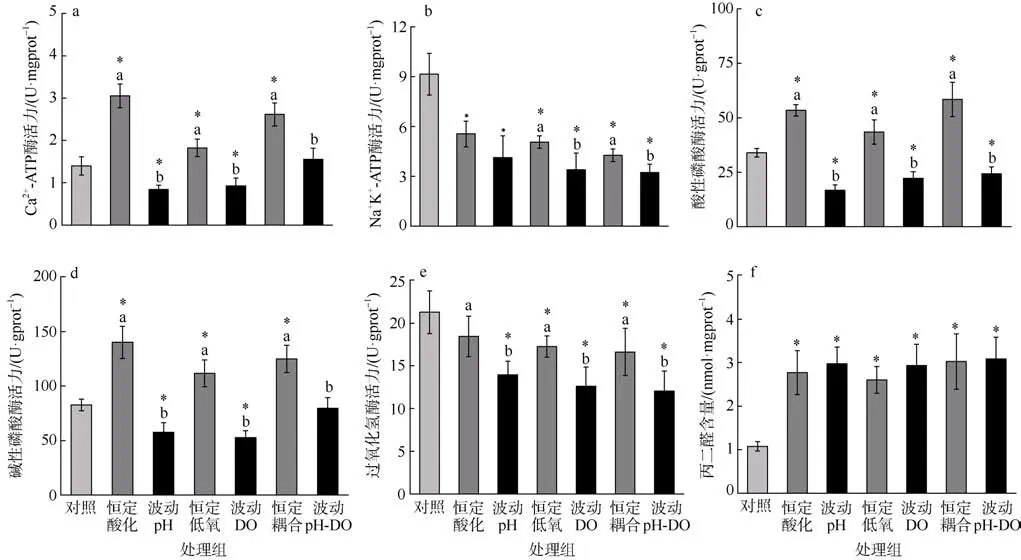
图3 海蜇碟状幼体暴露于恒定/波动的酸化和/或低氧处理后的酶活力*表示和对照组有显著差异(P<0.05); 误差棒上方不同英文小写字母表示恒定和波动的实验组之间存在显著差异(P<0.05)Fig. 3 Enzyme activities of R. esculentum ephyrae after constant/fluctuating acidification and/or hypoxia exposure. The *above error bar indicates the significant difference (P<0.05) as compared to control group; the different lowercase letters indicate significant differences (P<0.05) between constant and fluctuating treatment groups
与对照组相比, 所有处理组的海蜇碟状幼体Na+K+-ATP 酶活力均显著降低(P<0.05)。在恒定和波动处理组中, pH 和DO 之间都存在显著的交互作用(P<0.001; 表2、3)。相较于恒定处理, 碟状幼体在暴露于波动的低氧、酸化与低氧联合处理后Na+K+-ATP 酶活力显著降低(P<0.05; 图3b)。
2.4.2 酸性磷酸酶(ACP)和碱性磷酸酶(AKP)活力
与对照组相比, 恒定的酸化、低氧、酸化与低氧联合处理的海蜇碟状幼体ACP 活力均显著升高(P<0.05), 但波动的酸化、低氧、酸化与低氧联合处理组的ACP 活力则显著降低(P<0.05)。在恒定和波动处理组中, 恒定处理组的pH 和DO 之间没有交互作用(P>0.05; 表2、3), 但波动处理组的pH 和DO之间观察到交互作用(P<0.001; 表2、3)。碟状幼体的ACP 活力在暴露于酸化、低氧、酸化与低氧联合的恒定处理后, 均显著高于对应的波动处理组(P<0.05, 图3c)。
与对照组相比, 恒定的酸化、低氧、酸化与低氧联合处理的海蜇碟状幼体AKP 活力均显著升高(P<0.05), 但波动的酸化、低氧处理组的AKP 活力则显著降低(P<0.05)。在恒定和波动处理组中, pH 和DO 之间都存在交互作用(P<0.001; 表2、3)。碟状幼体的AKP 活力在暴露于酸化、低氧、酸化与低氧联合的恒定处理后, 均显著高于对应的波动处理组(P<0.05; 图3d)。
2.4.3 过氧化氢酶(CAT)活力和丙二醛(MDA)含量
恒定酸化组与对照组相比没有显著差异, 其余处理组的海蜇碟状幼体CAT 活力相较于对照组均显著降低(P<0.05)。在恒定和波动处理组中, 恒定处理组的pH 和DO 之间没有交互作用(P>0.05; 表2、3),但波动处理组的pH 和DO 之间观察到交互作用(P<0.05; 表2、3)。暴露于波动的酸化、低氧、酸化与低氧联合处理的碟状幼体, 相比于对应的恒定处理组, 其CAT 活力显著降低(P<0.05, 图3e)。
所有处理组的海蜇碟状幼体MDA 含量相较于对照组均显著升高 (P<0.05)。在恒定和波动处理组中, pH 和DO 之间都存在交互作用 (P<0.05; 表2、3)。碟状幼体的MDA 含量在酸化、低氧、酸化与低氧联合的恒定和波动处理组之间均没有显著差异(P>0.05, 图3f)。
3 讨论
3.1 酸化和低氧昼夜波动抑制海蜇碟状幼体生长
