甜玉米品质性状的主基因+多基因遗传分析
2022-11-25麦嘉埼梁日朗周捷成李坪遥刘鹏飞
蒋 锋,闫 艳,麦嘉埼,梁日朗,周捷成,李坪遥,刘鹏飞
(1仲恺农业工程学院农业与生物学院,广州 510225;2广州市特色作物种质资源研究与利用重点实验室,广州 510225)
0 引言
甜玉米具有营养价值高、风味独特,低脂肪高纤维和富含人体所必须的多种营养成分等综合优点[1]。果糖是甜玉米食用品质和营养品质的重要指标,其含量直接决定了甜玉米的甜度,进而影响了品质的优劣[2]。膳食纤维是除了传统的六类元素(蛋白质、脂肪、矿物质、水、碳水化合物和维生素)之外的第七类营养元素[3]。营养调查资料表明,膳食纤维能有效减少和预防冠心病、糖尿病等疾病的发生[4]。维生素C是人体必需的微量物质,在人体生命活动方面起着重要的作用,可以增强人体免疫力,预防人体动脉硬化,并且参与人体胶原蛋白的合成[5],还可以预防癌症等重大疾病[6]。随着人们生活质量的改善,开始注重日常饮食的营养结构,甜玉米越来越受到消费者的青睐,同时对其品质的要求也越来越高[7]。
盖钧镒[8]提出的植物数量性状主基因+多基因混合遗传模型分析方法可以鉴别主基因,检测出多基因效应、基因间的互作以及基因与环境间的互作,并给出目标性状的最佳遗传模型及遗传参数[9]。这一分析方法已经在多个玉米品质性状的遗传规律分析上得到应用。刘鹏飞等[10]对甜玉米果皮厚度进行分析,得出果皮厚度符合1对加性主基因+加性-显性多基因混合遗传。宋轶群等[11]、栗亚静等[12]研究表明玉米淀粉含量是由2对加性-显性-上位性+加性-显性多基因控制,毕成龙[13]、李颖[14]、包和平等[15]研究表明玉米淀粉含量由1对加性主基因+加性-显性多基因控制。李颖[14]对高油玉米蛋白含量遗传模型进行分析,得出蛋白含量符合2对主基因加性-显性-上位性+加性-显性多基因混合遗传模型。包和平等[16]对爆裂玉米蛋白质含量进行遗传模型分析,得出蛋白含量符合1对主基因+加性-显性多基因混合遗传模型;进茜宁等[17]研究表明玉米含油量符合1对加性主基因+加性-显性多基因混合遗传模型,李颖[14]和高树仁等[18]研究表明玉米含油量符合1对加性主基因-显性主基因+加性-显性-上位性多基因混合遗传模型。
