读心的比较研究:非人灵长类与人类在心理理论上的异同点及其解释*
2022-11-15殷融
殷 融
读心的比较研究:非人灵长类与人类在心理理论上的异同点及其解释*
殷 融
(潍坊学院教师教育学院, 山东 潍坊 261061)
利用非人灵长类开展比较研究是探索心理理论本质、起源和进化动因的重要途经。大量实证研究显示非人灵长类可以表现出一些基础心理理论成分, 包括联合注意、意图理解和一级观点采择, 这表明心理理论在灵长类动物的进化谱系中具有进化连续性。然而, 非人灵长类无法像人类一样达成二级观点采择和错误信念理解, 潜心智化假说、替代表征假说、最小限度心理理论假说和意识关系假说试图对此差异进行解释。尽管视角与具体内容不同, 但这些假说都强调, 非人灵长类无法对其他个体关于外部世界的心理信念形成表征, 信念表征是非人灵长类与人类读心能力的重要分界点。未来研究需要重点澄清共享意图、语言机制以及社会生态环境因素与心理理论产生的相互影响作用。
非人灵长类, 心理理论, 联合注意, 观点采择, 错误信念
1 引言
心理理论(theory of mind)也被称为读心(mind-reading)机制或心理状态归因(mental state attribution), 它指个体对他人心理状态进行推测并据此对他人行为做出因果解释的能力(Premack & Woodruff, 1978)。只有具备心理理论, 个体才能顺利完成交流、教学、合作或欺骗等活动, 因此这种心智能力在人类社会化生活中扮演重要角色。近30年来, 社会心理学、发展心理学、生理心理学和认知神经科学等心理学诸多领域都围绕该主题进行了大量研究, 涉及到包括婴幼儿心理理论发展特征、心理理论的研究范式、影响心理理论的环境因素、心理理论与自闭症的关系以及心理理论的神经机制等问题, 其中许多问题国内研究者已进行过相关介绍与讨论(陈丽娟, 徐晓东, 2020; 霍超等, 2021; 皮忠玲, 莫书亮, 2013; 钱淼, 傅根跃, 2014; 隋雪等, 2021)。
利用现存非人灵长类(non-human primates)开展心理理论比较研究, 心理学家可以回答一些仅以人类作为研究对象时难以解决的问题, 如心理理论与其他心智成分的进化关系、心理理论的必要认知准备以及人类心理理论的特异性等, 并在此基础上更好地认识心理理论的本质、起源和进化动因(莫书亮等, 2004)。因此, 许多关注心理理论的学者会将研究重心聚焦于非人灵长类。本文通过实证研究梳理, 总结非人灵长类在心理理论方面与人类相似及差异之处, 论述相关理论假说, 并对该领域今后重要研究方向进行展望。
2 非人灵长类心理理论的表现
非人灵长类是否也具有一定程度的心理理论?实际上, 心理理论这一概念最早就是Premack和Woodruff (1978)在研究黑猩猩的社会认知能力时首次提出的, 但在20世纪中后期, 许多研究者都曾认为心理理论是人类独有的认知特征(Heyes, 1998)。经过几十年实证研究积累, 该结论被逐渐颠覆, 这其中一个重要原因是实验范式的改进。
当考察人类特别是成人的心智能力时, 研究者可以将被试置于某一故事场景中, 直接向被试提问, 让其解释故事中主人公的行为或预测主人公之后行动。而在针对非人灵长类的实验中, 由于动物被试无法口头报告, 因此自发反应(spontaneous-response)实验模式被更普遍地采用(Horschler et al., 2020)。使用这一实验模式时, 研究者将被试置于包含另一个行动者的特定场景中, 被试作为观察者, 当行动者做出某些行为后, 研究者通过记录被试产生的自发反应, 如眼睛注视点变化、眼睛注视时间差异、注视偏好或选择倾向等, 分析被试表现是否符合心理理论特征。
从个体发展角度看, 心理理论作为一种高级社会认知能力, 存在从无到有、从简单到复杂的渐进发展过程。对于人类来说, 完整的心理理论系统包含许多子成分, 如联合注意(joint attention)、意图理解(goal understanding)、观点采择(perspective-taking)与错误信念理解(false beliefs understanding)等。这些成分复杂程度不同, 在人类个体一生中往往按照特定顺序出现(Horschler et al., 2020)。研究发现, 非人灵长类在这些子成分上具有不同表现(Krupenye, 2021)。
2.1 联合注意
联合注意指个体能够追随或吸引其他个体注意的能力(Horschler et al., 2020)。对于社会化动物来说, 个体只有先做到可以关注其他个体的注意聚焦对象, 与其他个体同时关注同一事物, 才能在此基础上理解他者的意图或想法, 进而对自身行为做出调整。因此, 联合注意被视为心理理论出现的萌芽(Bettle & Rosati, 2019)。联合注意可分为反应性联合注意和主动性联合注意, 前者表现为个体能够根据他人视线或身体姿势“捕捉”他人的关注对象, 后者则表现为个体可以利用眼神接触或动作促使他人关注自己或其他外部事物(Mundy & Newell, 2007)。研究发现, 人类婴儿最早在3个月大时已经能够跟随成年人目光注视周围物体, 9个月大的婴儿已可以灵活地通过注视或手势将他人注意引向特定目标(Rosati et al., 2016)。那么, 非人灵长类在联合注意方面具有怎样的表现?
对于灵长类动物来说, 反应性联合注意是一种无论在竞争(如根据捕食者注视方向做出及时躲避反应)还是合作(如接收关于食物来源的信息)环境下都能发挥重要生存价值的能力。反应性联合注意最主要的表现形式是注视追随(gaze- following), 目前研究表明, 大部分非人灵长类都能在某些情境下追随其他个体的目光, 同时它们在不同情境下还会对目光注视做出不同理解。更重要的是, 非人灵长类并不仅仅是对其他个体的眼神注视方向做出条件反射式被动反应, 它们会主动追踪其他个体的目光, 对其他个体的注视行为表现出极高兴趣。在动物实验中, 要检验这一假设通常使用“目标−遮挡”任务, 实验员会注视一个目标物, 由于存在遮挡, 这个目标物在动物被试视线之外, 动物如果要看到实验员看到的东西, 必须移动到实验员所在位置。实验证明, 几种类人猿和猕猴都可以做到这一点(Bettle & Rosati, 2019; MacLean & Hare, 2012)。此外除了视线方向, 非人灵长类还可以利用头部或身体朝向等线索来理解其他个体的注意力状态(以上结论相关实证研究见表1)。
与反应性联合注意相比, 主动性联合注意是一种更复杂的能力, 这种能力需要个体既能关注其他个体当前的注意状态, 也能理解如何成功将对方的注意力引导到特定事物上(Horschler et al., 2020)。研究发现, 许多非人灵长类在沟通时都会先评估交流对象的注意力方向, 如果信息接收者的状态不“达标”或没有做出恰当反馈, 非人灵长类会调整沟通策略, 它们会通过动作与声音吸引对方的关注、主动移动到对方面前或用眼神在目标物与信息接收者之间来回切换, 确定自己与交流对象都在关注同一物体(以上结论相关研究见表1)。因此, 非人灵长类也具有主动性联合注意的能力。
2.2 意图理解
意图理解指个体根据潜在目标或目的来理解其他个体行为的能力(Buttelmann et al., 2007)。这种机制在社会互动中也具有至关重要意义, 同样行为动作可能出于不同目的, 个体只有理解行动者意图, 才能预测其进一步行动。也就是说, 他既需要知觉到行动者在做什么, 又需要判断行动者为什么这么做。例如, 当一个人看到另一个人将手伸向苹果时, 他并不仅仅看到了“伸手”这个动作, 还能理解对方“想要苹果”这一意图, 进而能预测“对方会用手拿起苹果”。由于行为目的并不是可以直接观察到的, 因此意图理解意味着观察者能够将情境因素、被观察者的行为以及行动目标整合在一起, 并认知到其中的因果逻辑关系。研究发现, 人类婴儿早在出生半年后, 就能表现出意图理解能力, 例如他们会对突然中断的动作表现出诧异, 因为动作中断与他们对该动作功能的预期不符(Krupenye & Call, 2019)。
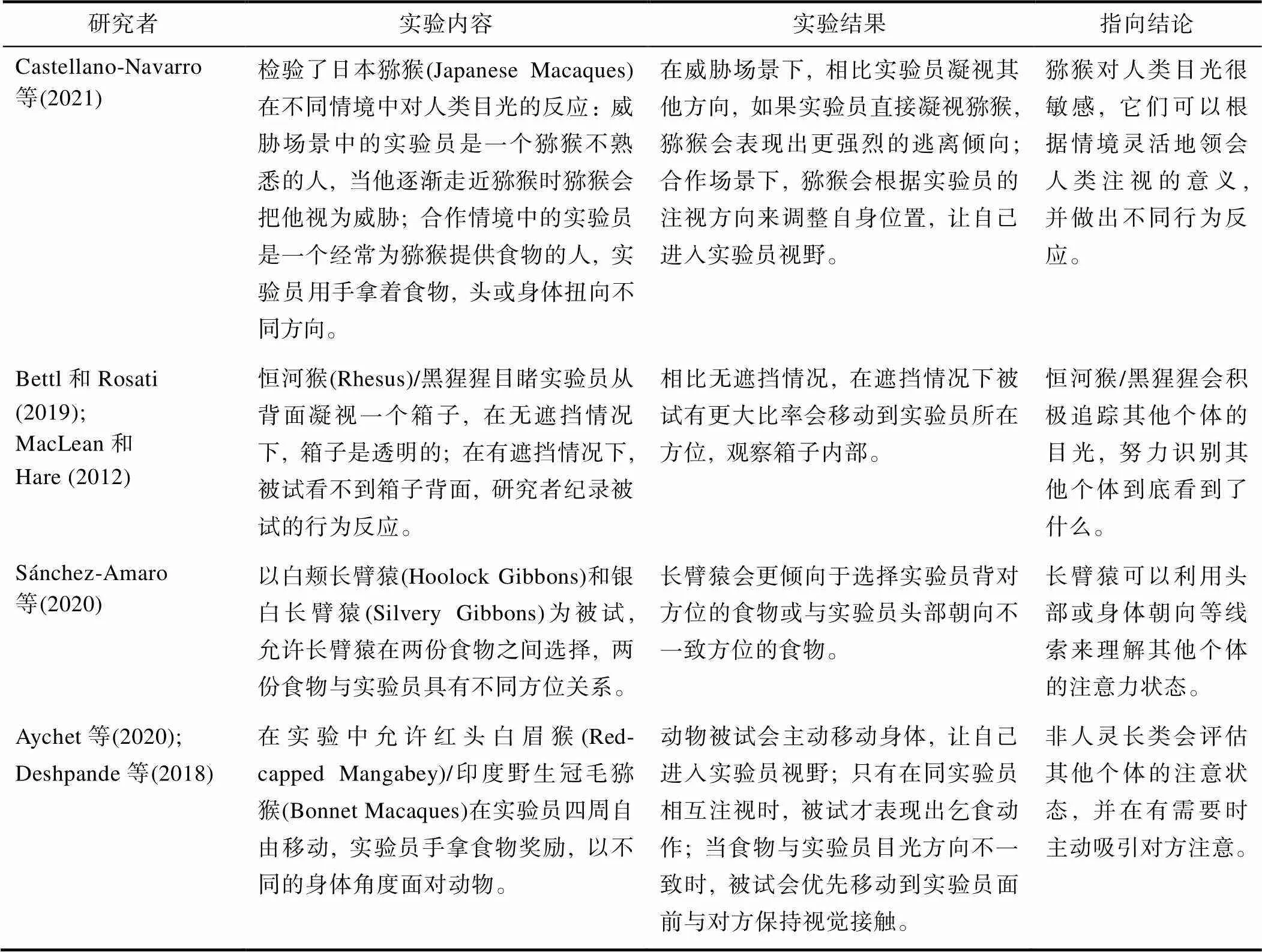
表1 非人灵长类联合注意的代表性实证研究
为检验非人灵长类的意图理解能力, 研究者探索了许多不同实验模式与实验情境。实证研究表明, 非人灵长类能够区分“有意的目标导向动作”与“无意动作”; 它们可以将背景信息与个体行为结合起来, 利用空间关系、潜在目标、过往经验、身体动作以及环境线索等信息理解他者的行为意图; 当学习其他个体的动作时, 它们能“去芜存菁”, 只模仿与行动目标有关的动作, 而不是完全复制(以上结论相关实证研究见表2)。