本研究表明, 不同昼夜变化的海水酸化和低氧对海蜇碟状幼体生长的影响不同, 恒定处理暴露7d后, 未出现显著差异, 而波动处理暴露7d 后, 海水酸化、低氧、酸化与低氧联合处理对海蜇碟状幼体的生长均显示显著抑制作用。有研究发现, 部分水母物种无论是底栖阶段还是浮游阶段, 都同时对环境有很大的适应性(Algueró-Muñiz et al, 2016)。海月水母(Aurelia aurita)的酸化暴露实验发现, pH 7.62~7.72 对其无致命影响(Treible et al, 2018)。大多数水母物种生活在溶解氧含量大于2mg·L–1的水体环境中, 但部分物种如大西洋金黄水母(Chrysaora quinquecirrha)和淡海栉水母(Mnemiopsis leidyi)等对低氧环境表现出耐受性(Purcell et al, 2001)。与其他浮游分类群相比, 碟状幼体阶段通常是水母生活史中最脆弱的阶段之一(Algueró-Muñiz et al, 2016)。碟状幼体在昼夜波动的酸化和低氧处理后表现出生长抑制, 可能是波动条件极值对碟状幼体摄食等方面产生的负面效应, 例如酸化等暴露下, 水母毒素的生物活性受影响, 夜光游水母(Pelagia noctiluca)的刺细胞受损并影响捕食行为(Morabito et al, 2013)。研究发现,饵料丰度是影响海蜇幼体生长发育及生存的重要因子, 碟状幼体的器官尚未发育成熟, 浮游及捕食能力较弱, 主要借助水流实现浮游和摄食, 营养对晚期碟状幼体的生长有显著的影响(刘顶海, 2011)。
3.2 酸化和低氧耦合降低海蜇碟状幼体收缩频率
海蜇碟状幼体的运动主要依靠缘叶的舒张和收缩, 通过收缩运动捕获猎物和躲避敌害等(Kikkawa et al, 2010)。本研究中, 对照组海蜇碟状幼体的收缩频率最高, 酸化或低氧的单独暴露对收缩频率没造成显著影响, 而酸化和低氧联合暴露后收缩频率显著降低, 在恒定和波动对应处理中无显著差异。有研究表明, 海蜇碟状幼体在pH 为7.15~8.15 的海水中能够正常生活, 而在pH 为7 或9 的水体中其收缩频率仅略低于正常值(宋晶 等, 2009)。Kikkawa 等(2010)研究显示, 海月水母(Aureliasp.)碟状幼体在暴露于5000μatmpCO2的酸化条件时收缩频率才出现降低趋势。Algueró-Muñiz 等(2016)研究表明, 海月水母(Aurelia aurita)碟状幼体在温度、pCO2和溶解氧的三因素胁迫下, 大多数处理组的收缩频率无显著差异, 而高温时的pCO2效应随DO 不同而不同,低DO 时呈显著正效应而高DO 时呈负效应。Winans等(2010)报道海月水母(Aurelia labiata)碟状幼体和螅状幼体在pH 7.5 和7.3 下处理后, 其存活及无性繁殖均无显著差异, 但发现低pH 处理导致平衡石减小, 每个平衡囊中的平衡石数量不变, 对平衡石的影响可能使碟状幼体表现出运动行为变化(Winans et al, 2010)。
3.3 酸碱平衡、免疫和抗氧化相关酶活力
3.3.1 Ca2+-ATP 酶和Na+K+-ATP 酶活力
当pH、DO 等环境因子发生变化时, 海洋生物需调整ATP 敏感的离子通道, 改变离子代谢速率等,从而适应环境因子的变化(肖武汉, 2014; 庄淑芳等, 2014)。Ca2+-ATP 酶和Na+K+-ATP 酶是分布于细胞膜表面的离子转运蛋白, 是环境胁迫时的敏感生物标志物(Marques et al, 2020)。Ca2+-ATP 酶对Ca2+的跨膜转运和维持酸碱平衡至关重要(Al-Horani et al, 2003)。Na+K+-ATP 酶能消耗ATP 驱动细胞膜两侧Na+、K+离子交换运输, 对于维持细胞内外渗透压平衡和离子调节等方面有重要调节作用(韩春艳 等,2016)。本研究显示, 恒定的酸化、低氧、酸化与低氧联合处理导致海蜇碟状幼体Ca2+-ATP 酶活力均显著升高, 但是波动的酸化、低氧处理的海蜇碟状幼体Ca2+-ATP 酶活力则都显著降低, 恒定处理中碟状幼体的Ca2+-ATP 酶活力均显著高于对应的波动处理组。