咖啡中丙烯酰胺的形成与控制研究进展
2022-11-30朱雨辰
柏 杰,朱雨辰,陈 芳
(中国农业大学食品科学与营养工程学院,国家果蔬加工工程技术研究中心,北京 100083)
丙烯酰胺(arylamide,AA)是一种易溶于水、乙醇等极性溶剂的化合物,易挥发,可通过皮肤、呼吸道、食道等进入人体内,其中饮食摄入是其最主要的暴露途径。欧盟食品安全委员会(European Food Safety Authority,EFSA)最新膳食暴露评估中,AA在日常饮食摄入的平均暴露水平为0.4~1.9 μg/(kgmb·d)[1]。国家食品安全风险评估中心研究数据显示,2012年我国AA的人均摄入水平为0.319 μg/(kgmb·d),与2000年相比增长70%,呈现逐年升高趋势[2]。AA进入机体代谢后可产生肝脏毒性、神经毒性、生殖毒性和致癌性,被国际癌症研究署(International Agency for Research on Cancer,IARC)列为2A类致癌物[3],从2002年在食品中发现至今,一直为国际食品安全领域的研究热点。
咖啡是世界三大饮料之一,全球已有80多个国家和地区种植咖啡。我国咖啡种植地以云南、海南为主,云南省咖啡种植面积和产量均占全国98%以上[4-5],是我国最大的咖啡生产基地。2011—2020年,云南省和海南省咖啡年产量总体呈上升趋势(表1),咖啡原料豆及深加工产品远销欧盟、美国等55 个国家和地区。我国咖啡消费力也不容小觑,2021年我国饮用咖啡的消费者达3.3亿 人,且咖啡消费年增长率达20%,远高于全球2%的年均增长率[6],市场需求持续增加。咖啡中含有咖啡因、绿原酸、葫芦巴碱等多种生物活性成分,具有抗氧化、降血脂、降低心血管疾病风险的功效[7]。但研究表明,咖啡中AA含量较高,有的品种甚至高达890 μg/kg[8]。波兰国民膳食AA(0.43 μg/(kgmb·d))的来源中,咖啡占比19%,仅次于面包、薯片[9]。日本一项研究表明,饮用咖啡的消费者AA暴露水平(169 ng/(kgmb·d))明显高于其他消费者(75 ng/(kgmb·d))[10]。可见咖啡是摄入AA的重要来源。
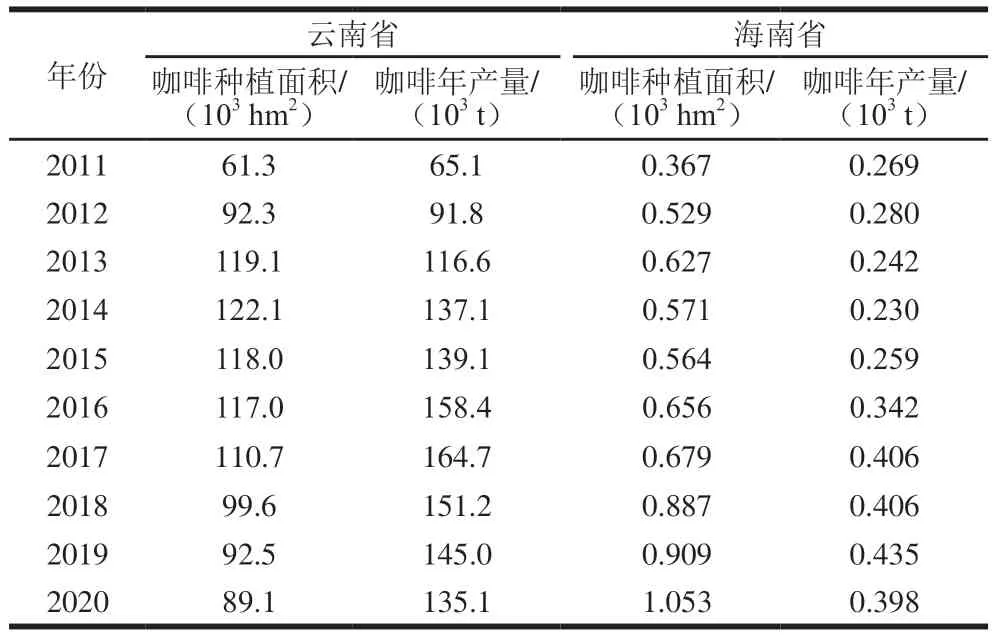
表1 我国咖啡主要生产地种植面积及产量统计[11-12]Table 1 Statistics of planting area and output of major coffee production areas in China[11-12]