另外, 一些非人灵长类还能捕捉到抽象图形运动中所暗含的“目的性”。例如, 屏幕上呈现两个运动点, 当两个点的运动轨迹一前一后较为一致时, 在人类看来, 这像是一个点“想要”追踪另一个点。研究发现, 至少部分松鼠猴(Atsumi & Nagasaka, 2015)和日本猕猴(Atsumi et al., 2017)可以区分两个点是在随机自由运动还是前后“追随”, 它们可以将抽象图形的动作轨迹组织为连贯序列并赋予其行动意图。
2.3 观点采择
观点采择指从另一个个体视角看待外部世界并推断其体验的能力(MacLean & Hare, 2012)。判断其他个体是否能知觉到某事件或掌握某些信息是人类心理理论的重要表现, 该能力是许多社会行为的基础, 如隐瞒秘密、偷窃以及有针对性的提供情报等。使用“预期违背任务”作为实验范式的研究发现, 人类婴儿在10至15个月大时就开始能判断某事物是否可被其他个体所看到(Baillargeon et al., 2018; Luo, 2011), 在这类研究中, 实验员看到目标物被藏到几个遮挡物中的一个, 婴儿被试目睹了实验员的观察过程, 如果实验员去打开错误的遮挡物, 婴儿会注视更长时间, 说明他们对实验员的行为产生了疑惑, 因为实验员看到了藏匿结果却做出错误选择。
同类研究发现, 非人灵长类在看到实验员做出与其预期相违背的选择时, 也会注视更长时间; 同时, 一些非人灵长类还可以利用自身经验来推断其他个体可能具有的特殊视角; 关于非人灵长类观点采择能力最有力的证据来自于“食物竞争任务” (MacLean et al., 2013), 在这类研究中, 非人灵长类被试会基于竞争对手的视角来预测对方先获取哪些食物, 也就是说, 它们不但能判断其他个体是否了解关于周围环境的某些信息, 还能以此作为推理依据, 想象其他个体会怎样思考和行动(以上结论相关实证研究见表3)。

表2 非人灵长类意图理解的代表性实证研究
观点采择能力具有不同的复杂程度, 非人灵长类能判断出其他个体是否可感知到某事物, 这种情况被称为一级观点采择(level-1 perspective- taking)。对于人类来说, 个体还能知道同一物体在他人看来可能是不同的, 这种情况被称为二级观点采择(level-2 perspective-taking), 皮亚杰的经典三山实验即反映了该能力在幼儿身上的发展过程(Moll & Meltzoff, 2011)。事实上, 人类还可以表现出比二级观点采择更复杂、层次更多的递归式心智解读能力, 如“我明白你相信A并没真的理解B的想法”就包含了4层递归关系。一些研究者认为, 人类能够理解的心理状态嵌套层次可以达到7层(O’Grady et al., 2015)。而实验表明, 非人灵长类可能不具备二级观点采择能力, 例如在食物竞争任务中, 如果被试与竞争者对情境的观察视角不同, 此时被试就无法代入竞争者视角做出正确预测(Karg et al., 2016)。因此, 非人灵长类无法理解自己与其他个体对同一物体的观察会有差异(以上结论相关实证研究见表3)。
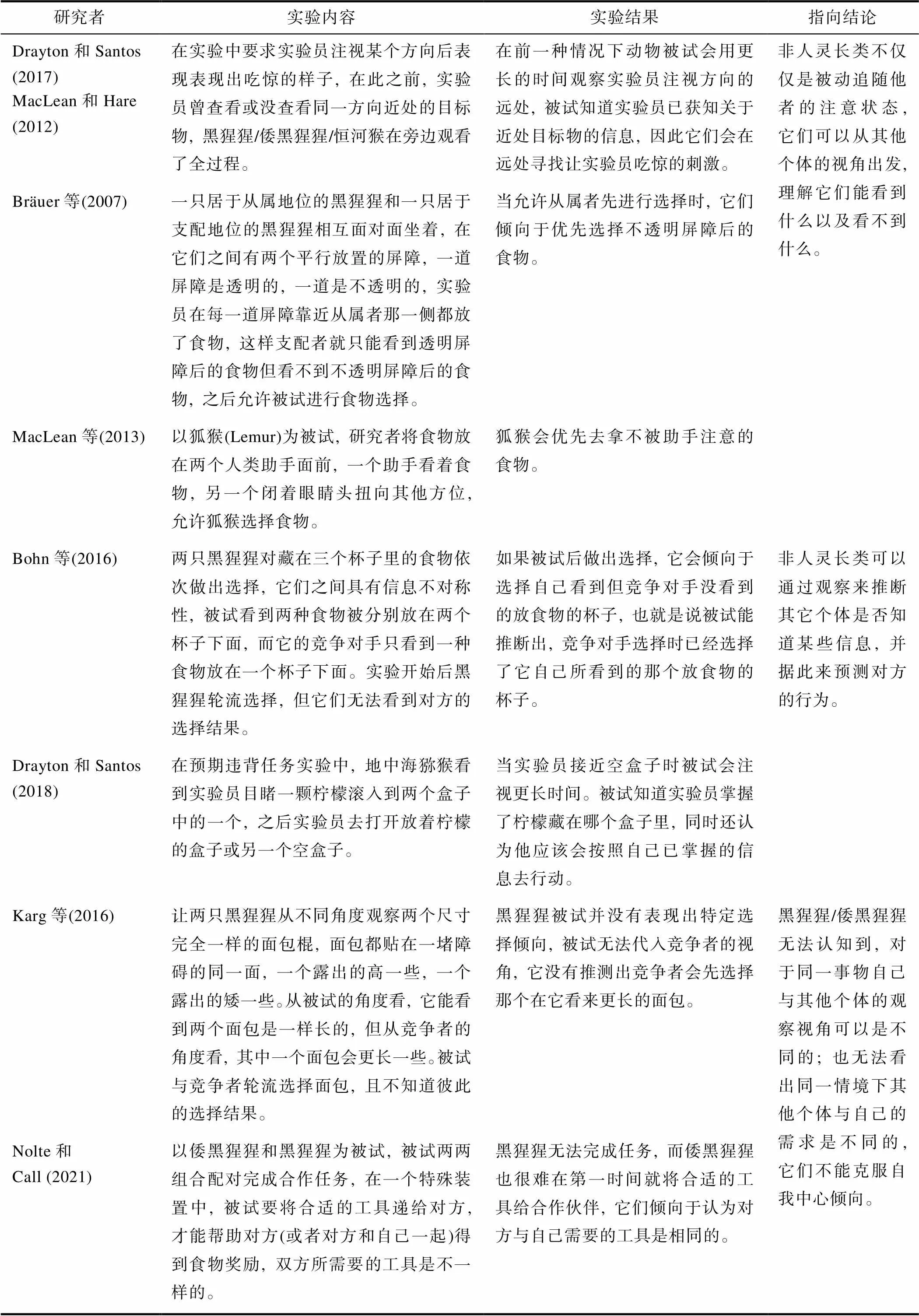
表3 非人灵长类观点采择的代表性实证研究
2.4 错误信念理解
错误信念是指个体理解其他个体对于外部世界的某种看法与现实不相符的情况(Krupenye et al., 2016)。个体要做到这一点, 需要同时认知到其他个体错误的信念以及他应该具有的正确信念, 同时还要能够区分它们的差异, 因此, 错误信念理解常被认为是成熟心理理论机制的体现, 也被认为是儿童心理理论发展的标志性节点(Tomasello, 2018)。例如, Baillargeon等人(2010)认为人类心理理论存在两个子系统:子系统一在生命第一年末发展成熟, 使得婴儿可以理解其他个体是否掌握某些信息, 而随后发展的子系统二使得婴儿可以理解与事实不一致的信息性心理状态, 即错误信念。
由于错误信念理解较二级观点采择更为复杂, 因此, 非人灵长类应该也不具备该能力。为验证这一观点, 一些研究者对经典研究范式进行了改编, 例如在食物竞争范式研究中, 如果被试理解竞争者形成了关于食物位置的错误信念, 那么允许它自己选择时, 它会明智做出对自己有利的选择, 但大量研究表明, 在类似实验任务中, 非人灵长类都无法表现出掌握错误信念的能力(以上结论相关实证研究见表4)。
过往研究发现, 儿童在4岁左右时才能对他人错误信念进行心理表征, 不过近年来一些使用内隐任务的实验发现, 1至2岁的婴儿就可以表现出理解他人错误信念的能力(Buttelmann et al., 2015; Knudsen & Liszkowski, 2012; Rakoczy, 2012)。在这些实验中, 研究者不再要求幼儿做出选择, 而是观察幼儿在某种故事情境下自然而然的行为反应是否符合对他人错误信念的理解。由于该实验模式对记忆容量、任务理解与专注度等其他认知能力要求较低, 更容易澄清被试单纯的错误信念理解能力, 一些研究测试了是否非人灵长类也能通过此类内隐任务。研究结果显示, 非人灵长类能够预测其他个体由错误信念而驱动的行为(Krupenye et al., 2016), 它们可以从其他个体的视角出发看问题, 认识到其他个体的行为不是由“现实”而是由“关于现实的信念” (尽管这些信念是错误的)所引发的(以上结论相关实证研究见表4)。
然而一个关键问题是, 这种测试是否真的可以说明非人灵长类能掌握其他个体与现实不一致的观念?在以成人或幼儿为被试进行的研究中, 通过内隐任务测试错误信念并不能获得较为稳定的实验结果, 而且当选择的测量指标不同时——如第一注视方向或注视时间, 甚至会出现相互矛盾的结论(Baillargeon et al., 2018; Barone et al., 2019; Kulke et al., 2019)。Tomasello (2018)认为, 在内隐任务中动物被试可能并不“知晓”行动者的信念是错误的, 它所做的只是追踪行动者的视角以及预测行动者将要做什么。换句话说, 非人灵长类只是掌握了一定的行动规则, 如“他人倾向于去他最后一次见到某物体的地方寻找该物体”, 但它们并没有考虑或理解行动者的心理状态与实际情况间的差异。总之, 目前大多数研究指向的结论依然是, 非人灵长类不具有错误信念理解能力, 或至少它们无法对其他个体的错误信念进行外显心理表征(相关解释见第4部分)。
3 对非人灵长类与人类心理理论共性的解释
研究非人灵长类读心机制的意义不仅在于了解灵长类动物的社会认知能力, 更重要的是, 通过比较分析非人灵长类与人类在心理理论方面的异同点, 有助于推进一些对心理理论基本问题的探索, 如心理理论的进化历程、心理理论的认知结构以及心理理论各种子成分间的关系。在很长一段时间, 众多研究者都认为心理理论是人类祖先在进化中与其他灵长类动物分化后独自发展出的能力, 是人类特有的社会性认知特征, 它的产生是由人类独特的语言、文化和社会化实践所驱动的(Heyes, 1998)。但当前大量研究采用不同反应指标、不同交互对象(人类或同类)或不同任务都证明, 非人灵长类也跨越了心理理论的门槛, 它们可以追随或吸引其他个体注意, 根据潜在目标和意图来理解其他个体行为, 并能从他者视角出发判断其他个体是否可感知到某事物, 这些能力不依附人类的语言和文化, 非人灵长类与人类在心理理论方面有许多共同特征。
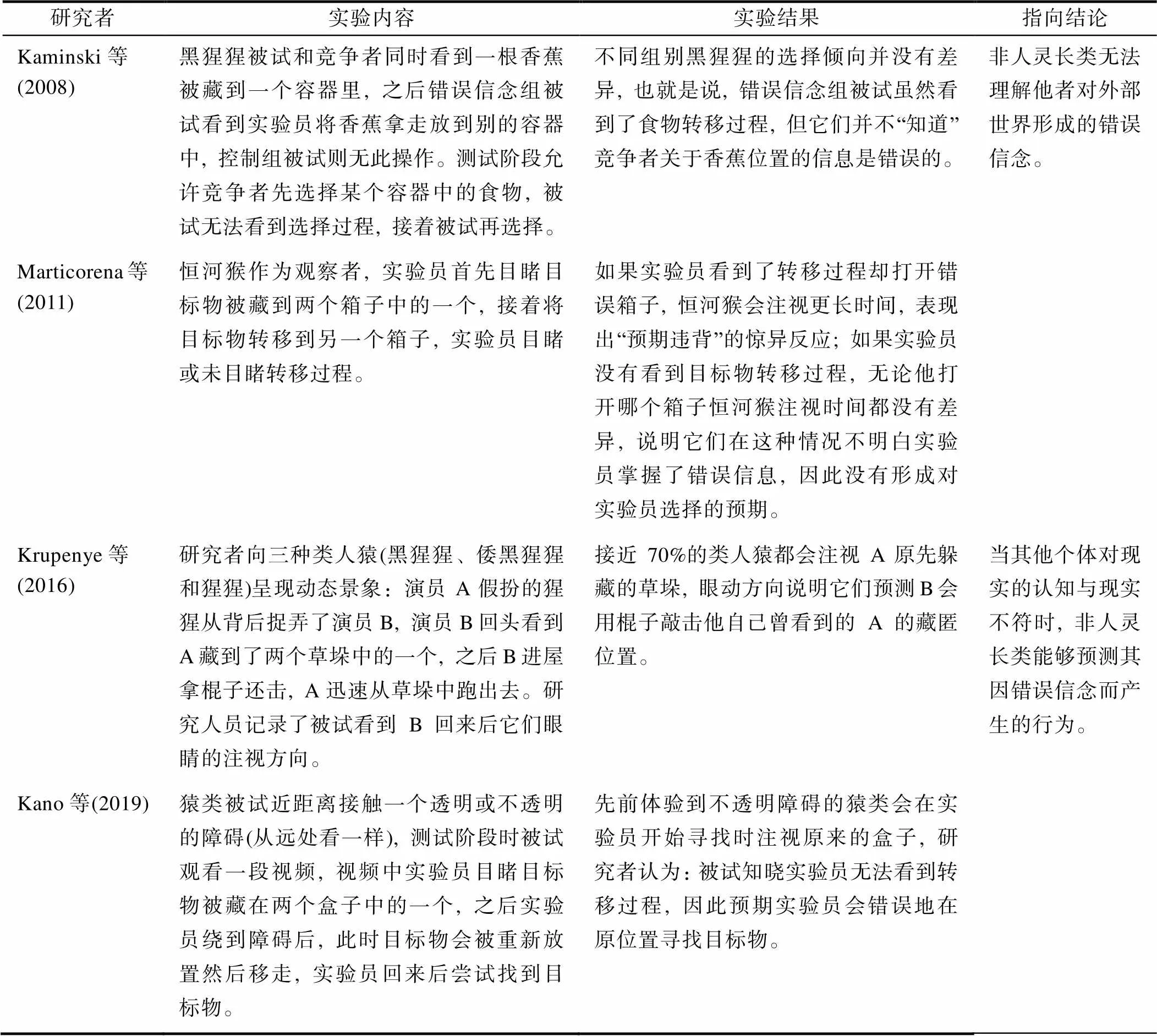
表4 非人灵长类错误信念理解的代表性实证研究
按照进化科学解释, 当不同物种具有某些相似的生理或心理机制时, 可能源于共同遗传(shared descent), 即该机制来自这些不同物种的共同祖先, 也可能源于趋同进化(convergent evolution), 即不同物种在相同进化压力下分别独立进化形成了同一机制(殷融, 赵嘉, 2021)。大量实证研究显示, 非人灵长类与人类在心理理论方面的相似表现是共同遗传的产物, 心理理论在灵长类动物的进化谱系中具有进化连续性(evolutionary continuity)。
3.1 进化连续性假设的行为研究证据
如前文所述, 心理理论具有不同成分, 在人类个体发育过程中, 它们是相继出现的:联合注意是心理理论发展的最早期表现, 幼儿首先形成了关注他人注意方向的能力, 之后才能在此基础上, 逐渐掌握将情境因素、被观察者的动作以及行动目标整合在一起的心理机制, 实现对他人行为意图的理解(Rosati et al., 2016); 同样, 幼儿必须先做到可追随其他个体的注意状态, 之后才发展出一级观点采择机制, 即具备判断其他个体是否能感知到某事物的能力(Baillargeon et al., 2018); 随着发育及经验累积, 幼儿逐渐认识到自己与他人对同一物体的观察会有差异, 于是形成二级观点采择(Moll & Meltzoff, 2011); 另外, 幼儿只有能够代入他人的观察视角, 才具备区分主观现实与客观现实的基础(Tomasello, 2018), 进而理解其他个体对于外部世界的认知可能具有错误信念。
