遮阴和施氮对黄檗幼苗叶片光合特性和碳氮计量特征的影响
2022-11-26孙悦燕郭跃东
孙悦燕,郭跃东*
(1 山西农业大学 林学院,山西太谷 030801;2 山西省重点实验室北方功能油料树种培育与研发,山西太谷 030801)
全球气候变化趋势下,植物光合固碳影响着全球生态系统碳循环,是评价碳循环的重要方法。森林生态系统对气候变化的物种响应特征在当今生理生态学上倍受关注,尤其是光因子和土壤氮(N)含量对植物生长发育和森林动态演替起主要作用,日益成为研究热点。天然林地光照资源和土壤氮素空间分布的异质性,客观上影响着林下幼苗光合能力,对其生长和林分动态产生重要影响,进而限制了植物对光、氮的利用效率,对植物碳代谢起着重要的调节作用。2000~2010年间,大气中氮化物含量日益增加,中国北部年氮总沉积速率达22.6 kg/hm2[1],2011年山西北部年氮沉降总量达到47.86 kg/hm2[2]。氮沉降加剧了土壤和叶片中氮含量的增加,对森林更新、土壤动物、微生物、生物多样性等产生了一定影响[3-5]。森林光照异质性对种子萌发、物种更新及生长[6]影响显著。有关光氮交互作用对喜光植物生长发育影响的研究表明,在强光条件下,植株光合及呼吸速率、光合潜力达到最大,而光强增加也致使植物光合对氮的需求量呈现增加趋势[7-9];遮阴(弱光)能有效降低植物的光合速率[10],叶片氮素增加也能显著提高其光合有效速率[7]。同时,植物能通过调节生物量分配,提高单株地上部分的生物量,增加叶片比叶面积和含氮量,有效提高自身光能利用率[11-12],虽然苗木生长对低氮具有一定的适应性,但持续氮素缺乏也会严重抑制苗木生长和存活[13]。森林树种只有在光强和氮素水平达到一定强度时,才能有效完成其更新过程。森林树种生态化学计量特征可反映林木体内养分吸收利用状况。外源氮输入直接影响植物养分元素C、N含量及C/N[14],增强林木光合能力,提高净光合速率及氮素光合利用率[15-16]。
黄檗 (Phellodendronamurense)属于芸香科(Rutaceae)黄檗属(Phellodendron),国家二级保护植物[17],为中国东北阔叶红松林顶极植被的“三大硬阔”树种之一,主要分布在东北地区小兴安岭和长白山林区(海拔300~1 200 m),在华北燕山山脉、北京、内蒙古、山西有少量分布[18-19]。黄檗喜光肥[20],林下不能正常生长[21]。在群落演替过程中,若无林隙或人工干预,黄檗幼苗由于林下光照不足,难以正常生长[17,20,22-23]。但也有研究发现黄檗具有一定的耐阴性,在全光照和郁闭度较高的林下也能完成有效更新[24-26]。