随着咖啡消费群体数量的不断扩大,通过咖啡摄入AA的食品安全风险日益受到消费者关注。因此,本文综述了AA毒性以及咖啡中AA形成机制、关键影响因素、控制方法等方面的研究进展,并对未来管理法规及控制技术应用等进行了展望。
1 AA毒性机制
食品中的AA进入机体后,在单加氧酶细胞色素P450 2E1酶的催化下生成环氧丙酰胺(glycidamide,GA),AA与GA都可与还原性谷胱甘肽(glutathione,GSH)结合[13],导致活性氧含量增多,氧化损伤与抗氧化损伤系统之间不平衡,引发氧化应激[14-15]。AA通过引起细胞内活性氧、活性氮等含量增多[16],引发线粒体功能障碍等氧化损伤,诱导细胞凋亡,进一步对肝脏系统、神经系统、生殖系统等产生危害。
AA被机体吸收后,率先在肝脏中代谢,代谢过程中引发氧化应激,造成肝脏损伤[17-18]。采用25 mg/(kgmb·d)AA灌胃处理大鼠21 d后,大鼠肝脏中GSH、谷胱甘肽转移酶(glutathioneS-transferase,GST)等氧化应激指标降低,造成肝脏细胞损伤[19]。进一步分离出大鼠肝细胞发现,AA可以激活半胱氨酸蛋白酶-3(Caspase-3),引发线粒体、溶酶体损伤,从而介导肝细胞凋亡[20]。Tan Xintong等[21]在小鼠的每日饮用水中加入质量分数0.003% AA,饲养16 周后发现AA组小鼠的肝脏线粒体呈现不规则形状,PTEN基因诱导激酶蛋白1等自噬蛋白表达水平增加,线粒体自噬加剧,从而加速细胞凋亡和坏死。可见高、低剂量的AA均能破坏线粒体结构,进一步导致肝脏细胞凋亡。肝脏能代谢和吸收90%以上的AA,被吸收的AA随血液进入机体全身。
代谢吸收的AA可对神经系统造成一定的损伤。流行病学研究表明,AA对职业暴露人群具有神经毒性。目前临床研究的慢性AA中毒患者通常表现为四肢麻木、肌力减退,甚至肢端肌肉萎缩、无法行走[22]。接触AA的工人血清中肿瘤标志物神经元特异性烯醇化酶水平显著升高[23],可进一步诱发神经母细胞瘤等神经性疾病。动物水平上,AA的急性和慢性中毒均能引起中枢和周围神经损伤,造成大鼠、小鼠的神经元退化和海马体损伤,行为学表现为四肢力量不足、运动障碍和空间记忆能力降低[24-26]。有学者持续用0.2~2.0 mg/(kgmb·d)的AA灌胃Wistar大鼠5 周,结果表明AA能激活小胶质细胞,并引起脑组织中白细胞介素(interleukin,IL)-1β、IL-6和IL-18等蛋白表达水平上调,这些促炎因子可以进一步诱导大脑皮层发生炎症反应[27],从而破坏血脑屏障,影响神经系统内的物质转运,造成神经系统的功能紊乱和代谢失调。细胞水平上,AA可通过多条通路引发神经细胞的氧化应激反应,通过激活小鼠星形胶质细胞和小胶质细胞的核因子E2相关因子2、核因子κB(nuclear factor κB,NF-κB)通路,导致细胞内活性氧与抗氧化系统失衡,进而引起细胞炎症反应,并造成细胞凋亡[14,28]。研究发现,AA可破坏神经细胞线粒体结构,造成线粒体DNA损伤、能量代谢失调[29],最终导致细胞凋亡或坏死。可见在神经系统中,氧化应激、线粒体功能障碍是AA发挥毒性的重要机制。