对应到人类个体发育过程中, 如前文所述, 非人灵长类所能掌握的心理理论成分主要是人类幼儿在生命早期就形成的较为基础的心理理论成分, 而无法掌握的复杂心理理论成分在幼儿心理发育周期中出现的也较晚。更重要的是, 包括原猴、新世界猴、旧世界猴和类人猿[1]注:按照进化出现顺序(从早到晚)以及与人类的亲缘关系(从远到近), 可以将灵长类分为原猴类(7000万年前)、新世界猴(4000万年前)、旧世界猴(3000万年前)与类人猿(1600万年前)四个类群。在内的多种非人灵长类都被证明具有基础心理理论成分(Krupenye, 2021)。例如研究发现, 在灵长类动物进化谱系中最早出现的狐猴(大约7000万年前)以及较早出现的卷尾猴(大约4000万年前)和长臂猿(大约4000万年前)也能通过与联合注意、观点采择及意图理解有关的测试(Drayton & Santos, 2014; MacLean et al., 2013; Sánchez-Amaro et al., 2020), 系统发育重建(phylogenetic reconstruction)表明心理理论在灵长类动物中具有深层进化根源(Krupenye, 2021)。
另一方面, 在灵长类进化史上出现较晚、与人类亲属关系更密切的类人猿(如猩猩、黑猩猩和倭黑猩猩)在一些实验中能表现出比其他非人灵长类(如原猴类)更高的读心水平(Horschler et al., 2020; Krupenye & Call, 2019), 这也体现了心理理论在灵长类动物中的进化连续性。例如在食物竞争范式研究中, 汤基猕猴需要更多行为线索才能像黑猩猩一样做出合理选择(Canteloup et al., 2016)。一些类人猿甚至可以通过经验投射来理解其他个体, 例如, 有镜子经验的黑猩猩明白, 镜子可以让个体获得他们身后事物的视觉信息, 即便一个人的面孔和眼睛没有朝向某个方向, 他也能通过镜子进行观察(Lurz et al., 2018); 在Karg等人(2015)的实验中, 黑猩猩看到食物被放到两个不同隔间里, 从黑猩猩视角看两个隔间完全一样, 但从实验员视角看两个隔间一个透明一个不透明。训练阶段的黑猩猩从实验员视角观察隔间, 实验阶段它们要尝试从隔间里偷走食物。结果显示, 当了解实验员视角后, 黑猩猩倾向于先取走不透明隔间里的食物; 但不了解实验员视角时, 黑猩猩从不同隔间取食物的倾向没有差异。也就是说, 黑猩猩能做到依据自己的主观经验来推断模拟实验员的视角。
因此, 已有结论都指向了进化的共同遗传特征:心理理论在灵长类动物谱系中具有进化连续性, 非人灵长类也具备心理理论的基础功能, 不同灵长类动物在心理理论方面的表现只具有程度差异而不是性质差异, 只是对于某些动物尤其是人类来说, 该机制得到了进一步的增强与精细化。
3.2 进化连续性假设的神经科学研究证据
来自神经科学的研究也解释了非人灵长类与人类在心理理论方面的共性, 并进一步证明了进化连续性假设。脑成像研究表明, 不同形式的外显和内隐心理理论任务会激活人类包括内侧前额叶皮层(medial prefrontal cortex, mPFC)、颞上沟(superior temporal sulcus, STS)和颞顶联合区(temporo-parietal junction, TPJ)在内的大脑网络(Marslen-Wilson & Bozic, 2018; Schurz et al., 2014)。研究发现, 一方面, 与非人灵长类相比人类大脑的这些脑区经历了明显扩张(Hill et al., 2010); 另一方面, 人类和非人灵长类大脑的这些部位又具有明显的解剖结构及连通相似性(Mars et al., 2013; Sallet et al., 2013), 更重要的是非人灵长类完成心理理论任务时激活的神经网络也涉及这些脑区。例如, Sliwa和Freiwald (2017)的实验显示, 猕猴观看其他猕猴互动的视频时, 会激活内侧和腹外侧前额叶皮层(medial and ventrolateral prefrontal cortex)。Hayashi等(2020)研究发现, 猕猴完成心理理论实验任务时的眼动倾向与内侧前额叶皮层激活程度具有相关性。研究者向一部分猕猴大脑注射了一种慢病毒载体“hM4Di”及其激活剂“氯氮平N-氧化物” (clozapine N-oxide), 这种病毒可以抑制内侧前额叶皮层的活动, 结果表明, 同时注射了这两种试剂的猕猴在实验时眼动偏好会消失。Ninomiya等人(2020)使用脑成像技术及病毒阻断剂的研究也证实了内侧前额叶皮层在猕猴理解他者行为中的作用。
除内侧前额叶皮层外, Roumazeilles等人(2021)研究发现, 当看到社交场景中的预期违背行为时, 猕猴的中颞上沟(middleSTS)有一处大脑区域会产生敏感反应, 研究者使用预测编码框架(predictive coding framework)分析表明, 该反应模式与人类完成心理理论任务时大脑颞顶联合区的反应模式高度相似, 中颞上沟可能是颞顶联合区在猕猴大脑中的同源区。Ninomiya等人(2021)研究也发现, 在交替执行任务中, 当同伴做出预期违背选择时, 作为观察者的猕猴其中颞上沟脑区的神经元会出现明显活动反应。以上研究结果表明, 萌发出心理理论机制的神经系统集中于内侧前额叶皮层和颞上沟, 这些脑区可以追溯到人类和非人灵长类的共同祖先, 灵长类动物读心能力的神经系统具有深刻进化根源, 而在人类进化过程中, 这些脑区经历了扩张与重组, 导致人类心理理论系统可以达到更复杂水平。不过, 相关的关键性转变发生于何种时期, 以及不同时代人类祖先心理理论能够达到什么水平, 至今还没有确定结论。
4 对非人灵长类与人类心理理论差异性的解释
进化连续性假设解释了灵长类动物读心机制的共同特征, 同时也表明人类心理理论在进化过程中得到了提升强化。从实际测试看, 非人灵长类在心理理论方面与人类的最大差距在于它们无法完成二级观点采择及错误信念理解相关的实验任务。这种区别是由何种内在机制所主导的?它反映了非人灵长类与人类具有哪些心智表征方面的深层差异?不同于人类, 动物被试无法通过语言描述自己的思维, 研究者只能通过行为反应解读其内在心理过程, 这就为解释它们行为模式的逻辑带来了困难。目前, 有几种理论从不同的角度尝试对此进行阐述。
4.1 潜心智化假说
潜心智化假说 (submentalizing hypothesis)认为, 非人灵长类被试在心理理论任务中的表现仅仅是一种对于刺激的低级感知反应, 在这一过程中, 由于知觉线索的刺激, 观察者唤醒了某些回忆, 呈现出知觉线索效应(Heyes, 2014)。例如, 如果非人灵长类被试看到行动者目睹目标物被藏在两个箱子中的一个, 之后行动者寻找目标物, 被试会长时间注视藏有目标物的箱子。按照潜心智化假说的理解, 这不是由于被试“认为”行动者会去他看到藏匿目标物的位置进行寻找, 真正原因是行动者的出现构成一种知觉线索, 刺激被试回忆起目标物被藏匿的场景, 所以被试才会注视藏匿目标物的箱子。潜心智化机制构成了心智理解的基本条件, 但它本身并不涉及对任何心理状态的表征, 非人灵长类只具有潜心智化机制(Heyes, 2014)。
潜心智化假说在理论上可以解释为什么非人灵长类无法完成更复杂的二级观点采择或错误信念任务, 原因在于这些场景下知觉线索效应的缺失。例如, 被试看到目标物在行动者不在场期间发生了转移, 此时行动者不再能充当唤醒被试场景记忆的刺激线索, 因此被试就无法表现出与行动者行动方向一致的反应。该假说主要的问题在于, 是否非人灵长类在心理理论实验任务中的表现确实是基于知觉线索而不是心理表征?如果此结论成立, 那么在同类实验中无生命特征的物品也应该能充当知觉线索, 唤起非人灵长类被试的场景回忆, 导致它们做出特定反应。已有研究证据似乎与此不符, 例如在Krupenye等人(2017)进行的实验中, 研究者设定了两种情境, 一种情境与经典实验一样, 类人猿被试看到行动者目睹目标物被藏在两个箱子中的一个, 之后行动者寻找目标物; 在另一种情境下, 以一个绿色剪影取代行动者, 但该剪影会做出像人类行动者一样的动作, 它“目睹”目标物藏到两个箱子中的一个并去接近某个箱子。结果显示, 被试只会在人类行动时长时间注视目标物所在位置。在另一个类似实验中, 类人猿被试看到人类行动者反复伸手够取目标物, 之后另一个实验员公开转移目标物, 被试会在行动者做出伸手动作时注视目标物所在位置; 但如果把行动者换成一个爪形道具, 则不会出现同样效应(Kano & Call, 2014)。这些研究结果并不符合潜心智化假说的预测, 这说明, 非人灵长类在类似实验中的预期注视反应来自于它们对行动者行为意图及其下一步动作的理解预测, 而不是对记忆的确认, 它们无法完成复杂心理理论任务的原因可能并非因为知觉线索的缺失。