目前,关于甜玉米品质性状的遗传研究,主要集中于淀粉含量、蛋白含量、含油量等,对于甜玉米果糖含量、膳食纤维含量、维生素C含量的遗传模型分析鲜有报道。本试验用主基因+多基因混合遗传模型分析方法对甜玉米果糖含量、膳食纤维含量、维生素C含量进行遗传模型分析,得出最适遗传模型,分析相关遗传参数,以期提高甜玉米分子育种效率,减少育种选择的盲目性。试验结果有望为甜玉米品质分子育种工作及甜玉米品质的遗传改良和选择提供一定的理论依据。
1 材料与方法
1.1 试验材料
试验亲本材料为果糖含量、膳食纤维含量和维生素C含量有显著差异的甜玉米自交系T77(高值亲本)和T15(低值亲本)。其中T77(高值亲本)果糖含量为(1.89±0.04)%、膳食纤维含量为(4.18±0.13)%、维生素C含量为(45.76±0.37)mg/100g,T15(低值亲本)果糖含量为(0.63±0.03)%、膳食纤维含量为(2.62±0.11)%、维生素C含量为(27.91±0.23)mg/100g。这2个自交系分别由台湾甜玉米品种‘华珍’和泰国甜玉米品种‘先甜5号’为基础材料,经8代自交、鉴定选育而成。
1.2 田间试验
2019年3—6月在仲恺农业工程学院广州市番禺区钟村教学科研基地,使用T15(低值亲本)和T77(高值亲本)分别作为父母本进行杂交。到2019年9—12月,对两亲本杂交产生的F1单株进行套袋严格自交,得到F2种子。2020年3—6月,种植2个亲本各20株及来自同一F1果穗的216个F2单株,选取200株发育良好的F2植株进行表型测定及遗传模型分析。
1.3 数据测定
授粉后22天,取各单株果穗进行品质检测。
果糖含量采用李志英等[19]的紫外分光光度法测定。膳食纤维的提取与含量测定参考孙艳[20]的方法。维生素C含量参考曾宪录等[21]按GB5009.86—2016采用2,6-二氯靛酚滴定法测定。
1.4 数据处理与分析
试验数据采用Microsoft Excel 2010进行整理,并对T77×T15组合的F2群体的果糖含量、膳食纤维含量和维生素C含量值进行频数分布统计。参照盖钧镒[8]、王建康等[22]、章元明等[23]的分析方法,利用南京农业大学提供的主基因+多基因遗传分析软件,计算11种模型的极大似然函数值和AIC值确定各个模型的符合程度。通过AIC值筛选备选模型,再进行适合性检验,确定最佳遗传模型,然后应用最小二乘法估算遗传参数。