可见,有关黄檗的耐阴性存在争议,关于光氮交互作用对林木光合、形态及生态化学计量元素影响鲜有报道。为深入了解黄檗的生态习性,尤其是其幼苗的耐阴性,本研究以黄檗3年生幼苗为对象,分析光氮复合作用对其光合特性和叶片形态的影响,旨在揭示黄檗幼苗对光照异质性和氮素的需求及其光合生理和生态化学计量特征对光氮复合作用的响应,以期为今后黄檗引种栽培及针阔混交林建设提供参考。
1 材料和方法
1.1 试验材料
供试2年生黄檗幼苗由山西省襄汾县国营苗木有限公司(35°38′ N, 116°02′ E)引栽于山西农业大学林业站(37°25′N、112°34′E),经一年缓苗,长势良好。种植地为壤土,其表层土壤全氮含量为2.48 mg·g-1,有机质含量为1.86 mg·g-1,田间持水量为27.64%。2018 年3月进行3年生黄檗幼苗田间移栽,株行距2.5 m×3.0 m,确保彼此互不影响,供足水分,及时除草,挑选均匀一致(苗高、地径及顶芽)的苗木,于5月下旬进行遮光和氮添加处理。
1.2 试验设计
采用光氮双因素随机区组设计,分别设置3个光照水平和3个氮添加水平,共组成9个处理组合,每个处理组合设4个区组(重复),每重复1株,共选取36株幼苗。整个试验期间保证苗木水肥充足,无杂草生长,8月底对试验处理材料进行气体交换参数和相对叶绿素含量的测定。
其中,3个光照水平分别为:(1)对照(S0),全光照100%,光照度为1 854.2 μmol·m-2·s-1;(2)轻度遮光(S1)[27],设置1层遮阳网透光率为对照(S0)的21.4%,光照度为396.8 μmol·m-2·s-1;(3)重度遮光(S2),设置2层遮阳网透光率为对照(S0)的8.7%,光照度为162.2 μmol·m-2·s-1。S2光强水平参考林隙自然环境下的光环境(9%~5%)设置[28-29]。
氮肥采用含氮质量分数为46%的尿素,距离植株0.15~0.3 m直径范围内,环状施肥,均匀撒开,覆盖1层1~2 cm厚土壤后用喷壶均匀喷洒3 000 mL自来水,使氮肥融化渗入土壤。3种施氮水平分别为:1)对照(F0),不施氮肥;2)轻度氮添加(F1),添加尿素总量13 g(28.21 g·m-2);2)重度氮添加(F2),添加尿素总量26 g(纯氮56.42 g·m-2)[16]。氮肥于6月和7月上旬分2次施加,第一次施撒尿素分别为5 g(F1,纯氮10.85 g·m-2)和10 g(F2,纯氮21.70 g·m-2),第二次分别为8 g(F1,纯氮17.36 g·m-2)和16 g(F2,纯氮34.72 g·m-2),分别占各施氮水平总量的38.46%、61.54%。