4.2 替代表征假说
替代表征假说(vicarious representation hypothesis)认为, 个体在表征客体时常常引入客体与主体间的关系, 从而为客体赋予关系属性(Nanay, 2020)。关系属性可以以自我为中心(self- centered)也可以以他者为中心(other-centered), 其中以他者为中心的关系属性(other-centered properties)指在不涉及他者情况下无法成立的属性, 例如“某物对他人来说是可食用的”, 而对该关系属性的表征则称为替代表征。“替代”这一概念强调出认知主体不需要对他者心理状态进行表征, 而仅仅是对客体的关系属性进行表征。与心理理论相比, 替代表征是一种要求较低的社会认知形式, 非人灵长类在心理理论任务中表现出的正是替代表征过程(Nanay, 2020)。
例如在食物竞争任务中, 非人灵长类被试倾向于选择不被竞争对手看到的食物, 但它并不需要推测竞争对手的意图或想法, 它只是将食物看作“竞争对手可接近的食物”或“竞争对手不可接近的食物”, 此时被试是把以竞争对手为中心的“可接近”这一关系属性赋予了不同食物, 并在此基础上做出选择。而非人灵长类之所以无法通过二级观点采择或错误信念测试, 是因为在这些场景下它们无法形成稳定的替代表征(Nanay, 2020)。例如在错误信念实验中, 被试目睹目标物在观察者离开时发生了转移, 这实际上改变了被试对目标物的表征方式, 被试会先后产生两种矛盾的替代表征——“行动者看到藏匿位置”的目标物和“行动者未看到藏匿位置”的目标物, 被试无法将两种矛盾的替代表征整合在一起, 因而无法做出正确反应。
总之, 按照替代表征假说的解释, 非人灵长类在心理理论任务中其实是对客体和他者的关系进行了表征, 而不是真正理解他者的心理状态; 它们之所以无法像人类一样完成某些心理理论任务, 是由于无法同时持有两种对立的表征。然而, 替代表征本身似乎并不能排除心理理论的作用, 例如“行动者看到藏匿位置”的目标物, 这一表征本身就包含了对行动者的观点采择。更重要的是研究发现, 一些非人类灵长类确实能够同时持有对一个事物两种不同的心理表征。例如, 类人猿为自己选择食物奖励时, 它们能够摆脱镜头扭曲或遮挡物的影响, 区分一个物体视觉体积与真实体积的差异(Karg et al., 2014; Krachun et al., 2016); Hirel等人(2020)使用透镜和镜子作为工具进行的实验证明, 棕色卷尾猴(Brown Capuchins)和汤基猕猴熟悉装置特性后, 它们可以忽略透镜或镜子带来的欺骗效应, 选择出真正的大份食物。如果非人灵长类能够将“看起来比较小”和“实际上比较大”这两种对立表征相整合, 区分“表象−现实” (appearance-reality)的差异, 它们为什么无法在心理理论任务中将两种对立的替代表征(如“行动者看到藏匿位置”的目标物和“行动者未看到藏匿位置”的目标物)相整合?替代表征理论无法对此进行解释。
4.3 最小限度心理理论假说
最小限度心理理论假说(minimal theory of mind hypothesis)认为, 心理理论可分为最小限度心理理论和成熟心理理论两个系统, 成人二者兼有, 而人类婴儿和非人灵长类则只具有最小限度心理理论(Butterfill & Apperly, 2013)。最小限度心理理论的运作机制与心理理论有所不同:当行动者遇到(encounter)某个场景时, 可以对场景中的关系(如目标物与位置)进行登记(register), 观察者表征的是行动者是否实现了关系登记, 即信念类似(belief-like) 状态, 而不是行动者的真实信念(Apperly & Butterfill, 2009)。在观察者看来, 当行动者要执行针对特定物体的目标导向行为时, 他会按照登记的关系进行行动。
该假说解释了为什么一些非人灵长类可以通过错误信念内隐测试。例如在研究中观察者可以看到目标物从一个箱子转移到另一个箱子, 而行动者看不到。以类人猿(Kano et al., 2019)和猕猴(Hayashi et al., 2020; Horschler et al., 2021)作为观察者的实验证明, 它们会在行动者开始寻找目标物时先看原来放置目标物的箱子, 并对这些箱子注视更长时间。按照最小限度心理理论的解释, 这是由于观察者表征的是“行动者登记了他最后一次遇到目标物的位置”, 因此, 它们在该场景中可以通过注视表达出对行动者行动方向的预期, 但它们本身并没有真正理解行动者的信念, 更没有将行动者信念与真实情况进行比较。
Butterfill和Apperly (2013)强调, 同心理理论相比, 最小限度心理理论的使用会受到较多限制, 一是该机制主要针对目标导向行为, 二是该机制主要在位置场景中能发挥作用, 但在其他场景中则无法实现对“登记”的表征。例如, 如果行动者目睹两份食物被放在不同容器中, 一份较多, 一份较少, 在行动者离开后, 向放置较少食物的容器中再放置更多食物, 此时行动者对不同容器中食物相对数量的信念是错误的, 而非人灵长类作为观察者可能无法预测行动者的选择。
因此, 最小限度心理理论既能解释为什么部分非人灵长类可以在目标物位置移动的内隐测试中表现出预期注视反应, 也能说明为什么它们无法通过其他二级观点采择或错误信念理解任务。然而, 该理论过于笼统, 研究者在阐述中没有对一些基本概念进行特别清晰明了的说明, 如所谓的“登记”是如何运行的?“信念类似状态”与信念表征到底有什么不同?如果这些问题无法回答, 那最小限度心理理论也只是通过创造定义, 用新问题取代了旧问题, 但本质上依然没有澄清非人灵长类在心理理论上的局限之处。
4.4 意识关系假说
意识关系假说(awareness relations hypothesis)认为, 非人灵长类具有一种“意识关系”, 它们能将自己所认知的真实世界信息与其他个体联结起来(Martin & Santos, 2016)。也就是说, 非人灵长类在读心时表征的是真实信息与其他个体之间的“连通−闭合”通道, 基于这种“连通−闭合”特性它们可以完成观点采择任务。但如果观察者认为相关信息在行动者意识之外(行动者没有察觉)发生了变化, 那么该意识关系就会“关闭”, 此时观察者就无法基于意识关系预测行动者的行为(Horschler et al., 2019), 这就解释了为什么非人灵长类无法达成二级观点采择或错误信念理解。意识关系假说与最小限度假说有些相似, 但该假说更加明确具体地指出了观察者表征的内容。
近年来新出现的一种“转移−复原”实验模式可以对意识关系假说进行验证。在这类实验中, 被试先是看到行动者目睹目标物被藏到特定位置, 之后主试在行动者未察觉时移动目标物, 但接着又将目标物移回最初位置。如果被试可以对行动者关于外部世界的信念形成心理表征, 此时行动者依然持有关于目标物位置的正确认知, 被试应该可以预测他们的行为; 但如果像意识关系假说所强调的, 被试表征的是信息与行动者之间的关联性, 在被试眼中, 情境于行动者察觉外发生了变化(尽管又恢复了原状), 此时连通性关闭, 被试便无法预测行动者的行为, 当前的实验结果更为支持后一种可能。
例如在Drayton和Santos (2018)的研究中, 恒河猴看到实验员目睹水果被藏到两个盒子中的一个, 盒子颜色不同, 当把两个盒子的位置进行180度旋转后, 假如实验员没有看到旋转过程, 虽然水果还是在原来盒子里, 实验员具有关于水果位置的“正确信念”, 但当实验员选择“错误”盒子时恒河猴没有表现出预期违背反应。相反, 如果实验员也看到了旋转过程, 恒河猴会在实验员选择“错误”盒子时表现出预期违背反应。可见, 影响恒河猴判断的并不是实验员所持有的信念, 而是他与情境信息之间的关联性(实验员是否观察了情境改变)。
再例如, 在Horschler等人(2019)的研究中, 实验员目睹目标物被放进一个盒子, 之后有人将目标物从盒子里拿出来再放回去, 实验员没有看到该操作, 而恒河猴被试观看了所有过程。如果恒河猴能追踪实验员的心理状态并和真实情况进行比较, 它们应该认为实验员依然掌握目标物的正确位置信息。但实验结果表明, 恒河猴并不期望实验员拥有正确信念。因此, 恒河猴可能能做出的推理是“目标物在A处, X看到了放置过程, 所以X知道目标物的位置”, 而它们无法做出的推理是“目标物在A处, X看到了放置过程, 目标物被移动后又放回A处, 所以X关于目标物位置信息的知识依然是对的”。