2 结果与分析
2.1 F2群体品质性状的统计分析
对200个F2单株果穗的果糖含量、膳食纤维含量及维生素C含量进行数据统计,分析平均值、标准差、变幅大小及变异系数(表1),并作频数分布图(图1、2、3)。从F2分布参数和频数分布图来看,果糖含量性状的偏度为-0.12,未显著偏离正态分布,膳食纤维含量和维生素C含量的偏度分别为1.15和-1.10,其绝对值大于1,显著偏离正态分布。
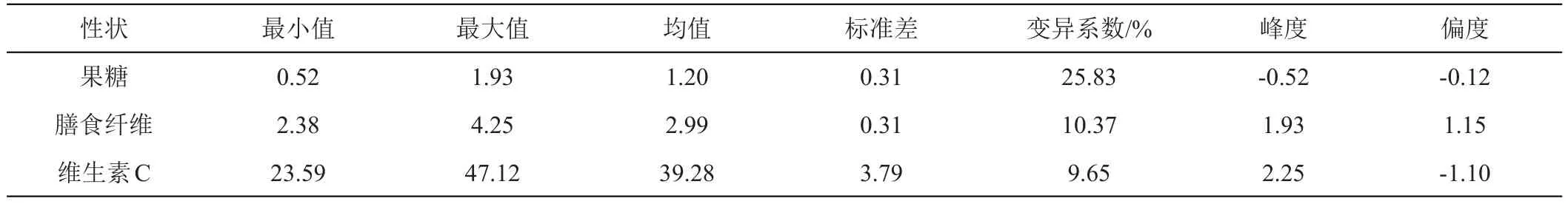
表1 200个F2世代描述统计分析
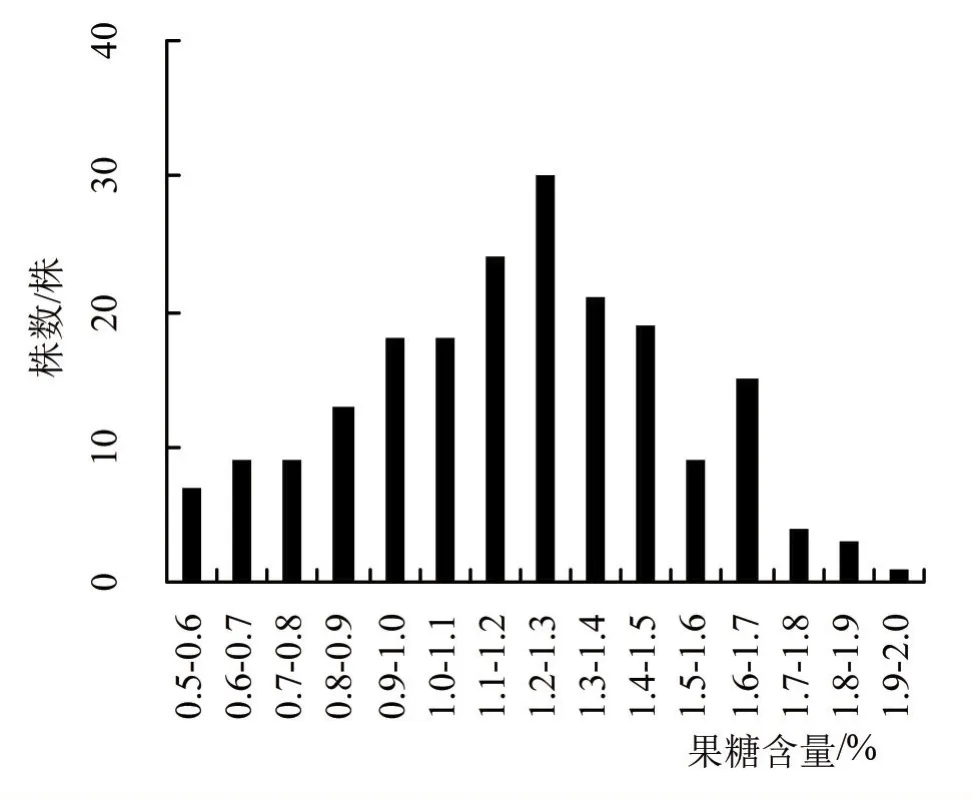
图1 甜玉米T77×T15组合F2代果糖含量的频数分布
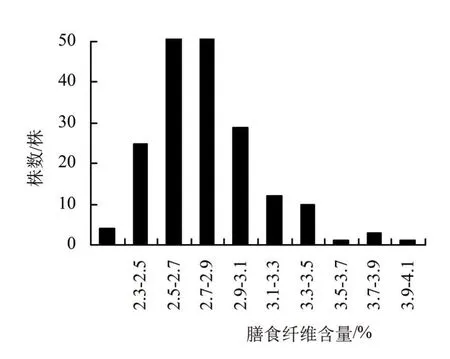
图2 甜玉米T77×T15组合F2代膳食纤维含量的频数分布

图3 甜玉米T77×T15组合F2代维生素C含量的频数分布
2.2 甜玉米品质性状备选遗传模型分析
应用主基因+多基因遗传分析软件计算出11种遗传模型的AIC值和极大似然值(表2)。初步筛选出AIC值较小的模型作为备选模型,进一步进行适合性检验,确定最佳的遗传模型。果糖含量A-1的AIC值最小,且B-2和B-3的AIC值与之相近,可作为备选模型;膳食纤维含量B-1的AIC值最小,且A-1和A-4的AIC值与之相近,可作为备选模型;维生素C含量B-1的AIC值最小,且A-1、A-3和B-5的AIC值与之相近,可作为备选模型。

表2 11种遗传模型的极大似然值和AIC值
2.3 备选模型的适合性检验
根据U12、U22、U32(均匀性检验)、nW2(Smirnov检验)、Dn(Kolmogorov检验),对果糖含量、膳食纤维含量、维生素C含量3个品质性状的备选模型进行适合性检验(表3),结合AIC值大小选出3个品质性状的最适遗传模型。
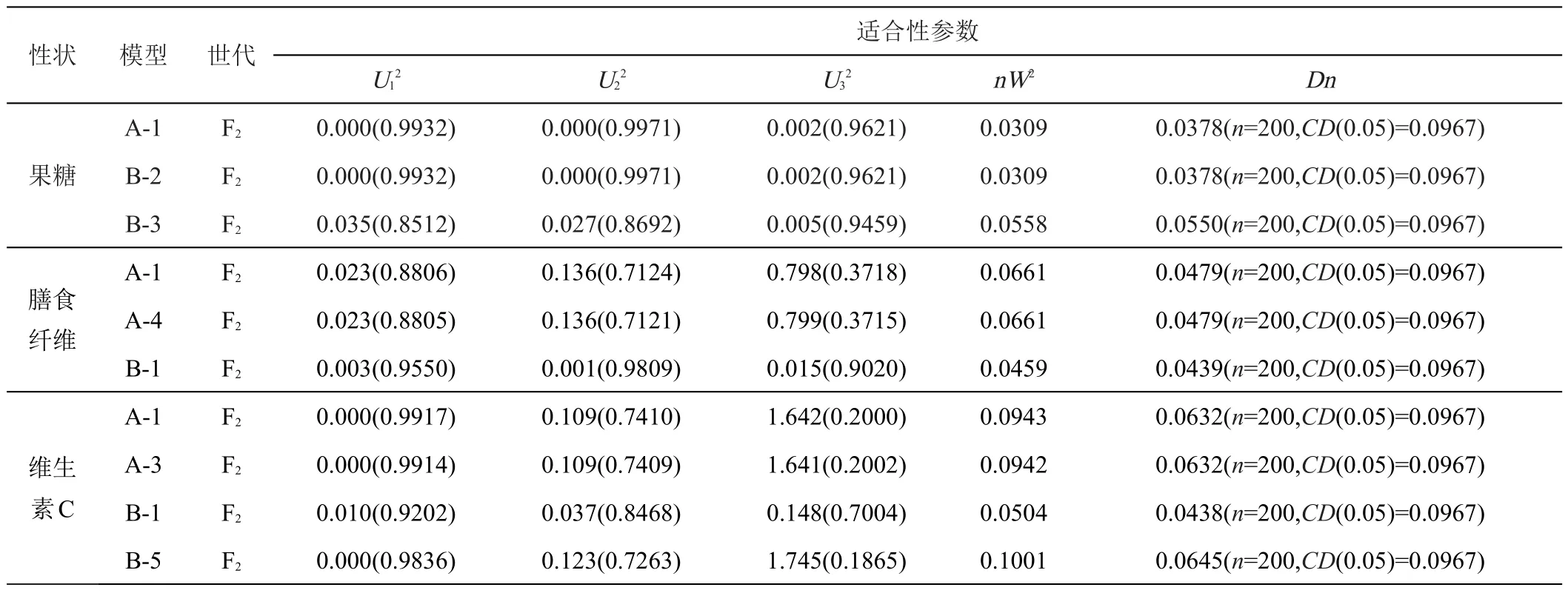
表3 备选遗传模型的适合性检验