1.3 测定指标及方法
1.3.1 相对叶绿素含量用便携式叶绿素含量测定仪(SPAD-502,Japan)测定黄檗幼苗叶片相对叶绿素含量(SPAD值),选择试验处理幼苗第3侧枝上的第3-4对,当年生、健康无病虫害叶片,每片叶子按主脉分左、右和叶尖测量3次,取平均值作为该叶片相对叶绿素含量值。
1.3.2 气体交换参数叶片选取方法同1.3.1,于9:00-14:00之间,使用Li-6400光合仪(Li-COR Inc.,Nebraska,U.S.A)配备的标准叶室与Li-6400-02B红蓝光源,在光合有效辐射为1 200 μmol·m-2·s-1,叶温30 ℃,相对湿度60%时进行净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间二氧化碳浓度(Ci)、大气二氧化碳浓度(Ca)等气体交换参数的测定,然后按照公式计算气孔限制值(Ls)和瞬时水分利用效率(WUE)。
Ls=1-Ci/Ca
WUE=Pn/Tr
1.3.3 化学计量元素C、N含量选取叶片方法同1.3.1,105 ℃杀青120 min,65 ℃烘干48 h至恒重,研钵手工研磨,去除主次叶脉后过筛(200目),用百万分之一天平称取样品2~3 mg,放入锡舟,并用镊子将锡舟包好,清除锡舟外样品,保证样品质量。采用色谱技术(PE2400 Series II CHNS/O 元素分析仪),CHN 模式测定叶片全碳和全氮元素含量,并计算C∶N及瞬时光合氮利用效率(PNUE),PNUE=Pn/N。
1.4 光氮互作处理综合效应评价
采用模糊数学中的隶属函数(Membership)法[7],对采集的黄檗幼苗生理及化学计量元素指标进行光氮处理效应综合评价:
Ui=(Xi-Xmin)/(Xmax-Xmin)
如果为负相关, 则用反隶属函数进行转换, 计算公式为:
Ui= 1-(Xi-Xmin)/(Xmax-Xmin)
上述公式中,Ui是第i个指标的隶属函数值;Xi是指第i个测定指标;Xmax指该指标的最大值;Xmin指该指标的最小值。Ui值介于0~1之间,越大表明幼苗对遮光适应力及其对氮添加的耐性越强。按照隶属函数均值大小划分各处理黄檗幼苗抗逆性,1≥Ui>0.6为高效型,0.6≥Ui>0.5为中效型,Ui≤0.5为低效型[7]。
1.5 数据处理