总之按照意识关系假说的理解, 非人类灵长类只能做到将客观真实信息与其他个体产生联结, 判断其他个体是否掌握真实信息, 但它们无法像人类那样追踪其他个体的心理状态, 并将他掌握的信息与真实情况进行比较; 一旦外部世界的信息在行动者知觉之外发生改变, 不管这些改变与行动者原有认知是否一致, 非人灵长类都会认为他们无法掌握对外部世界的正确表征。意识关系假说的内容较为清晰具体, 它既能说明为什么非人灵长类能通过观点采择测试, 也能说明为什么它们无法通过二级观点采择与错误信念测试, 具有较强解释力, 同时近年来新出现的“转移−复原”实验模式也进一步证实了该假说的可靠性。但该假说难以澄清的是, 为什么在内隐错误信念理解实验中, 非人灵长类可以对离场后(“意识关系”已关闭)又返回的行动者表现出预期注视反应。
综合来看, 以上4种假说都强调:第一, 在人类个体掌握成熟的心理理论前, 有一种尚不完善的初级表征系统(如潜心智化机制、替代表征机制、最小限度心理理论或意识关系系统), 这些表征系统为成熟心理理论的发展奠定了基础, 非人灵长类只具备初级表征系统; 第二, 非人灵长类之所以难以完成与二级观点采择或错误信念理解相关的任务, 是因为初级表征系统在这些情境中无法再正常运行(如知觉线索效应缺失、替代表征冲突、关系登记失败、意识关系中断); 第三, 非人灵长类无法对其他个体关于外部世界的心理信念形成表征, 信念表征是非人灵长类与人类读心能力的重要分界点。
不同之处在于, 潜心智化假说与替代表征假说更为极端, 它们分别认为非人灵长类在心理理论实验中做出的正确反应主要依靠对场景的记忆确认或对客体的关系属性表征, 直接否认了非人灵长类具有解读他者心理状态的能力。相比之下, 最小限度心理理论假说和意识关系假说则依然支持非人灵长类可以理解他者心理状态, 综合目前实证研究来看, 后两种假说更能兼容已有研究结论, 但它们也都有无法澄清的问题。一种可能是, 二级观点采择与错误信念理解这类高级心理理论能力依赖许多不同的子加工机制, 而不同实验模式对加工机制的需求有所差异, 因此才导致非人灵长类在不同实验模式中产生了看似相互矛盾的表现。另外, 一些研究者质疑当前研究所采用的任务都无法真正测试非人灵长类被试是否构造了他者关于外部世界的知识(Horschler et al., 2020)。因此, 对于非人灵长类与人类心理理论差异及相关解释, 尚需要未来研究更完备的检验。
5 争议问题及研究展望
只有具备了一定的心理理论, 有机体才能理解其他个体的经验、需求和意图, 进而对其行为进行解释和预测, 因此, 一个物种的心理理论发展水平与其社会化认知特征密切相关。本文系统梳理了非人灵长类心理理论的相关实证研究, 重点突出了非人灵长类与人类在心理理论方面的异同点, 并对相关假说解释进行了总结介绍。该领域目前已积累了丰富的研究成果, 但与此同时许多结论仍存在分歧, 一些重要研究方向尚未探索。例如对人类与非人灵长类读心能力差异的几种理论假说, 虽然一定程度上解释了非人灵长类为什么不能像人类一样运用心理理论, 但这些假说都没有涉及到心理理论的进化动因, 即到底哪些因素导致人类获得了更高水平的心理理论, 或哪些因素限制了非人灵长类心理理论的发展。今后的研究可重点聚焦几个主题, 对这些研究主题的关注与探讨可以为解答“人−猿”读心差异的深层原因提供线索, 进一步澄清对动物认知能力的认识; 同时也有助于研究者深入理解人类心理理论的发展及运行机制, 为破解心理理论研究领域广泛存在的一些争论提供启发。
5.1 心理理论与共享意图的关系
本文分析了非人灵长类在心理理论实验任务中表现不同于人类之处, 实际上除了能力差异外, 二者运用心理理论的场景也有所区别, 对于人类来说, 心理理论是一种在竞争与合作场景中都能发挥重要作用的心理机制, 而非人灵长类似乎更倾向于在竞争场景下启动读心机制, 竞争活动会更显著影响它们心理理论的运用。例如, 当有障碍物遮挡视线时, 尽管恒河猴了解自己视角与行动者视角是不一样的, 它们也能够做到移动至行动者视角去观察行动者所关注的内容, 但只有存在竞争时它们才会这么做, 在竞争环境下, 恒河猴更“善于”利用自己的观点采择能力, 而在非竞争环境下它们缺乏足够动机这样做(Bettle & Rosati, 2019)。
再例如, 在Hare和Tomasello (2004)完成的一个经典实验中, 研究者教导黑猩猩玩一个选择游戏, 黑猩猩知道食物被藏在三个桶中的一个, 它们只有一次选择机会; 在黑猩猩做出选择前, 一个实验员用手指向其中一个桶(人类幼儿在很小时就能理解此时这一指向性手势的含义), 黑猩猩会盯着实验员手指的方向看, 但在选择时却会随便选一桶。也就是说, 它们注意方向会被手势所吸引, 可无法理解实验员是将有价值的信息分享给它们。然而在另一种竞争条件下, 实验员热身阶段就和猩猩抢食物, 之后在实验选择阶段实验员努力将手伸向其中一个桶, 这时黑猩猩倾向于选择这个桶, 它们可能明白, 竞争者想获得的桶里会有食物。由此可见, 黑猩猩对竞争意图更为敏感。实际上, 许多早期研究之所以得出非人灵长类缺乏心理理论的结论, 或许正是由于实验模式的原因, 在早期实验中, 非人灵长类无法在合作情境下理解人类演示者给出的线索。
这些研究表明, 非人灵长类可能根本无法理解互惠合作意图或善意沟通, 这限制了它们运用心理理论的情境。与之相对应, 研究发现, 尽管非人灵长类对他人的注意力状态很敏感, 它们会追随其他个体目光, 同时可以识别他者的具体目标, 但非人灵长类的目光追随行为通常是由“自主关注”所驱动的, 如它们通过眼神方向来判断对方有没有在关注自己想获得的特定目标物, 但它们很难理解其他个体在目光中蕴含的“共享意图(shared intent)”。当儿童捕捉到个体向其发出的交际信号后, 会追随信息发出者的注视方向, 他们期待信息发出者的视觉线索是有所指向的。如果信息发出者注视方向没有儿童期待的目标物出现, 他们会注视更长时间(Csibra & Volein, 2008; Senju & Csibra, 2008), 但类似现象在非人灵长类身上无法得以复现。例如, 在Kano等人(2018)以黑猩猩为被试进行的实验中, 实验组的实验员主动向黑猩猩释放交际信号, 而控制组则通过其他刺激将黑猩猩的注意力吸引到实验员面部, 之后实验员注视一个目标物或干扰物。释放交际信号会导致被试对目标物和干扰物的注视时间都更长, 但与不释放交际信号的情况相比, 释放交际信号并不会使被试对目标物的注视时间显著长于对干扰物的注视时间。也就是说, 黑猩猩没有理解人类发出的交际信号和之后注视方向的组合功能, 它们无法识别出交际信号中释放的分享信息的意图, Bettle和Rosati (2021a)以恒河猴为被试的类似实验也获得了相同结果。
因此一种合理的猜想是, 虽然非人灵长类能够预测他者心理状态, 但这种能力的基础是与其他个体开展竞争, 它们更容易感知到来自他者的竞争意图而不是合作意图。而在人类进化过程中, 特殊社会压力导致人类发展出对共享意图的敏感性, 例如, 一旦人类祖先对技术和工具产生高度依赖, 自然选择便可能“选择”出互相分享知识经验的倾向, 知识传播需求为精准读心机制的进化创造了一个关键先决条件。当个体间能够建立共同目标并为了共同目标而相互协调时, 心理理论能力会得到进一步提高。今后研究应关注这一可能性, 通过实证研究方式, 进一步探索共享意图对灵长类心理理论发展的影响。
5.2 心理理论与语言机制的关系
复杂语言交流系统与心理理论系统都是人类重要的社会化认知特征, 二者之间影响关系是心理理论研究领域一个一直存在争议的问题。研究发现, 人类婴儿语言的萌芽期与快速发展期正对应了心理理论的萌芽期与快速发展期(Milligan et al., 2007); 儿童接受语言训练可以提高他们在心理理论任务中的得分(Hale & Tager-Flusberg, 2003); 有手语学习经验的聋哑儿童比无手语经验的聋哑儿童在心理理论发展水平上要更高(Pyers & Senghas, 2009); 个体心理理论发展水平与其对讽刺语及双重否定等对话技巧的理解力存在相关性(Matthews et al., 2018); 患有自闭症的儿童会同时表现出语言发展迟缓和缺乏联合注意的特征, 而联合注意可以预测幼儿语言发展趋势(Bettle & Rosati, 2021b)。
在人类进化过程中, 心理理论是否构成了语言进化的助推器?首先, 语言沟通的前提是个体间能互相理解对方想法, 更高阶的观点采择能力可以使个体代入交流对象的视角, 让其超越自我为中心来认知情境, 也就是说, 心理理论发展水平决定了接收者对信息的理解。事实上, 人类所有的交流形式, 无论是手语、口语还是书面语, 都依赖交流者可以依据时空背景与语境对交流符号的意义进行语用推理(pragmmatic inferencing), 语义欠明是人类语言交流的重要特征, 如果接受者无法代入信息发出者的视角, 将会很难完全理解语言符号的真实意义(Berman et al., 2010)。例如, 当一个人对另一个人说“别坚持了”时, 这句话可以理解为要求对方放弃某观点、要求对方停止某个具体行为或善意地要求对方休息等多重意义, 而具体含义必须在情境中通过推理获得。从这点看, 心理理论是人类复杂语言交流得以实现的重要基础, 凭借心理理论, 个体能够在具体情境中理解他人的沟通意图(Scott-Phillips, 2015)。更重要的是, 如果个体间能够预测彼此某些行为模式的结果, 他们可以在长期互动中形成一些仪式化(ritualization)表达。例如, A做出一系列动作XYZ后, 会引发B的回应, 长此以往, B在A做出第一个动作X时就会有所回应, 同时A也意识到, 只要做出X就会引发B的回应, X就成为了一种仪式化表达, 而这正是语言形成的第一步。也就是说, 凭借心理理论, 同伴能在共同经验的基础上将具体事物与语言符号之间建立联系, 使抽象符号获得表征意义(Pleyer, 2017)。因此, 人类读心能力的提高可能是促进语言进化的关键转变, 也正是因为心理理论方面的限制, 大大制约了非人灵长类语言表征系统的发展。