AA具有较强的生殖毒性,能够影响动物的生育能力和生殖器官正常活动。Haleem等[30]对雄性白化鼠进行持续8 周的AA灌胃处理(10 mg/(kgmb·d)),发现AA可激活雄性大鼠睾丸内NF-κB通路,并下调紧密连接蛋白表达,氧化应激状态的持续增强使睾丸细胞凋亡数量增多、睾丸间质细胞功能异常,导致睾丸毒性。AA灌胃Wistar雌性大鼠血清中孕酮和雌二醇等激素水平降低,不同生长阶段的卵泡均发生凋亡,卵巢内细胞凋亡和自噬增多,卵泡刺激素、黄体生成素等激素分泌异常,影响卵泡发育和黄体形成,最终导致卵巢功能障碍[31]。AA通过加剧生殖器官内氧化应激反应,引起细胞凋亡和组织病变,降低精子活力、卵巢或睾丸质量[32-34],使动物的生育能力下降。
氧化应激是AA在机体中引发各种损伤的主要途径,除肝脏毒性、神经毒性和生殖毒性外,AA也会对膀胱[35]、肾脏[36]等器官产生毒性,并能加剧肿瘤发展进程[37],还有研究表明膳食AA的摄入显著增加了子宫内膜癌和卵巢癌的发病率[38]。因此,降低AA在食品中的含量可以从源头上减少AA的毒性危害。
2 咖啡中AA的形成机制
咖啡中的AA主要来源于还原糖与天冬酰胺(asparagine,ASN)发生的美拉德反应。生咖啡豆中蔗糖相对含量占5%~9%,在烘焙中蔗糖的糖苷键断裂,分解产生果糖和葡萄糖[39-40],是生成AA的重要前体物质[41],ASN相对含量占0.70%~1.26%,与蔗糖相比相对含量较低,是AA生成的主要限制因素[42]。
高温烘焙过程中,咖啡豆处于低水分含量、酸性条件(pH 4.5~5.5)下,果糖易发生烯醇化反应,生成高活性的呋喃果糖基阳离子(fructofuranosyl cation,FFC)[43],FFC在低水分活度下极易环化脱水,形成5-羟甲基糠醛(5-hydroxymethylfurfural,HMF)[44]。葡萄糖则可以与氨基酸发生美拉德反应形成Schiff碱,随后Schiff碱发生1,2-烯醇化,生成1,2-烯胺醇[45],并进一步脱氨脱水缩合形成HMF。此外,葡萄糖可以脱水形成3-脱氧葡萄糖酮(3-deoxyglucosone,3-DG)[46],3-DG进一步脱水形成HMF。