研究表明, 日本虎斑猛水蚤(Tigriopus japonicus)无节幼体在酸化暴露后Ca2+-ATP 酶活力显著增强, 可能是环境胁迫下生物体进行离子调节作用, 造成能耗增加及对生殖、发育的负面影响(李飞 等, 2018)。海月水母(Aurelia labiata)的酸化和重金属离子暴露实验显示, 海洋酸化和 Cu2+均对Ca2+-ATP 酶活性具有较强的抑制效应, 且酸化条件会导致碟状幼体的钙质结构如平衡石受损(Winans et al, 2010)。不同程度酸化低氧胁迫大黄鱼(Larimichthys crocea)幼鱼的研究显示, 慢性低氧胁迫下其鳃Ca2+-ATP 酶活力与对照组无显著性差异,急性酸化胁迫下其鳃Ca2+-ATP 酶活力呈先增加后减少趋势, 慢性酸化、急性低氧酸化协同胁迫下,Ca2+-ATP 酶活力随时间延长趋向恢复至初始水平,发现低氧与酸化之间存在拮抗性(曾姣, 2021)。另外,酸化和低氧对华美盘管虫(Hydroides elegans)变态期幼虫蛋白质组的研究表明, 在一定程度上, 机体对低氧的生理响应可能削弱由酸化作用引起的不利影响, 酸化和低氧对生物体显示出拮抗作用(Wu, 2002;Fabry et al, 2008)。本研究中, 酸化、低氧、酸化与低氧联合处理的恒定模式和波动模式均导致海蜇碟状幼体Na+K+-ATP 酶活力显著降低, 相较于恒定处理, 碟状幼体在暴露于波动的低氧、酸化与低氧联合处理后 Na+K+-ATP 酶活力被显著抑制。Na+K+-ATP 酶活力降低表明生物体在胁迫环境中可能呈现代谢率和代谢需求降低, 与本实验结果类似,大黄鱼(Larimichthys crocea)幼鱼在急性低氧胁迫下其鳃中 Na+K+-ATP 酶活力显著低于对照组(曾姣,2021)。研究表明海水pH 降低导致的组织损伤或引起Na+K+-ATP 酶活力变化, 海水酸化胁迫导致长牡蛎(Crassostrea gigas)鳃中的Na+K+-ATP 酶活力显著下降(王晓芹, 2018)。
3.3.2 酸性磷酸酶(ACP)和碱性磷酸酶(AKP)活力
磷酸酶通过水解磷酸单酯去除底物分子上的磷酸基团, 是一类可催化相应底物去磷酸化的酶, 又称磷酸单酯水解酶, 根据其催化作用的最适pH 分为酸性磷酸酶(ACP)和碱性磷酸酶(AKP)。磷酸酶广泛存在于动物各组织中, 在去磷酸化反应、生理代谢、免疫防御和环境适应等方面起重要作用, 外源物质通过机体第一道免疫防线后, ACP、AKP 和溶菌酶(Lysozyme, LZM)等酶类物质可发挥降解作用,当免疫水平改变时磷酸酶会产生相应变化(霍达,2020)。ACP 主要位于溶酶体和内膜系统, 是溶酶体的标志酶(Pipe, 1990), 在酸性环境下催化磷酸单酯水解成酚和磷酸, 破坏表面含磷酸酯的异物, 起到预防感染的作用(徐彦, 2012); AKP 为含锌(Zn2+)和镁(Mg2+)的金属酶, 存在于绝大多数动物细胞的质膜上, 能够促进钙的吸收, 参与碳酸钙的分泌和积聚, 碱性环境下催化磷酸单脂水解成乙醇和磷酸盐,并且参与磷酸基团的转移, 在动物代谢过程中起重要的调控和解毒作用(徐彦, 2012)。本研究结果表明,恒定的酸化、低氧、酸化与低氧联合处理导致海蜇碟状幼体ACP 和AKP 活力均显著升高, 但波动的酸化、低氧、酸化与低氧联合处理导致ACP 活力显著降低, 波动的酸化、低氧处理导致AKP 活力显著降低。恒定的酸化、低氧、酸化与低氧联合处理后碟状幼体的ACP 和AKP 活力均显著高于对应的波动处理组。有研究显示, ACP 或AKP 活力升高表明环境胁迫在某种程度上诱导了机体内免疫相关防御机制,反之酶活力降低可能意味着免疫损伤, 刺参(Apostichopus japonicus)在暴露于低氧胁迫下也表现出ACP 活力显著升高(霍达, 2020)。