然而另一方面, 语言符号又是抽象表征的重要中介工具, 非人灵长类只能以图像思维表征其他个体的行为或心理状态(Bettle & Rosati, 2021b), 但人类可以将各种关系、信息及他者信念以语言形式保存下来, 实现对这些内容的加工, 这是人类与非人灵长类在心理表征方面的重要区别。当以语言作为表征形式时, 主体可以编码复杂的嵌套内容, 比如“诸葛亮认为司马懿会相信他在空城中布置了埋伏”。如果不借助语言系统进行梳理, 很难想象人类如何完成这种复杂的递归式心智解读。此外, 在个体发育过程中, 要认识和理解他人心理状态并对其行为作出准确预测, 儿童必须积累一定的个人知识和经验, 语言作为表征外部世界及内部意识的符号系统, 为儿童获取知识经验提供了最有效途径。
从这一视角看, 语言系统又是复杂心理理论的前提, 可能正是由于语言表征功能的突破, 导致人类形成了比非人灵长类更高水平的心理理论。当然, 心理理论和语言之间也可能存在其他关系, 例如心理理论的基础成分促成了复杂语言系统的进化, 而语言系统又进一步提升了人类心理理论水平; 或者二者的进化演变都受到了第三种因素的影响, 例如随着生存环境的改变, 人类祖先出现了更高的合作与交流需求, 这一进化趋势既是人类独有认知能力自然延伸的结果, 同时也重塑了人类的认知系统, 在相互作用的循环中, 随着行为多样化的扩展, 心理理论与交流模式都日趋复杂。目前这些假设均未得以检验, 在今后研究中, 研究者应关注非人灵长类心理理论特征与其交流系统间的关系, 通过对不同非人灵长类开展比较研究, 为揭示人类的心理理论及语言进化之谜寻求启示。
5.3 心理理论与社会生态环境的关系
除了有别于人类外, 不同非人灵长类动物之间的认知特征与行为模式本身也存在很大差异, 而这些差异又与它们的社会生态环境具有共变关系, 这就为理解心理理论的进化动因提供了重要线索。例如, Sallet等人(2011)研究发现, 猕猴大脑中颞上沟脑区的大小与社群规模具有正相关性。MacLean等人(2013)比较了5种群体规模不同的狐猴在食物竞争任务中观点采择能力的表现, 动物被试有机会从能看到或不能看到它们的实验员那里偷取食物, 结果显示, 群体规模较大的物种在竞争性偷窃任务中表现得比群体规模较小的物种更好。Nolte和Call (2021)让黑猩猩或倭黑猩猩配对完成任务(相比黑猩猩, 倭黑猩猩的群体规模更大), 被试只有将合适的工具传递给搭档, 对方才能成功操纵一个装置。在无收益组, 被试的帮助行为不会得到任何回报, 而在合作组, 双方都可以通过帮助行为获得回报。结果显示, 两种条件下黑猩猩几乎都没有与同类分享工具, 而所有雌性倭黑猩猩都互相分享了工具。
除了种群规模外, 一些学者认为, 更专制且具有攻击性的社会环境可以作为复杂社会认知技能的催化剂(Gavrilets & Vose, 2006), 这种观点被称为“马基雅维利智力假说(Machiavellian intelligence hypothesis)”, 与之相对立的自我驯化假说(self domestication hypothesis)则认为, 复杂社会认知技能源于对包容性社交环境的需要(Hare & Woods, 2020), 心理理论的进化也符合这一规律。为解答该争议, 一些研究探讨了在社会容忍方面有差异的非人灵长类是否会有不同的心理理论表现, 研究结果更支持自我驯化假说。例如, Kano等人(2015)研究发现, 与黑猩猩相比, 倭黑猩猩相互之间的关系更为友善, 这导致它们从小就在解读同伴的社交线索(如凝视方向和面部表情)方面可以获得更多经验。Joly等人(2017)的实验以不同种类猕猴为被试, 在实验任务中, 被试必须解读主试给出的线索才能从隐藏位置获得食物奖励。实验结果显示, 更友好宽容的那类猕猴更有可能成功完成任务。Rosati和Santos (2017)在研究中比较了地中海猕猴和恒河猴的注视追随特征。研究发现, 两种动物在幼年时都表现出很高的注视跟随性, 但恒河猴随着年龄增长联合注意力水平会下降, 而社会容忍度更高、攻击性更低的地中海猕猴则没有。
总之, 许多研究者都认为, 社会认知是随着群体生活的需求而得以进化的。在灵长类动物群体中, 个体必须与自己的群体伙伴开展竞争或合作以获得生存及繁殖机会, 因此, 自然选择会更青睐那些有利于个体开展社交、实施社会控制以及解释与预测他者行为的认知机制。心理理论作为一种重要的社会认知特征, 必然与一个物种的社会生态环境密切相关, 今后通过开展更多的差异性比较研究, 可以让研究者探索心理理论与社会属性的关系, 同时也可以让研究者检验心理理论各子成分是否具有不同的社会驱动因素, 为理解心理理论的获得机制提供重要依据。
陈丽娟, 徐晓东. (2020). 文学阅读如何影响读者的心理理论.(3), 434−442.
霍超, 李祚山, 孟景. (2021). 自闭症谱系障碍个体的共情干预: 扬长还是补短?(5), 849−863.
莫书亮, 赵迎春, 苏彦捷. (2004). 心理理论的比较认知研究.(6), 860−867.
皮忠玲, 莫书亮. (2013). 婴儿心理理论的发展: 表现和机制.(8), 1408−1421.
钱淼, 傅根跃. (2014). 心理理论的自发反应范式: 方法、结果与解释.(1), 27−37.
隋雪, 史汉文, 李雨桐. (2021). 语言加工过程中的观点采择及其认知机制.(6), 990−999.
殷融, 赵嘉. (2021). 语法的进化连续性及进化起源解释.(7), 1264−1278.
Apperly, I. A., & Butterfill, S. A. (2009). Do humans have two systems to track beliefs and belief-like states?(4), 953–970.
Atsumi, T., Koda, H., & Masataka, N. (2017). Goal attribution to inanimate moving objects by Japanese macaques (Macaca fuscata).(1), 40033−40033.
Atsumi, T., & Nagasaka, Y. (2015). Perception of chasing in squirrel monkeys (Saimiri sciureus).(6), 1243−1253.
Aychet, J., Pezzino, P., Rossard, A., Bec, P., Blois-Heulin, C., & Lemasson, A. (2020). Red-capped mangabeys (Cercocebus torquatus) adapt their interspecific gestural communication to the recipient's behaviour.(1), 12843.
Baillargeon, R., Buttelmann, D., & Southgate, V. (2018). Invited commentary: Interpreting failed replications of early false-belief findings: Methodological and theoretical considerations., 112−124.
Baillargeon, R., Scott, R. M., & He, Z. (2010). False-belief understanding in infants.(3), 110−118.
Barone, P., Corradi, G., & Gomila, A. (2019). Infants' performance in spontaneous-response false belief tasks: A review and meta-analysis., 101350.