采用R语言(R x64 3.5.1)软件(Robert Gentleman and Ross Ihaka,the Statistics Department of the University, Auckland),以ps.aov包进行双因素方差分析,并利用agricolae包LSD法进行多重比较(α=0.05)。
2 结果与分析
2.1 遮光和氮添加对黄檗幼苗叶片气体交换参数和叶绿素含量的影响
表1显示,黄檗幼苗叶片的气体交换参数净光合速率(Pn)、蒸腾速率(Tr)、水分利用率(WUE)、水分限制值(Ls)、气孔导度(Gs)以及相对叶绿素含量(SPAD值)均受到遮光、氮添加显著影响(P<0.05),除Pn外还受到光氮交互作用的极显著影响(P<0.01);同时,各指标光、氮单因素方差分析的F值表现为光照因素大于氮添加因素,说明光照是影响黄檗幼苗相对叶绿素含量和气体交换参数的主要因素。
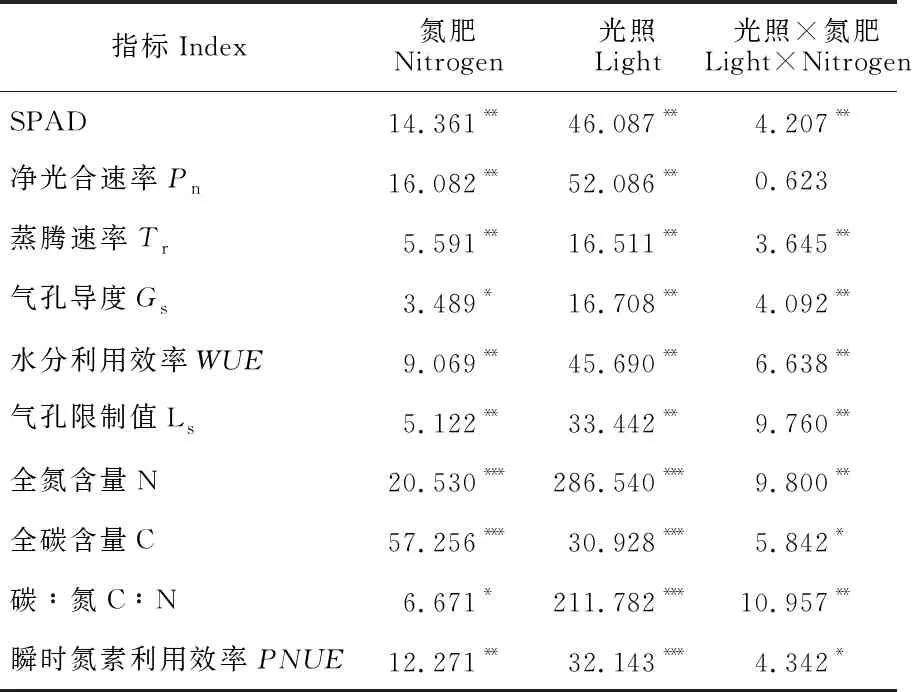
表1 光氮交互下黄檗幼苗叶片光合气体交换参数和化学计量特征的方差分析(F值)
2.1.1 遮光效应表2显示,与全光环境S0相比,轻度遮光(S1)处理黄檗幼苗叶片的SPAD值、Tr和Gs均显著增加(P<0.05),分别达到S0的1.20、1.20和1.32倍,而其Pn则稍有降低,其WUE和Ls则均显著降低;重度遮光(S2)处理的SPAD值仍显著高于S0,却较S1显著降低,其Gs与S0无显著差异,而其Pn、Tr、WUE和Ls则均比S0显著降低;尤其是S2处理的Pn仅为S0的63.05%,也比S1处理显著降低33.22%(P<0.05)。遮阴能诱导黄檗幼苗通过增加叶绿素合成来适应弱光环境,但却明显抑制了叶片的净光合速率和水分利用效率。
2.1.2 氮添加效应表2同时显示,与不施肥对照F0相比,轻度氮添加(F1)黄檗幼苗叶片的SPAD值和Tr分别显著降低10.50%和15.06%,而其余指标则均无显著变化;重度氮添加(F2)黄檗幼苗的SPAD值、Pn、WUE、Tr和Gs分别比F0显著降低6.02%、12.76%、8.97%、13.67%和17.20%,仅其Ls与F0无显著差异;与F1处理相比,F2处理黄檗幼苗的SPAD值显著增加,而其Pn、WUE和Ls则分别显著降低20.38%、13.39%和8.35%,其余指标无显著变化。黄檗幼苗叶片光合气体交换参数和叶绿素含量随着氮添加水平增加而下降,且重度氮添加的抑制效应大多达到显著水平。
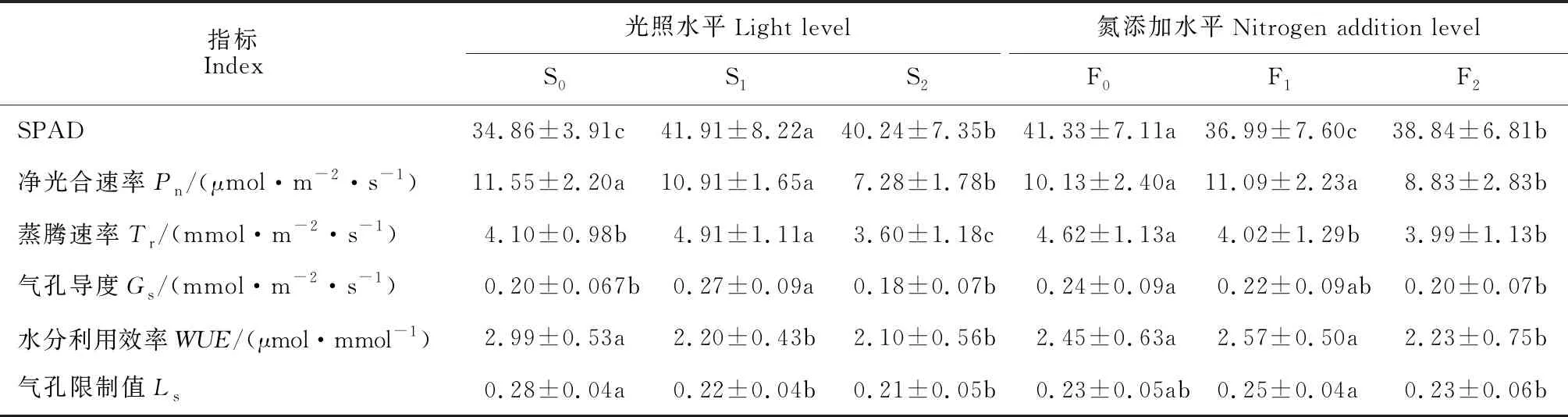
表2 遮光和氮添加下黄檗幼苗相对叶绿素含量和气体交换参数的变化