果糖、葡萄糖通过多种途径生成的HMF,与ASN反应生成中间体3-氨基丙酰胺(3-aminopropionamide,3-APA),并经历脱氨形成AA[47-48]。在250 ℃条件下加热蔗糖,最终生成的HMF中90%来源于果糖,10%来源于葡萄糖[44],可见FFC的快速反应大大加快了果糖的消耗。此外,葡萄糖和果糖也能相互转化[49],在果糖快速消耗的情况下,葡萄糖可以烯醇化成果糖参与生成AA。HMF是美拉德反应在酸性条件下的常见产物,由于咖啡果糖烯醇化和FFC脱水成HMF的反应速率高于其他途径,因此,果糖-FFC-HMF反应途径是咖啡烘焙中AA形成的最主要途径(图1)[40,50]。
3 咖啡加工工艺对AA生成的影响
咖啡的生产制备流程主要包括原料筛选、采后处理、烘焙、研磨萃取等工艺。依据加工方式的不同,咖啡主要分为速溶咖啡和现磨咖啡,现磨咖啡又衍生出土耳其咖啡、意式咖啡、美式咖啡等多种品类。咖啡鲜果经采收、去皮、干燥、分级得到咖啡豆,咖啡豆原料中蔗糖和ASN的含量、咖啡豆研磨工艺、烘焙温度、烘焙时间以及不同的咖啡萃取冲泡方式是影响咖啡AA含量的重要因素。
3.1 原料选择
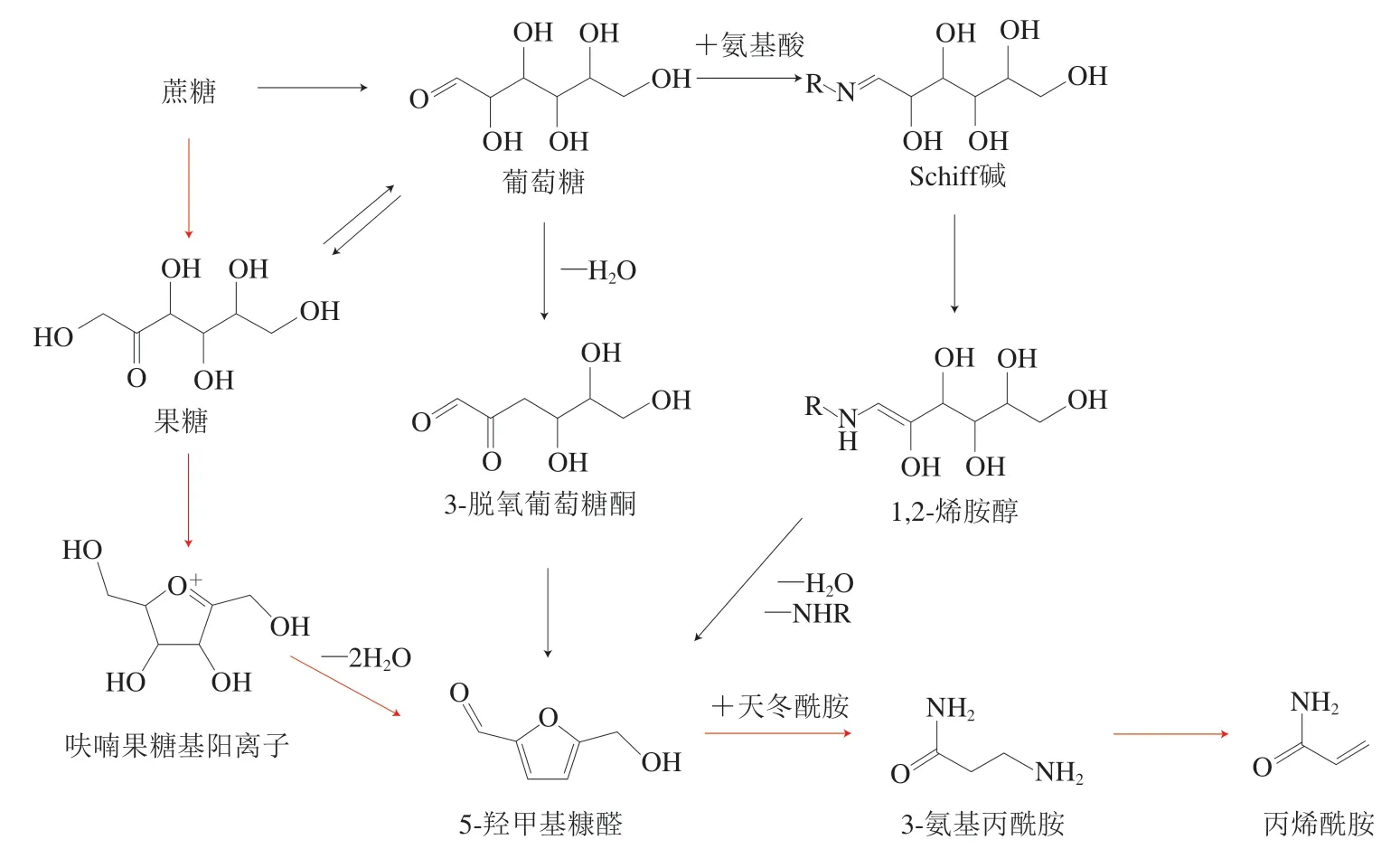
图1 咖啡中丙烯酰胺的主要形成机制Fig. 1 Major formation mechanism of acrylamide in coffee
依据咖啡豆的种类,咖啡品种主要分为小粒种咖啡(阿拉比卡咖啡)、中粒种咖啡(罗布斯塔咖啡)和大粒种咖啡(利比里卡咖啡)。利比里卡咖啡豆由于香气差、风味苦和豆粒大小不一,不适用于商业生产,目前大面积种植的咖啡豆是阿拉比卡和罗布斯塔,前者是最主要的商用咖啡品种。不同的咖啡生产原料中AA前体含量也不相同,阿拉比卡咖啡豆的蔗糖含量为73 mg/g,高于罗布斯塔咖啡豆的蔗糖含量(45 mg/g)。氨基酸含量则与之相反,阿拉比卡咖啡豆中的ASN、苯丙氨酸和丙氨酸等多数氨基酸含量低于罗布斯塔咖啡豆[51]。因此,罗布斯塔咖啡豆比阿拉比卡咖啡豆更容易生成AA,烘焙后的罗布斯塔咖啡豆AA含量(708 ng/g)明显高于阿拉比卡咖啡豆(374 ng/g)[52-53]。
3.2 烘焙工艺
采集后的咖啡果经干制法[54]或湿制法[55]加工获得生咖啡豆,再经过烘焙获得熟咖啡豆,如图2所示。通常烘焙温度为150~200 ℃,深度烘焙温度也可高于200 ℃[56-57]。烘焙工艺不仅是咖啡风味的来源,而且是影响AA含量的关键因素。AA在烘焙过程中总体呈现先增加后减少的趋势[58-60]。Hu Huihui等[61]实时监测咖啡烘焙过程中AA含量变化发现,烘烤温度从室温升高至220 ℃的过程中,在180 ℃时咖啡豆中的AA含量最高,随后AA含量持续下降。