Berman, J. M. J., Chambers, C. G., & Graham, S. A. (2010). Preschoolers’ appreciation of speaker vocal affect as a cue to referential intent.(2), 87−99.
Bettle, R., & Rosati, A. G. (2019). Flexible gaze-following in rhesus monkeys.(5), 673−686.
Bettle, R., & Rosati, A. G. (2021a). The evolutionary origins of natural pedagogy: Rhesus monkeys show sustained attention following nonsocial cues versus social communicative signals.(1), e12987.
Bettle, R., & Rosati, A. G. (2021b). The primate origins of human social cognition.(2), 96−127
Bohn, M., Call, J., & Tomasello, M. (2016). The role of past interactions in great apes’ communication about absent entities.(4), 351−357.
Bräuer, J., Call, J., & Tomasello, M. (2007). Chimpanzees really know what others can see in a competitive situation.(4), 439−448.
Buttelmann, D., Carpenter, M., Call, J., & Tomasello, M. (2007). Enculturated chimpanzees imitate rationally.(4), F31−F38.
Buttelmann, D., Schütte, S., Carpenter, M., Call, J., & Tomasello, M. (2012). Great apes infer others’ goals based on context.(6), 1037−1053.
Buttelmann, F., Suhrke, J., & Buttelmann, D. (2015). What you get is what you believe: Eighteen-month-olds demonstrate belief understanding in an unexpected- identity task., 94−103.
Butterfill, S. A., & Apperly, I. A. (2013). How to construct a minimal theory of mind.(5), 606−637.
Canteloup, C., & Meunier, H. (2017). 'Unwilling' versus 'unable': Tonkean macaques' understanding of human goal-directed actions., e3227.
Canteloup, C., Piraux, E., Poulin, N., & Meunier, H. (2016). Do Tonkean macaques (Macaca tonkeana) perceive what conspecifics do and do not see., e1693.
Castellano-Navarro, A., Macanás-Martínez, E., Xu, Z., Guillén-Salazar, F., MacIntosh, A. J. J., Amici, F., & Albiach-Serrano, A. (2021). Japanese macaques’ (Macaca fuscata) sensitivity to human gaze and visual perspective in contexts of threat, cooperation, and competition.(1), 5264.
Csibra, G., & Volein, Á. (2008). Infants can infer the presence of hidden objects from referential gaze information.(1), 1−11.
Deshpande, A., Gupta, S., & Sinha, A. (2018). Intentional communication between wild bonnet macaques and humans.(1), 5147.
Drayton, L. A., & Santos, L. R. (2014). Capuchins’ (Cebus apella ) sensitivity to others’ goal-directed actions in a helping context.(3), 689−700.
Drayton, L. A., & Santos, L. R. (2017). Do rhesus macaques, Macaca mulatta, understand what others know when gaze following?, 193−199.
Drayton, L. A., & Santos, L. R. (2018). What do monkeys know about others' knowledge?, 201−208.
Gavrilets, S., & Vose, A. (2006). The dynamics of Machiavellian intelligence.(45), 16823−16828.
Hale, C. M., & Tager-Flusberg, H. (2003). The influence of language on theory of mind: A training study.(3), 346−359.
Hare, B., & Tomasello, M. (2004). Chimpanzees are more skilful in competitive than in cooperative cognitive tasks.(3), 571−581.
Hare, B., & Woods, V. (2020).. New York: Penguin Random House.
Hayashi, T., Akikawa, R., Kawasaki, K., Egawa, J., Minamimoto, T., Kobayashi, K., ... Hasegawa, I. (2020). Macaques exhibit implicit gaze bias anticipating others’ false-belief-driven actions via medial prefrontal cortex.(13), 4433−4444.e5
Heyes, C. (1998). Theory of mind in nonhuman primates.(1), 101−114.
Heyes, C. (2014). Submentalizing: I am not really reading your mind.(2), 131−143.
Hill, J., Inder, T., Neil, J., Dierker, D., Harwell, J., & Essen, D. V. (2010). Similar patterns of cortical expansion during human development and evolution.(29), 13135−13140.
Hirel, M., Thiriau, C., Roho, I., & Meunier, H. (2020). Are monkeys able to discriminate appearance from reality.104123.
Horschler, D. J., MacLean, E. L., & Santos, L. R. (2020). Do non-human primates really represent others’ beliefs?(8), 594−605.