2.1.3 光氮交互效应由表3可知,黄檗幼苗叶片的Pn在各光氮组合处理间均无显著差异。黄檗幼苗叶片的其他参数在各光氮组合处理间均存在显著性差异。其中,SPAD值与其他气体交换参数表现不同,以S1F0、S2F0和S1F2处理组合显著较高,而以全光照(S0)处理组合显著最低。气体交换参数Tr和Gs以光照S1处理组合明显较高,而以S2F1、S0F2、S2F2明显较低;WUE和Ls均以S0F2显著最高,以S0F0、S0F1次之,而以S2F2和S2F0处理组合显著最低。
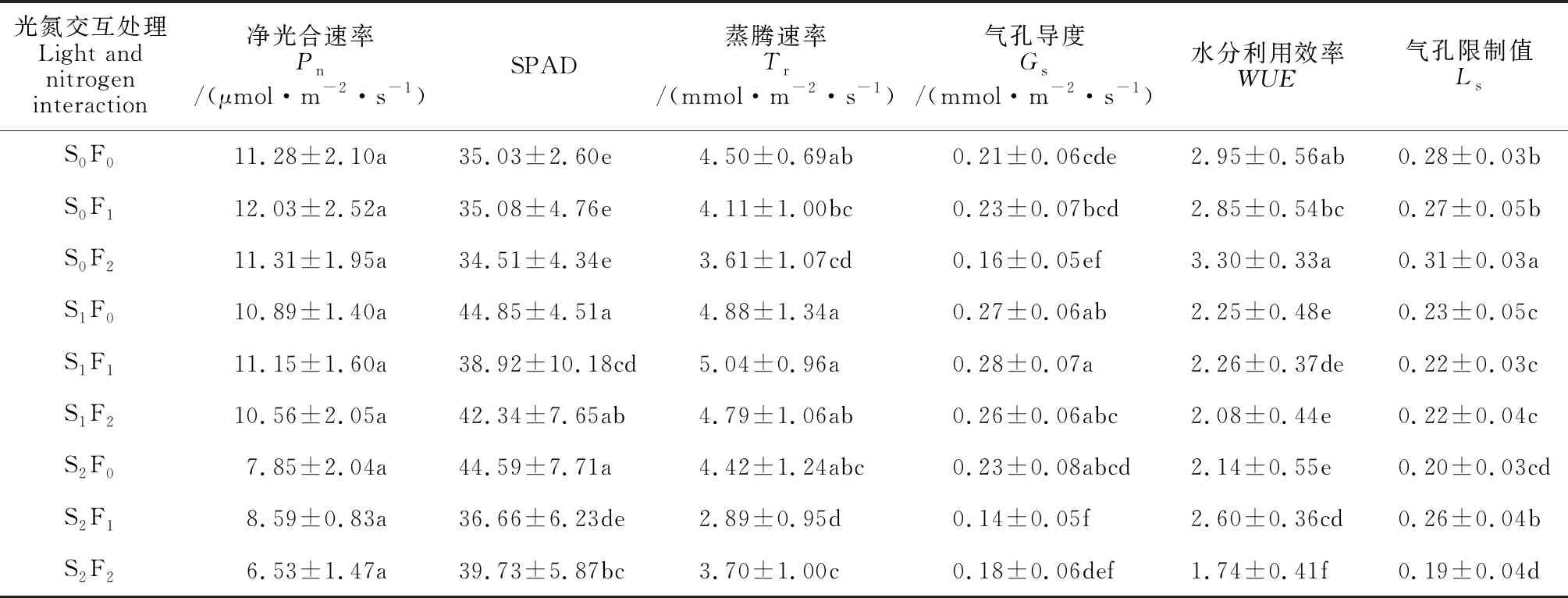
表3 光氮交互下黄檗幼苗相对叶绿素含量和气体交换参数的变化
2.2 光氮交互对黄檗幼苗叶片化学计量特征的影响
黄檗幼苗叶片N含量、C含量、C∶N及瞬时光合氮利用效率(PNUE)的方差分析结果(表1)显示,遮光、氮添加及其交互作用均对黄檗幼苗叶片的全N含量、全C含量、C∶N及PNUE产生了显著(P<0.05)影响,且单因素对黄檗幼苗碳氮化学计量特征的遮光F值(F光)远大于氮添加F值(F氮),说明光照是影响黄檗幼苗叶片化学计量特征的主要因素。
2.2.1 遮光效应由表4可知,随遮光水平增加,黄檗幼苗叶片的N和C含量均呈现出先降低后升高的趋势,而其C∶N和PNUE则呈现先增加后陡降的趋势,且它们在光照处理间均存在显著差异。其中,S1处理叶片N和C含量分别比S0显著降低18.32%和4.41%,同时也分别比相应的S2处理显著降低25.28%和3.03%;S2处理叶片的N含量比S0显著升高9.32%,其C含量则比S0显著降低1.41%;黄檗S1处理幼苗叶片的C∶N和PNUE分别比S0对照显著升高16.50%和16.54%,而S2处理的C∶N和PNUE分别比S0显著降低10.27%和27.65%,也分别比S1处理显著降低22.98%和37.92%。可见,随遮光程度从轻到重,光资源受到限制,黄檗幼苗通过调节体内养分利用率来应对不利光环境。
2.2.2 氮添加效应随着氮添加水平的增大,黄檗幼苗叶片的N和C含量均先增加后降低,C∶N逐渐增加,PNUE逐渐降低(表4)。其中,F1处理黄檗幼苗叶片N和C含量分别比F0对照显著提高4.17%和5.51%,同时也显著高于相应的F2处理,而F2处理的N含量比F0显著降低3.53%,其C含量与F0相近;F2处理黄檗幼苗叶片的C∶N分别是F1处理和F0对照的1.06和1.03倍,其叶片的PNUE则分别是F1处理和F0对照的74.82%和82.68%。可见,轻度氮添加使黄檗幼苗叶片的N和C含量显著增加,重度氮氮添加使黄檗幼苗叶片的C∶N显著增加,PNUE显著降低。