有研究将阿拉比卡和罗布斯塔咖啡豆烘焙温度分别从90 ℃上升到215 ℃,在10 min时达到175~177 ℃,阿拉比卡和罗布斯塔咖啡的AA含量分别达到最高水平(1 045±28)μg/kg和(795±25)μg/kg,随后AA含量持续下降[62]。咖啡烘焙早期AA含量的升高与ASN大量消耗相关,同时某些氨基酸可以促进AA的形成,如咖啡中的丝氨酸可以热分解形成丙酮酸、丙烯酸和游离氨来加速AA的产生[63]。随后AA含量的下降主要有两个原因:一是AA与咖啡类黑素的化学相互作用,类黑素蛋白质骨架的亲核氨基可与AA发生迈克尔加成反应[64];二是AA具有挥发性,Badoud等[65]使用14C标记AA发现,在咖啡烘焙过程中,有28%被标记的AA降解成了挥发性化合物。
咖啡豆烘焙程度是影响AA含量的关键因素。Schouten等[59]采用热空气法烘焙阿拉比卡咖啡豆,对比了浅度烘焙(烘焙6 min、最终温度147 ℃)、中度烘焙(烘焙11 min、最终温度172 ℃)、深度烘焙(烘焙18 min、最终温度191 ℃)获得的咖啡豆中AA含量以及咖啡品质,结果发现,中度烘焙咖啡AA积累量最多[66],浅度烘焙以及深度烘焙的咖啡豆中AA含量水平相对较低。深度烘焙咖啡由于咖啡烘焙时间较长,ASN和蔗糖消耗完全,类黑素含量逐渐增多,高温促进了AA与类黑素的亲核加成反应和AA挥发,使得AA含量下降。
值得注意的是,3 种烘焙程度对咖啡豆烘焙后的品质也有显著的影响。浅度烘焙得到的咖啡酸度较高、风味欠佳,苦味和涩度不足,消费者接受度低。中度烘焙咖啡的甲硫醇、2-糠基硫醇等咖啡香气成分不断积累,产生咖啡独有的香味[56],凭借其鲜明风味为消费者所喜爱。而深度烘焙咖啡伴随逐渐浓郁的香气,2-糠基硫醇含量持续增加,二甲基硫醚及部分醛类物质水平明显降低,苦涩程度较高,口感最醇厚。浅度和中度烘焙的咖啡中AA含量分别可达600~700 μg/kg和300~1 130 μg/kg,均高于深度烘焙咖啡(100~200 μg/kg)。
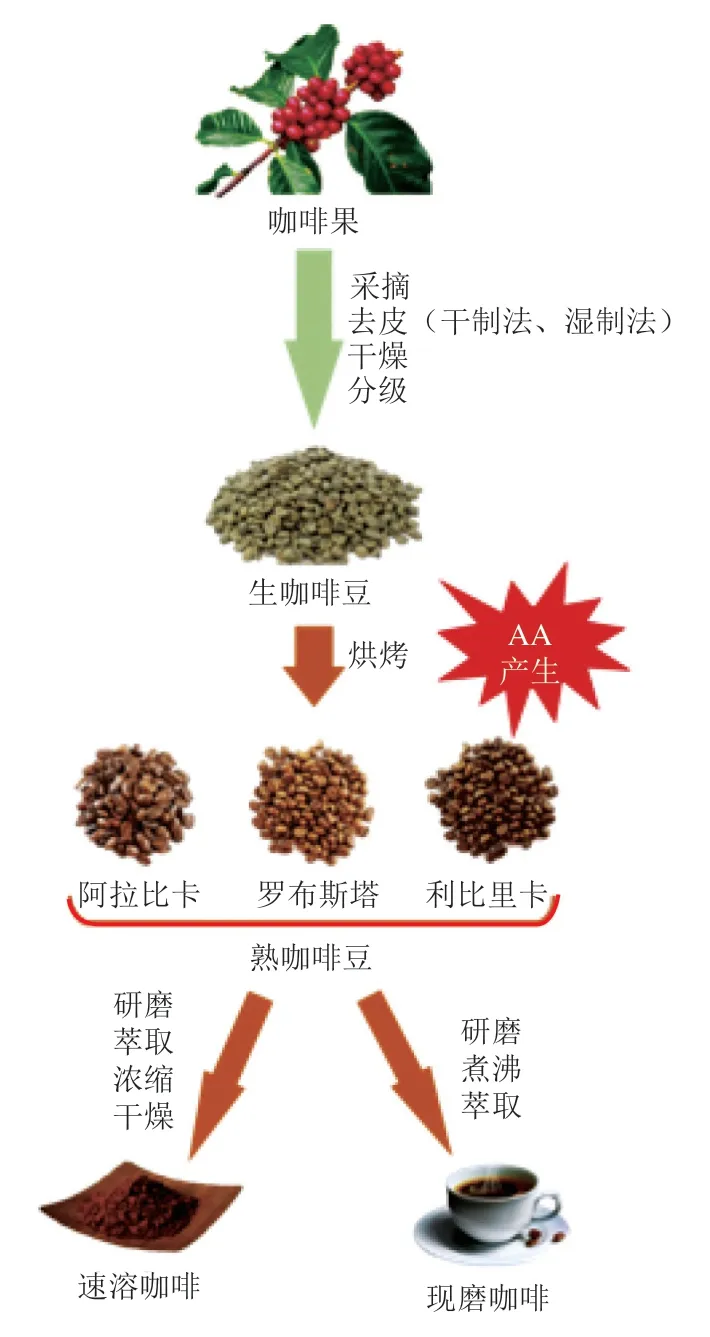
图2 咖啡的生产与加工过程Fig. 2 Production and processing of coffee
烘焙后的咖啡在不同的贮存条件下AA含量也会发生变化。姚子升[67]将焙炒后的咖啡置于-18、-4、25、37 ℃下贮存12 个月,发现37 ℃贮存最有利于降低AA水平。Delatour等[68]发现室温下贮存烘焙后的咖啡,6~12 个月后AA含量降低40%~60%。可见烘焙咖啡中的AA并不稳定,在适宜温度下长期贮存时会缓慢分解。