果糖含量 3 个备选模型的U12、U22、U32、nW2和Dn值均未达到显著水平,模型A-1的AIC值最小,因此选A-1为果糖含量的最适遗传模型。说明果糖含量的遗传模型是1对主基因控制的加性-显性混合遗传模型。
膳食纤维含量3个备选模型的U12、U22、U32、nW2和Dn值均未达到显著水平,模型B-1的AIC值最小,因此选B-1为膳食纤维含量的最适遗传模型。说明膳食纤维含量的遗传模型是2对主基因控制的加性-显性-上位性混合遗传模型。
维生素C含量4个备选模型的U12、U22、U32、nW2和Dn值均未达到显著水平,模型B-1的AIC值最小,因此选B-1为维生素C含量的最适遗传模型。说明维生素C含量的遗传模型是2对主基因控制的加性-显性-上位性混合遗传模型。
2.4 甜玉米品质性状最适模型的成分分布
甜玉米果糖含量最佳遗传模型为A-1,遗传表现为加性-显性遗传,F2群体受1对主基因控制,3个主基因型成分按1:2:1分布(表4);甜玉米膳食纤维含量最佳遗传模型为B-1,遗传表现为加性-显性-上位性遗传,F2群体受2对主基因控制,9个主基因型成分按1:2:1:2:4:2:1:2:1分布(表5);甜玉米维生素C含量最佳遗传模型为B-1,遗传表现为加性-显性-上位性遗传,F2群体受2对主基因控制,9个主基因型成分按1:2:1:2:4:2:1:2:1分布(表5)。

表4 最适模型下果糖含量的成分分布的均值、权重和标准差
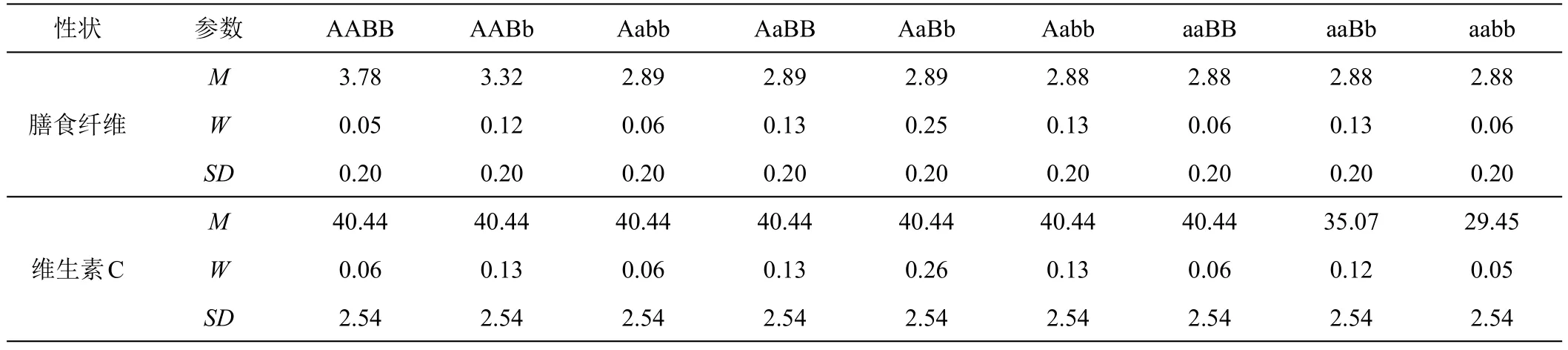
表5 最适模型下膳食纤维含量和维生素C含量的成分分布的均值、权重和标准差
2.5 甜玉米品质性状遗传模型的参数估计
根据已确定的3个品质性状的最适遗传模型,应用最小二乘法[24],对F2群体3个品质性状进行遗传参数估计(表6)。
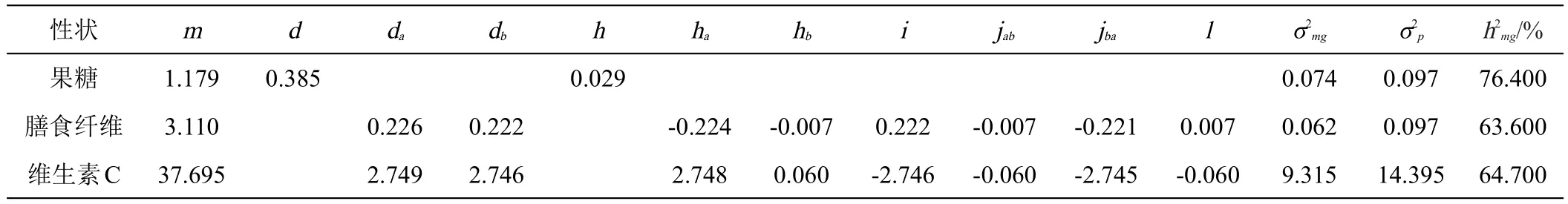
表6 甜玉米品质性状遗传参数估计值
甜玉米果糖含量的遗传模型符合A-1,F2分离群体受1对主基因控制,控制甜玉米果糖含量的1对主基因的加性效应值为0.385,显性效应值为0.029,说明控制甜玉米果糖含量的主基因以加性效应为主,主基因遗传率为76.4%。