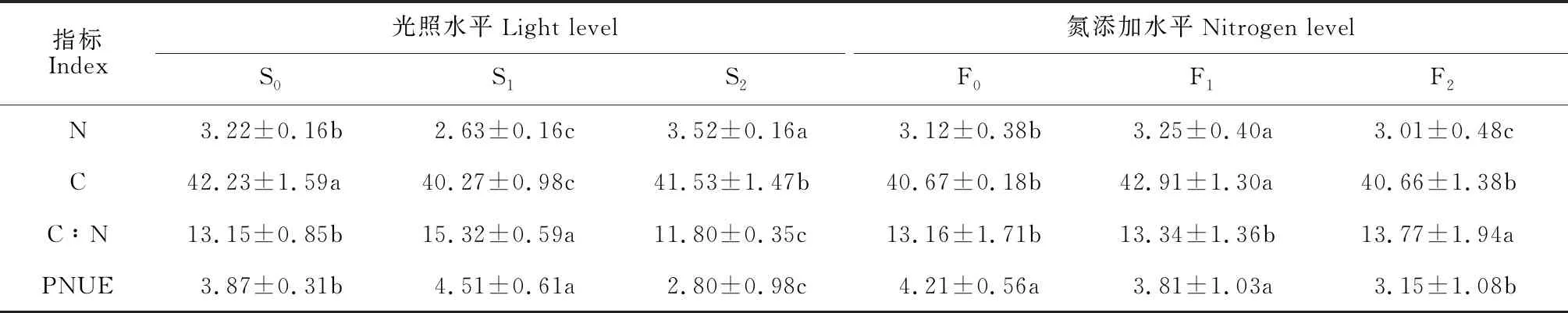
表4 遮光和氮添加下黄檗幼苗叶片化学计量特征的变化
2.2.3 光氮交互效应表5显示,黄檗幼苗叶片N含量在光氮组合处理S2F1、S2F2显著较高,而在组合处理S1F0、S1F1、S1F2显著较低,最高的S2F1、S2F2处理分别比最低的S1F2处理显著提高了49.29%和43.41%;黄檗幼苗叶片C含量在光氮组合处理S0F1、S2F1显著较高,而在S1F2处理最低并与其余组合处理差异显著,S0F1、S2F1处理分别比S1F2处理显著增加12.46%、10.79%;黄檗幼苗叶片C∶N以光氮组合处理S1F2、S1F0、S1F1显著较高,而以组合处理S2F0、S2F1、S2F2显著降低,最高的S1F2处理比最低的S2F2处理显著提高了37.51%;黄檗幼苗叶片PNUE在光氮组合处理S1F1、S1F0显著较高,而以组合处理S2F1、S2F2显著较低,并与其余处理差异显著,最高的S1F1、S1F0处理分别是最低S2F2处理的2.75和2.68倍。可见,黄檗幼苗叶片N和C含量在重度遮光/轻度氮添加时(S2F1)具有较大值,而其叶片C∶N和片PNUE在轻度遮光/无氮添加(S1F0)时具有较大值。
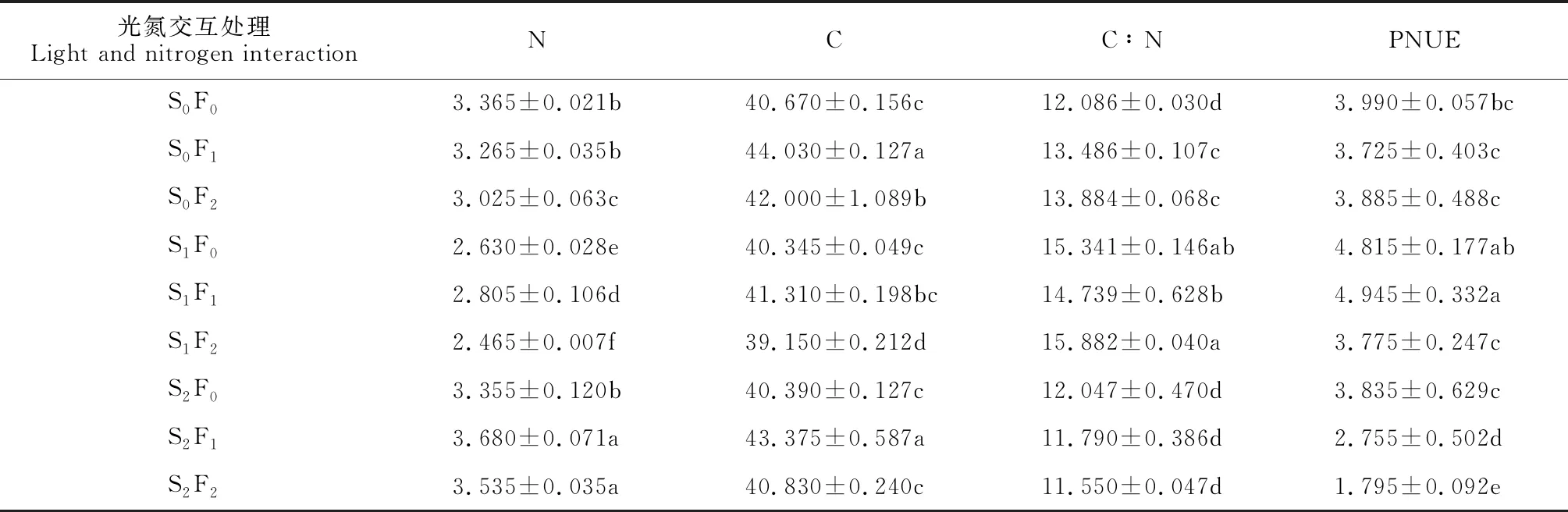
表5 光氮交互下黄檗幼苗叶片化学计量特征的变化
2.3 光氮交互对黄檗幼苗生理及化学计量特征影响效应的综合评价
采用隶属函数法综合评价各光氮复合作用对黄檗幼苗叶片生理特性及化学计量特征影响。由表6可知,光氮处理组合S1F0、S0F1和S1F1隶属函数平均值较高,介于0.631~0.652之间,为高效型;光氮处理组合 S0F2、S0F0和S1F2隶属函数平均值次之,介于0.533~0.559之间,属于中效型;而光氮处理组合S2F0、S2F1和S2F2,隶属函数平均值介于0.239~0.466之间,属于低效型。黄檗幼苗叶片生理特性及化学计量元素对光氮复合处理的隶属函数综合评价总均值为0.518,属于中效型。综上,全光照(S0)和轻度遮光(S1)条件下3种氮素水平均较适宜黄檗幼苗生理生长,其中以轻度遮光时S1F0和S1F1及全光照时S0F1为最佳组合。重度遮光条件下3种氮素水平处理黄檗幼苗叶片生理特性及化学计量特征表现均最差,不适宜黄檗幼苗生长。
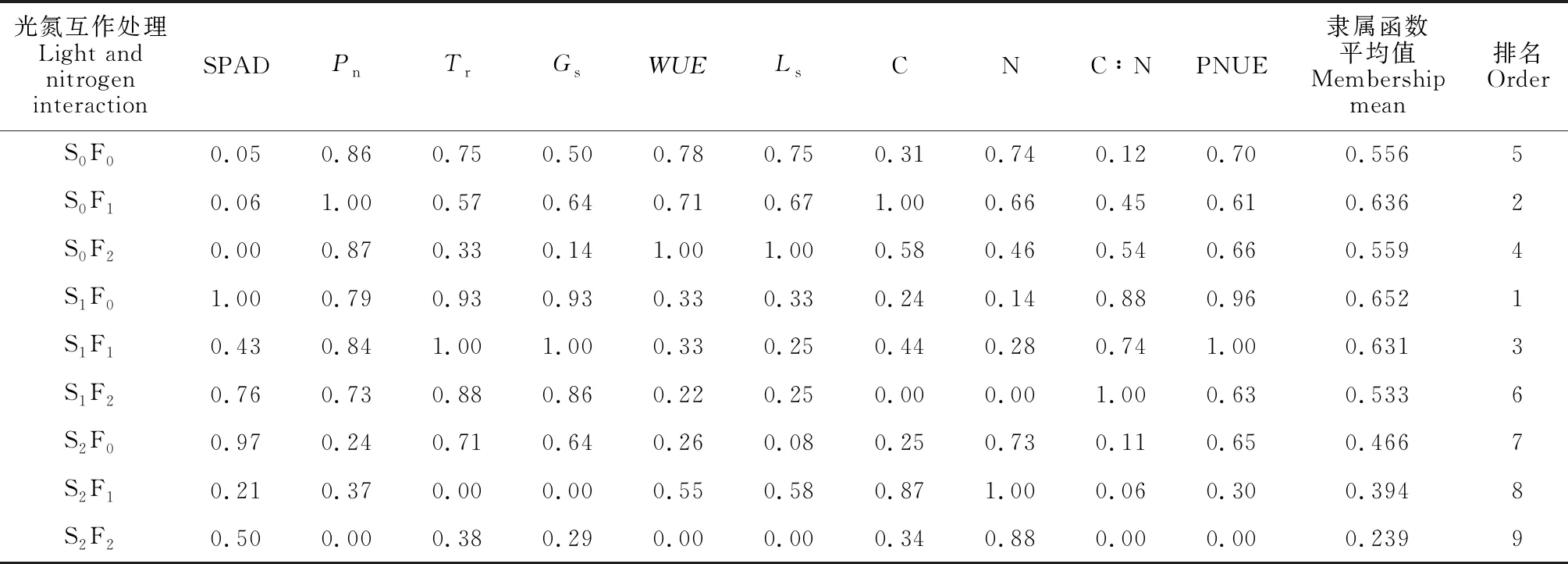
表6 光氮交互对黄檗幼苗生理及化学计量特征的综合效应评价
3 讨 论
3.1 黄檗幼苗光照需求及其可塑性