环境适宜时ACP和AKP 活力较低, 生物体内ACP 和AKP 活力的增强表明机体免疫功能被激活, 推测碟状幼体通过采取主动调节进而维持体内免疫及酸碱平衡, 昼夜波动胁迫过度使生物免疫受损时难以调节维持稳态则表现出一种被动病理特征(朱晓闻, 2012)。同样, 温度、盐度、pH 对马氏珠母贝(Pinctada martensii)稚贝生理学特征的联合效应研究显示, pH 的一次效应和二次效应对马氏珠母贝稚贝体内ACP 和AKP 活力均有极显著影响, 当存在环境因子胁迫时, 免疫功能表现出一段增强期(文春根 等, 2009; 朱晓闻, 2012)。3.3.3 过氧化氢酶(CAT)活力和丙二醛(MDA)含量
当环境因子发生不利于生物体的变化时, 生物体内活性氧自由基含量升高, 活性氧作为高活性分子, 会氧化细胞成分而造成细胞膜损伤等(Valko et al, 2007)。为了保护机体免受氧自由基造成的损伤,生物体采取一系列自适应策略, 通过抗氧化系统来降低活性氧水平。抗氧化酶是抵抗活性氧的第一道防线, 例如超氧化物歧化酶(SOD)、过氧化氢酶(CAT)等(霍达, 2020)。CAT 是抗氧化系统中清除多余活性氧自由基的主要抗氧化酶之一(Magnadóttir,2006)。本研究结果显示, 除恒定酸化组与对照组无显著差异外, 其余处理组的海蜇碟状幼体CAT 活力相较于对照组均显著降低, 且暴露于波动的酸化、低氧、酸化与低氧联合处理的碟状幼体, 相比于对应的恒定处理组, 其CAT 活力显著降低。同样, 海月水母(Aurelia coerulea)碟状幼体和海葵(Exaiptasia pallida)暴露于酸化条件时, 其CAT 活力也受到显著抑制(Siddiqui et al, 2015; 王雷, 2020)。另外, 在低氧、高温、高温低氧三种胁迫下, 刺参(Apostichopus japonicus)的CAT 活力均显著降低(霍达, 2020)。研究发现, 当生物处于环境胁迫下, 海洋生物体内的抗氧化酶会发生一定的变化, 仅轻度环境胁迫时,抗氧化酶体系会显著升高, 而当处于严重的环境胁迫下时, 生物体的有关酶活力会显著下降(张培玉等, 2007; 姜娓娓 等, 2017)。丙二醛(MDA)是脂质过氧化作用的最终产物, 可以作为氧化应激损伤的主要生物标志物, 反映生物体内氧自由基引起的应激压力(del Rio et al, 2005)。本研究中, 酸化、低氧、酸化与低氧联合的恒定和波动处理组的海蜇碟状幼体MDA 含量相较于对照组均显著升高。实验表明,环境压力造成机体氧化应激而引起脂质过氧化, 进而对生物体的组织和细胞造成氧化损伤, 低氧胁迫和高温低氧胁迫下刺参(Apostichopus japonicus)中MDA 含量极显著升高与本实验海蜇碟状幼体结果相似(霍达, 2020)。Shi 等(2005)表明MDA 积累程度与氧化应激水平呈正相关。环境胁迫诱导产生了大量活性氧自由基且无法被有效清除, 生物体遭受氧化损伤, 与此同时抗氧化酶活力受抑制, 脂质过氧化程度加剧, 因而作为脂质过氧化作用代谢产物的MDA 含量就会增加(Martínez-Álvarez et al, 2005)。
综上所述, 海洋酸化和低氧在不同程度上影响海蜇碟状幼体的生理代谢和生长发育, 且酸化和低氧之间表现出部分拮抗作用, 如对 Ca2+-ATP酶、酸性磷酸酶和碱性磷酸酶活力的影响。整体上,昼夜波动的暴露模式对海蜇碟状幼体的损伤程度大于恒定的暴露模式, 表明碟状幼体为海蜇生活史中极为脆弱的阶段, 关于海蜇其他生活史阶段对酸化和低氧胁迫不同暴露模式的响应还有待研究, 进一步揭示钵水母类群适应气候变化和抵抗逆境的关键机制。