甜玉米膳食纤维含量的遗传模型符合B-1,F2分离群体受2对主基因控制,控制甜玉米膳食纤维含量的2对主基因的加性效应值分别为0.226和0.222,2对主基因的显性效应值均为负值,分别为-0.224和-0.007,表明低膳食纤维含量相对于高膳食纤维含量表现为显性。2对主基因加性效应之和显著大于显性效应之和(|da+db|>|ha+hb|),甜玉米膳食纤维含量的遗传以2对主基因的加性效应为主。第1对主基因的显性度|ha/da|=0.9912,表明第1对主基因表现为部分显性;第2对主基因的显性度|hb/db|=0.0315,表明第2对主基因表现为加性效应。2对主基因之间存在多种互作,其中加性互作效应值i为0.222,显性互作效应值l为0.007,加性和显性互作效应值jab和jba分别-0.007和-0.221,表明基因间的互作会对膳食纤维含量产生影响。从二阶遗传参数来看,2对主基因遗传方差为0.062,主基因遗传率为63.6%。
甜玉米维生素C含量的遗传模型符合B-1,F2分离群体受2对主基因控制,控制甜玉米维生素C含量的2对主基因的加性效应值分别为2.749和2.746,2对主基因的显性效应值分别为2.748和0.06,加性效应之和显著大于显性效应之和(|da+db|>|ha+hb|),表明甜玉米维生素C含量的遗传以2对主基因的加性效应为主。第1对主基因的显性度|ha/da|=0.999,表明第1对主基因表现为部分显性;第2对主基因的显性度|hb/db|=0.022,表明第2对主基因表现为加性效应。2对主基因之间存在多种互作,其中加性互作效应值i为-2.746,显性互作效应值l为-0.06,加性和显性互作效应值jab和jba分别-0.006和-2.745,表明基因间的互作会对维生素C含量产生影响。从二阶遗传参数来看,2对主基因遗传方差为9.315,主基因遗传率为64.7%。
3 讨论
植物数量性状遗传体系“主基因+多基因遗传模型”,这一分析方法已在花生[25]、小黑麦[26]、玉米[27]等多种作物的数量性状遗传规律分析上得到应用。本试验运用主基因+多基因分析的方法,以果糖含量、膳食纤维含量和维生素C含量差异较大的甜玉米自交系配制杂交组合T77(高值亲本)×T15(低值亲本),对果糖含量、膳食纤维含量和维生素C含量进行遗传模型分析。结果表明:甜玉米果糖含量最佳遗传模型为A-1模型,为加性-显性遗传模型,受1对主基因控制遗传,主基因遗传率为76.4%。