3.3 咖啡加工方式
现阶段,市场上较为常见的咖啡产品有现磨咖啡和速溶咖啡两种形式。随着消费水平的提高,口感丰富的现磨咖啡倍受推崇。制备工艺的不同带来了各品类咖啡的独特风味,同时造成咖啡豆烘焙程度、咖啡浓度的差异,因此AA水平也存在显著差异。
烘焙后的咖啡豆经研磨、煮沸、萃取后即为现磨咖啡。而目前现磨咖啡的种类多样,不同国家消费者对咖啡的香气、浓度等有不同的需求,从而产生多种咖啡制作工艺,也影响了终端咖啡饮品中AA含量。例如,土耳其咖啡将咖啡豆研磨极细后直接冲泡,饮用时仍有咖啡粉沉淀,咖啡中AA含量高达50.1 mg/kg[69]。意式咖啡是通过压力诱导95 ℃热水快速冲击咖啡粉、渗透萃取出的高浓度咖啡,具有强烈的香味及苦味,AA含量为38.6 mg/kg[70]。美式咖啡则是在意式咖啡基础上加水稀释制成,或采用滴滤式咖啡壶缓慢萃取,含有较低的苦味和香味,AA含量最低,为2.1 mg/kg[71]。3 种咖啡AA含量的差异主要缘于其制备过程中咖啡豆研磨、烘焙工艺上的不同。Derossi等[71]指出3 种咖啡的AA含量差异(土耳其咖啡>意式咖啡>美式咖啡)是由于咖啡豆研磨粒径不同。3 种咖啡研磨颗粒平均粒径依次为美式咖啡(710 μm)>意式咖啡(480 μm)>土耳其咖啡(320 μm)。粒径更小则更易溶解,在萃取过程释放更多AA,因此土耳其咖啡的AA含量最高。意式咖啡普遍采用高压萃取方式,咖啡浓度高,因而AA含量较高。而美式咖啡的烘焙程度和咖啡浓度低,故AA含量较低。
速溶咖啡是在咖啡豆烘焙、研磨、萃取后,进行适当浓缩,再通过喷雾干燥或冷冻干燥得到速溶咖啡粉,包装后流向市场。与现磨咖啡相比,速溶咖啡缺乏优质的香气与风味[72],且在预干燥、喷雾干燥等环节挥发了一部分AA,速溶咖啡中AA含量为96.4~346.5 μg/kg,低于现磨咖啡的AA水平(17.7~776.1 μg/kg)[73]。
4 咖啡中AA的控制方法
咖啡中AA的形成与原料的选择、加工条件和技术等密切相关。添加外源物质通过减少咖啡豆中AA前体的含量或降解生成的AA,以降低咖啡中的AA水平;优化加工技术能影响AA的生成和反应阶段,从而控制咖啡中AA的产生。
4.1 添加外源物质
ASN含量是咖啡中AA生成的主要限制性因素,生咖啡豆中ASN的浓度与烘焙咖啡的AA浓度呈正相关[74]。添加天冬酰胺酶可以降低ASN水平,进而降低咖啡中的AA含量[75-76]。Correa等[77]用5 000 U/kg天冬酰胺酶(EC 3.5.1.1)处理阿拉比卡咖啡豆2 h,烘焙得到的咖啡中AA水平下降59%,并且酶处理对咖啡中绿原酸、咖啡因、咖啡酸等活性物质含量没有显著性影响。