光照强弱是影响植物生理特性的主导因素,在森林更新过程中光照强度对林木幼苗的生长发育乃至成活影响更甚。本研究表明黄檗幼苗受到轻度遮光(透光率为21.4%)时能够通过提高叶片叶绿素相对含量(SPAD),降低水分利用效率(WUE)和气孔限制值(Ls),增强幼苗光合能力来适应弱光环境,表明3年生黄檗幼苗对遮光有一定的调节能力。另有研究发现林下黄檗幼苗能忍耐一定程度的遮阴,且能完成正常更新[24-26],可见适度遮阴能有效改善植物体光合特性[7]。有关研究表明遮阴显著降低黄檗叶片的光合速率[21];弱光下黄檗幼苗通过增加叶片叶绿素含量,能有效利用弱光,积累有机物质,且其光能利用率高于全光环境[27,30-31],能适应更宽的光强范围[32],提高在多变光环境条件下的适应性。本研究结果与前人相关报道一致。
3.2 氮添加对黄檗幼苗生理特性的影响
植物体氮含量是影响植物生长的重要因子之一。本研究发现在适度施氮水平下,黄檗幼苗通过调节气孔导度和蒸腾速率,增加水分利用率及气孔限制值,以适应氮添加,提高净光合速率。但过量施氮则抑制黄檗幼苗叶片气孔的开张,降低气孔导度、蒸腾速率和水分输送效率,抑制其光合作用,导致净光合速率下降,光合能力降低,影响其正常生长。相关研究表明氮添加整体上可促进植物的光合作用,且随施氮水平增加,植物光合速率先增加后降低,即随施氮量增加,叶片氮含量增加,净光合速率显著增加,当氮添加程度超过一定阈值,植物体水分被用于调节水分利用率,抑制氮代谢水平,光合效率受到限制,这是其适应氮添加的生态策略[33-35]。关于黄檗、长白落叶松(Larixolgensis)、考来木(Correacarmen)及羊草(Leymuschinensis)的相关研究也得出相似结论[35-37]。
3.3 光氮互作条件下黄檗幼苗的生理生态适应机制
光照条件和氮肥对植物的生长至关重要,其中任一因素的变化都可直接影响叶片养分特征。轻度遮光时黄檗幼苗通过提高氮养分利用效率,降低叶片氮含量,使PNUE和C∶N显著升高。同时在光资源轻度受限时也可通过提高氮吸收利用效率有效补偿碳损失,致使光合作用较全光照下略有降低,但二者并无显著差异。因此遮阴条件下适量氮供应最有利于其生理生长[25]。本研究中重度遮光时,黄檗幼苗叶片C、N含量同时增加,但其PNUE和C∶N显著降低,表明光资源严重不足时,幼苗的养分利用补偿机制受损,使得净光合速率显著降低。有关木荷(Schimasuperba)、玉米(Zeamays)、烤烟和高山杜鹃(Rhododendronlapponicum)相关研究也得出类似的结论[38-40]。