蒋锋等[28]以8个超甜玉米自交系为亲本,利用Hayman法分析了超甜玉米果糖含量的杂种优势及遗传模型。结果表明,果糖性状的遗传受加性基因和非加性基因的共同控制,狭义遗传力为10.92%,广义遗传力为94.15%。单明珠等[29]研究表明甜玉米含糖量的遗传符合加性-显性模型。
本研究中甜玉米膳食纤维含量和维生素C含量的最佳遗传模型均一致,符合B-1模型,为加性-显性-上位性遗传模型,受2对主基因控制遗传,主基因遗传率分别为63.6%和64.7%。胡新洲等[30]以8个超甜玉米自交系为亲本,按照Griffing完全双列杂交方法分析表明,可溶性膳食纤维和不溶性膳食纤维的遗传受加性基因和非加性基因的共同控制。蒋锋等[31]对甜玉米膳食纤维含量进行QTL定位,在第4,第9染色体上检测与膳食纤维相关的QTL,共4个。分别可解释11.17%、9.13%、16.40%、18.42%的表型贡献率,共解释55.12%的表型贡献率。李冉冉等[32]研究了不同授粉方式下玉米纤维含量的遗传规律,结果表明,在开放授粉条件下纤维含量的遗传力为87.27%,自交授粉条件下,纤维含量的遗传力为80.68%。
综上所述,本试验的结果与前人的结论相似或一致,下一步还需要利用不同材料组合,进行多世代、多年份、多地区的种植试验,对试验结果进行补充和验证。本试验的结果表明,甜玉米果糖含量、膳食纤维含量及维生素C含量的遗传率较高,在育种实践中,对这3个性状的遗传改良和选择可在早期世代进行,可采用聚合回交或轮回选择的方法来积累微效基因以提高育种效率。同时,本试验结果得到的为目标性状的遗传模型,不能确定控制性状的基因的位置,后续研究需要借助分子标记技术对相关基因进行定位和挖掘,结合遗传模型,为甜玉米品质的遗传改良和选择提供一定的理论依据。
4 结论
应用主基因+多基因混合遗传模型分析方法,对品质性状差异较大的甜玉米杂交组合T77(高值亲本)×T15(低值亲本)的F2代品质性状进行遗传分析。结果表明,甜玉米果糖含量的最佳遗传模型为A-1,为1对加性-显性主基因遗传模型,主基因遗传率为76.4%,膳食纤维含量和维生素C含量的最佳遗传模型为B-1,为2对加性-显性-上位性多基因混合遗传模型,主基因遗传率分别为63.6%和64.7%。