有研究从青霉菌NMKA511中分离出的L-天冬酰胺酶(2 000 U/kg)处理阿拉比卡咖啡豆后,分别进行浅度(210 ℃、20 min)和深度烘焙(240 ℃、25 min),最终两种咖啡产品中AA相对含量分别降低了80.7%和75.8%[78]。天冬酰胺酶通过降低ASN水平从而阻碍了AA生成,而添加丙烯酰胺酶可以将生成的AA降解。从真养产碱杆菌AUM-01中提取出的丙烯酰胺酶可以将AA降解成丙烯酸和氨,采用该酶在70 ℃下处理咖啡,咖啡中AA浓度降低50%~70%(初始浓度1.4 mmol/L)[79]。Bedade等[80]使用壳聚糖和海藻酸钙制成海藻酸盐-壳聚糖复合微珠,将从草酸盐贪铜菌ICTDB921中提取的丙烯酰胺酶共价固定化,随后作用于含有质量浓度100~500 mg/L AA的速溶咖啡,30~65 ℃下60 min内可以完全降解AA。与游离酶处理相比,固定化酶可以扩大酶的最适作用温度范围,更有助于减少咖啡中AA的积累。酶可以高效地作用于相应底物,从而降低AA含量,但成本相对较高,不利于在食品工业生产中推广。
添加某些氨基酸也能减少咖啡中AA的含量。在含质量浓度6.2 μg/L AA的罐装黑咖啡中添加1.05 mg/g半胱氨酸,121 ℃下处理6 min,半胱氨酸的巯基基团能与AA的碳碳双键发生亲核加成[81-82],AA相对含量降低至原本的90%[83]。赖氨酸、甘氨酸降低AA含量的效果弱于半胱氨酸,而胱氨酸、酪氨酸的加入对AA的形成没有显著影响[84]。但半胱氨酸加热会释放出硫化氢,可能会对咖啡的风味产生不良影响。
除了降解AA前体和降解AA,在AA产生的进程中阻碍HMF的反应也是有效降低AA含量的方法。在180 ℃下,儿茶素可与HMF缩合形成二聚体加合物,从而减少AA的合成,将质量浓度5 mg/mL儿茶素添加到HMF+ASN(均为10 mmol/L)模型中,能降低70%的AA水平[85]。由此推测,将儿茶素加入到咖啡中也能有效控制AA的形成,同时降低另一危害物HMF的含量。此外,酵母菌既可以将AA前体HMF转化为羟甲基糠醇,也能将AA降解成丙烯酸。研究发现,在含1 020 μg/kg AA的速溶咖啡中加入1%酵母和10%蔗糖,发酵72 h后AA含量降低62%[86],蔗糖可为酵母细胞提供充足碳源,酵母菌发酵对人类无害且易于适配咖啡生产线,但其对咖啡风味和口感的影响仍需探究。
综上,在咖啡中添加天冬酰胺酶、丙烯酰胺酶、含有巯基的氨基酸或蛋白质、酵母菌、儿茶素等外源物质可以有效降低AA含量,但添加时需考虑对美拉德反应的促进作用和对咖啡风味的影响。
4.2 优化加工技术