本研究中,光、氮单因素对黄檗幼苗生理及碳氮化学计量特征的分析显示,光照是比氮素更重要的影响黄檗幼苗生理特性的因素。隶属函数综合评价表明,轻度遮光、无氮添加(S1F0)和轻度氮添加(S1F1)及全光照、轻度氮添加(S0F1)为适宜黄檗幼苗生长的最佳光氮组合,且S0F1处理的黄檗幼苗叶片C含量和净光合速率均最大。光氮互作对闽楠(Phoebebournei)幼苗光合生理特性的影响研究得到类似结论[7]。在全光照条件下,黄檗幼苗碳代谢及光合速率较快,受轻度氮添加时叶片气孔打开,降低蒸腾速率,保证胞间CO2浓度,降低水分利用速率,导致叶片N含量降低,光合速率增加,利于碳累积;轻度氮添加下随着氮添加水平提高,幼苗气孔导度开始降低,蒸腾速率继续下降,水分利用效率和气孔限制值增加,叶片N、C含量降低,叶片氮除主要用于自身氮代谢功能消耗外[25],分配给光合作用的氮较少,导致PNUE和C∶N增加,光合速率降低。光照充足时,黄檗幼苗光合生理作用旺盛,在适量氮添加时有利于体内养分累积及光能利用,但当氮过量时(重度氮添加)反而不利于其幼苗的碳积累[41]。
黄檗幼苗叶片SPAD值轻度遮光下升高,在同时受到轻度氮添加时较对照(F0)降低,但在同时受到重度氮添加下反而高于对照,这与SPAD值与叶片氮含量之间的正相关结果[42]不一致。国外对3种观赏木本植物的研究表明,荚蒾属(Viburnum)树种在SPAD值与叶片氮含量相关性上区别于其他树种,可能是单位面积干叶重(LMA)变异较大,叶片单位重量氮含量、单位面积氮含量与SPAD之间相关性受LMA转换因子影响导致相关性有别于其他物种[43]。LMA功能性状指标的变异在杨树叶片发育表型可塑性中扮演重要作用[44-45]。未来将通过对LMA、比叶面积等相关光合生理指标进行进一步研究,揭示黄檗幼苗叶片表型可塑性与光氮互作的机理。
综述所述,黄檗幼苗叶片生理特性及化学计量特征对光氮交互处理响应,经隶属函数综合评价属于中效型,轻度遮光时无氮添加(S1F0)和轻度氮添加(S1F1)及全光照时轻度氮添加(S0F1)为适于幼苗生长的最佳光氮组合,且S0F1处理黄檗幼苗叶片的C含量及净光合速率均最大,光环境是影响黄檗幼苗光合作用的主导因子。黄檗属于喜光树种[46],在不受光照限制的情况下,增施适量氮肥在一定程度上可以增加叶片相对叶绿素含量,利于生长;但光资源受限和过量增施氮肥却抑制叶绿素的合成及氮素的输送,光合作用效率降低,不利于黄檗幼苗对水分及养分资源的利用,影响其地上部分的正常生长,总体长势较弱。因此,当光照不足时,应该增施适量氮肥使植物体在保证生理代谢能力不变的前提下,减小蒸腾失水,保证水分吸收,以提高光合速率。