Horschler, D. J., Santos, L. R., & MacLean, E. L. (2019). Do non-human primates really represent others' ignorance? A test of the awareness relations hypothesis., 72−80.
Horschler, D. J., Santos, L. R., & MacLean, E. L. (2021). How do non-human primates represent others' awareness of where objects are hidden?, 104658.
Joly, M., Micheletta, J., Marco, A. D., Langermans, J. A., Sterck, E. H. M., & Waller, B. M. (2017). Comparing physical and social cognitive skills in macaque species with different degrees of social tolerance.(1862), 20162738.
Kaminski, J., Call, J., & Tomasello, M. (2008). Chimpanzees know what others know, but not what they believe.(2), 224−234.
Kano, F., & Call, J. (2014). Great apes generate goal-based action predictions: An eye-tracking study.(9), 1691−1698.
Kano, F., Hirata, S., & Call, J. (2015). Social attention in the two species of pan: Bonobos make more eye contact than Chimpanzees.(6), e0129684.
Kano, F., Krupenye, C., Hirata, S., Tomonaga, M., & Call, J. (2019). Great apes use self-experience to anticipate an agent’s action in a false-belief test.(42), 20904−20909.
Kano, F., Moore, R., Krupenye, C., Hirata, S., Tomonaga, M., & Call, J. (2018). Human ostensive signals do not enhance gaze following in chimpanzees, but do enhance object- oriented attention.(5), 715−728.
Karg, K., Schmelz, M., Call, J., & Tomasello, M. (2014). All great ape species (Gorilla gorilla, Pan paniscus, Pan troglodytes, Pongo abelii) and two-and-a-half-year-old children (Homo sapiens) discriminate appearance from reality.(4), 431−439.
Karg, K., Schmelz, M., Call, J., & Tomasello, M. (2015). The goggles experiment: Can chimpanzees use self- experience to infer what a competitor can see?, 211−221.
Karg, K., Schmelz, M., Call, J., & Tomasello, M. (2016). Differing views: Can chimpanzees do Level 2 perspective-taking?(3), 555−564.
Knudsen, B., & Liszkowski, U. (2012). 18-month-olds predict specific action mistakes through attribution of false belief, not ignorance, and intervene accordingly.(6), 672−691.
Krachun, C., Lurz, R., Russell, J. L., & Hopkins, W. D. (2016). Smoke and mirrors: Testing the scope of chimpanzees' appearance-reality understanding., 53−67.
Krupenye, C. (2021). The evolution of mentalizing in humans and other primates. In M. Gilead & K. N. Ochsner (Eds.),(pp. 107−129). Cham: Springer International Publishing.
Krupenye, C., & Call, J. (2019). Theory of mind in animals: Current and future directions.(6), e1503.
Krupenye, C., Kano, F., Hirata, S., Call, J., & Tomasello, M. (2016). Great apes anticipate that other individuals will act according to false beliefs.(6308), 110− 114.
Krupenye, C., Kano, F., Hirata, S., Call, J., & Tomasello, M. (2017). A test of the submentalizing hypothesis: Apes' performance in a false belief task inanimate control.(4), 1343771.
Kulke, L., Wübker, M., & Rakoczy, H. (2019). Is implicit theory of mind real but hard to detect? Testing adults with different stimulus materials.(7), 190068.
Luo, Y. (2011). Do 10-month-old infants understand others' false beliefs?(3), 289−298.
Lurz, R., Krachun, C., Mahovetz, L., Wilson, M. J. G., & Hopkins, W. D. (2018). Chimpanzees gesture to humans in mirrors: Using reflection to dissociate seeing from line of gaze., 239−249.
MacLean, E. L., & Hare, B. (2012). Bonobos and chimpanzees infer the target of another's attention.(2), 345−353.
MacLean, E. L., Sandel, A. A., Bray, J., Oldenkamp, R. E., Reddy, R. B., & Hare, B. A. (2013). Group size predicts social but not nonsocial cognition in Lemurs.(6), e66359.
Mars, R. B., Sallet, J., Neubert, F. X., & Rushworth, M. F. S. (2013). Connectivity profiles reveal the relationship between brain areas for social cognition in human and monkey temporoparietal cortex.(26), 10806−10811.
Marslen-Wilson, W. D., & Bozic, M. (2018). Dual neurobiological systems underlying language evolution: Inferring the ancestral state., 176−181.
Marticorena, D. C. W., Ruiz, A. M., Mukerji, C., Goddu, A., & Santos, L. R. (2011). Monkeys represent others’ knowledge but not their beliefs.(6), 1406−1416.
Martin, A., & Santos, L. R. (2016). What cognitive representations support primate theory of mind.(5), 375−382.
Matthews, D., Biney, H., & Abbot-Smith, K. (2018). Individual differences in children’s pragmatic ability: A review of associations with formal language, social cognition, and executive functions.(3), 186−223.
Milligan, K., Astington, J. W., & Dack, L. A. (2007). Language and theory of mind: Meta-analysis of the relation between language ability and false-belief understanding.(2), 622−646.
Moll, H., & Meltzoff, A. N. (2011). How does it look? Level 2 perspective-taking at 36 months of age.(2), 661−673.
Mundy, P. C., & Newell, L. (2007). Attention, joint attention, and social cognition.(5), 269−274.
Nanay, B. (2020). Vicarious representation: A new theory of social cognition., 104451.
Ninomiya, T., Noritake, A., & Isoda, M. (2021). Live agent preference and social action monitoring in the macaque mid-superior temporal sulcus region.(44), e2109653118.
Ninomiya, T., Noritake, A., Kobayashi, K., & Isoda, M. (2020). A causal role for frontal cortico-cortical coordination in social action monitoring.(1), 5233−5233.
Nolte, S., & Call, J. (2021). Targeted helping and cooperation in zoo-living chimpanzees and bonobos.(3), 201688.
O’Grady, C., Kliesch, C., Smith, K., & Scott-Phillips, T. C. (2015). The ease and extent of recursive mindreading, across implicit and explicit tasks.(4), 313−322.
Pleyer, M. (2017). Protolanguage and mechanisms of meaning construal in interaction., 69−90.
Premack, D., & Woodruff, G. (1978). Does the chimpanzee have a theory of mind.(4), 515−526.
Pyers, J. E., & Senghas, A. (2009). Language promotes false-belief understanding: Evidence from learners of a new sign language.(7), 805−812.
Rakoczy, H. (2012). Do infants have a theory of mind.(1), 59−74.
Rochat, M. J., Serra, E., Fadiga, L., & Gallese, V. (2008). The evolution of social cognition: Goal familiarity shapes monkeys' action understanding.(3), 227−232.
Rosati, A. G., Arre, A. M., Platt, M. L., & Santos, L. R. (2016). Rhesus monkeys show human-like changes in gaze following across the lifespan.(1830), 20160376.
Rosati, A. G., & Santos, L. R. (2017). Tolerant Barbary macaques maintain juvenile levels of social attention in old age, but despotic rhesus macaques do not., 199−207.
Roumazeilles, L., Schurz, M., Lojkiewiez, M., Verhagen, L., Schüffelgen, U., Marche, K., ... Sallet, J. (2021). Social prediction modulates activity of macaque superior temporal cortex.(38), eabh2392.
Sallet, J., Mars, R. B., Noonan, M. P., Andersson, J. L., O’Reilly, J. X., Jbabdi, S., … Rushworth, M. F. S. (2011). Social network size affects neural circuits in Macaques.(6056), 697−700.
Sallet, J., Mars, R. B., Noonan, M. P., Neubert, F. X., Jbabdi, S., O'Reilly, J. X., ... Rushworth, M. F. (2013). The organization of dorsal frontal cortex in humans and macaques.(30), 12255− 12274.
Sánchez-Amaro, A., Tan, J., Kaufhold, S. P., & Rossano, F. (2020). Gibbons exploit information about what a competitor can see.(2), 289−299.
Schurz, M., Radua, J., Aichhorn, M., Richlan, F., & Perner, J. (2014). Fractionating theory of mind: A meta-analysis of functional brain imaging studies., 9−34.
Scott-Phillips, T. (2015).. New York: Macmillan International Higher Education.
Senju, A., & Csibra, G. (2008). Gaze following in human infants depends on communicative signals.(9), 668−671.
Sliwa, J., & Freiwald, W. A. (2017). A dedicated network for social interaction processing in the primate brain.(6339), 745−749.
Tomasello, M. (2018). How children come to understand false beliefs: A shared intentionality account.(34), 8491−8498.
Yamamoto, S., Humle, T., & Tanaka, M. (2012). Chimpanzees’ flexible targeted helping based on an understanding of conspecifics’ goals.(9), 3588−3592.
Comparative studies of mind reading: Similarities and differences in theory of mind between non-human primates and humans and corresponding explanations
YIN Rong
(School of Teacher Education, Weifang University, Weifang 261061, China)
Comparative studies using non-human primates is an important way to explore the essence, origin and evolution of theory of mind. A large number of empirical studies have shown that non-human primates have some basic components of a theory of mind, including joint attention, goal understanding and perspective-taking, suggesting that theory of mind has evolutionary continuity in primate evolutionary lineages. However, non-human primates are unable to achieve second-order perspective selection and false belief understanding as humans do. The submentalizing hypothesis, vicarious representation hypothesis, minimal theory of mind hypothesis and awareness relations hypothesis attempt to explain the differences. Although specific views and perspectives vary, these theories all emphasize that non-human primates are unable to form representations of other individuals' mental beliefs about the external world, and that belief representation is an important dividing point between non-human primates and humans in mind reading. Future research needs to further clarify the interaction between shared intention, language mechanism, social ecological environment and theory of mind.
non-human primates, theory of mind, joint attention, perspective-taking, false belief
2022–02–16
* 山东省教育科学“十四五”规划课题2021QYB010资助。
殷融, E-mail: yorkns@sina.cn
B844