咖啡的工业化生产中,常采用滚筒烘焙机常压加热咖啡豆,而新型咖啡加工技术可以在提高烘焙效率、缩短烘焙时间的同时降低AA的含量。Rattanarat等[87]采用250 ℃的过热蒸汽烘焙罗布斯塔咖啡豆,与常规的热空气烘焙法相比,烘焙温度和水分活度显著提高,烘焙时间显著缩短,同时过高温度下AA挥发和分解增强,所获得的中度烘焙(亮度值25)和深度烘焙(亮度值20)的咖啡中AA相对含量分别降低了84%和75%。此外,真空烘焙也是减少咖啡中AA含量的有效手段,低压条件下水分蒸发速率加快,对水溶性低分子质量化合物产生汽提作用,从而提高了AA的挥发速率。与常压烘焙的咖啡相比,200 ℃下真空烘焙(0.15 kPa)的阿拉比卡咖啡豆中AA相对含量减少约50%[88]。
抑制咖啡中AA形成的措施多种多样,适当地联合采用多种措施还有一定的协同作用,例如在使用天冬酰胺酶之前先用热蒸汽法预处理咖啡豆,可以增大咖啡豆内部孔隙直径,从而增大酶与咖啡豆中ASN接触的表面积,提高酶的利用率[77]。添加天冬酰胺酶、丙烯酰胺酶、酵母等外源物质,以及应用过热蒸汽、真空处理等新型加工技术均有利于减少咖啡产品中AA含量,但相关研究尚不完善,未来可增加对多种加工手段对比或协同作用的研究,咖啡工业生产中AA的控制措施仍值得探讨。
5 结 语
咖啡作为风靡全球的饮品,欧美、阿拉伯地区的众多国家饮用咖啡已成日常习惯,近年来我国咖啡的消耗量也持续增大。咖啡摄入已成为膳食中AA暴露的重要来源之一,会给人体带来不容忽视的AA毒性风险。欧盟委员会在2017年提出食品中AA抑制措施的应用建议和AA的推荐限定水平,其中建议咖啡中AA含量应当低于400 μg/kg[89]。而我国现有的食品安全法规尚未明确食品中AA的安全限量和抑制措施。此外,我国咖啡产量近10 年来已大幅上升,以云南小粒咖啡为代表的中国咖啡正逐渐扩大市场,而我国本土的咖啡加工技术、AA等危害物控制研究较少。本文总结了AA的毒性和咖啡中AA控制的工艺和技术,可为推动制定我国相关食品AA安全限量标准提供一定的参考。
从减少AA摄入考虑,更推荐消费者饮用深度烘焙咖啡,但不同国家和地区的消费者对咖啡烘焙程度存在一定的偏好性,例如美国消费者更喜欢轻度烘焙咖啡,中度和深度烘焙咖啡更受南欧消费者青睐,引导消费者统一选择深度烘焙咖啡并不现实,应当采取一定的措施减少各种类型咖啡中AA的形成,从而降低通过咖啡摄入AA的风险。然而,目前咖啡中AA的控制研究还停留在实验阶段,尚未在工业化生产规模中得到验证,对于是否会产生其他有害物质也未全面探究。因此,咖啡中AA的抑制措施仍是亟待解决的食品安全问题。未来针对AA在食品工业化生产中的控制措施及作用机理仍需进一步深入探究,以便后续为科学评估和合理制定食品中危害物限值提供科学依据。
